
Abstract
Introduction
C
One promising modality is low-level laser irradiation (LLLI), also known as low-level laser therapy (LLLT). LLLI is one of developments in phototherapy and is a relatively noninvasive treatment in which nonthermal laser irradiation is applied to the target site. The light penetrates the skin without any heating effect. This treatment does not also damage the skin and does not have any side effects. 6 LLLI activates existing biological chromophores in cells and organisms at low doses of visible light of an appropriate wavelength, leading to the generation of reactive oxygen species such as singlet oxygen and superoxides. 7 In addition to the photodynamic effects of LLLI on cells, 8 there have been numerous reports describing the phenomena of biostimulation and bioinhibition to resolve inflammation and trigger the immune system. 9 Commonly used wavelengths of LLLI (685 and 830 nm) applied at fluences in the range of 1–20 J/cm2 appear to be associated with considerable changes in bacterial growth, even in the absence of photosensitizers. 10,11
During the past two decades, a large amount of reliable information in this modern field of medicine has been acquired, and most of the data have been obtained from in vitro laboratory investigations. LLLI has been shown to have potential sensitizing effects on Trichophyton rubrum, 12 C. albicans. 13 and Aspergillus niger, 14 facilitating the killing of these microorganisms. However, in vivo research is extremely important. The action of LLLI on microorganisms may be affected by the environmental conditions of the target sites, including the presence of saliva, the pH, mucosa characteristics, and the action of the immune system. 15 To answer this question, many C. albicans infection models have been developed in experimental animals. Historically, the rat has been used far more often than the mouse as a host for experimental oral Candida infections, as the animal's mouth provides an accessible area in which an infection resembling denture stomatitis can be established. Alternatively, systemic Candida infection in mice has been used to explore the potential application of alternative treatment protocols in both immunocompromised and immunocompetent settings. 16
Despite the promising results obtained in vitro, no rigorous study has focused on the alteration in pathogenicity of C. albicans in an animal model in response to LLLI. Our animal study was performed to confirm that the morphological changes in C. albicans caused by in vitro LLLI led to changes in pathogenicity, as well. Therefore, in the present study, we investigated the effects of LLLI using a diode laser at 685 and 830 nm wavelengths on the pathogenicity of C. albicans in the absence of a photosensitizer. These effects were assessed using in vitro growth curve method and an in vivo model of disseminated candidiasis in mice.
Materials and Methods
Fungal isolates
Four clinical C. albicans isolates from the vaginal discharge of patients with recurrent vulvovaginal candidiasis were used in this study. For the preparation of each isolate, a smear was prepared from a clinical specimen using a sterile cotton swab and glass slides. Microscopic examination of the sample was performed to detect the presence of yeast by methylene blue staining. Further microscopic examination was performed using saline and 10% KOH, and the isolates were grown on Sabouraud dextrose agar containing chloramphenicol (SC Difco, East Molesey, U.K.). Species level identification was performed using Candida Chrome agar (CCA, Difco), and further characterization was performed by assessing germ tube production after 3 h (35–37°C) of incubation in serum and the morphology of the yeast when grown on corn meal agar (CMA, Difco) containing Tween 80. Sugar assimilation tests were performed using the API 20C AUX system (BioMerieux SA, Marcy-L'Etoile, France). 17 A stock isolate was serially passaged on SC and incubated at 35–37°C for 48–72 h. After a pure culture was obtained, the selected fungal isolate was used for the entire experiment to provide standardization. C. albicans colonies were then transferred to treatment and control plates so that there was only one colony <1 cm2 surface area on each plate.
Antifungal susceptibility testing
In vitro antifungal susceptibility testing was performed using the broth microdilution method according to the recommendations of the European Committee for Antimicrobial Susceptibility Testing (EUCAST) 18 and the Clinical and Laboratory Standards Institute (CLSI) guidelines. 19 The following antifungal agents were used: amphotericin B (AMB), flucytosine (5-FC), fluconazole (FCZ), itraconazole (ITC), voriconazole (VRC), caspofungin (CAS). and terbinafine (TER).
LLLI
A total of 12 plates were used for each treatment and control group. C. albicans colonies were directly exposed to various doses of laser light that were chosen according to commonly applied fluences for laser therapies. The BTL-5000 Laser series (Medicinos Projektai, Prague, Czech Republic) was used for the laser irradiation. This device was specifically designed to provide uniform irradiation from beyond the wall of the culture plates on which the microorganisms were seeded, according to the parameters established for the study. To avoid contamination, the laser beam was delivered from 1 cm above the surface of the only colony on the plate through the glassy door of the plate (Fig. 1).
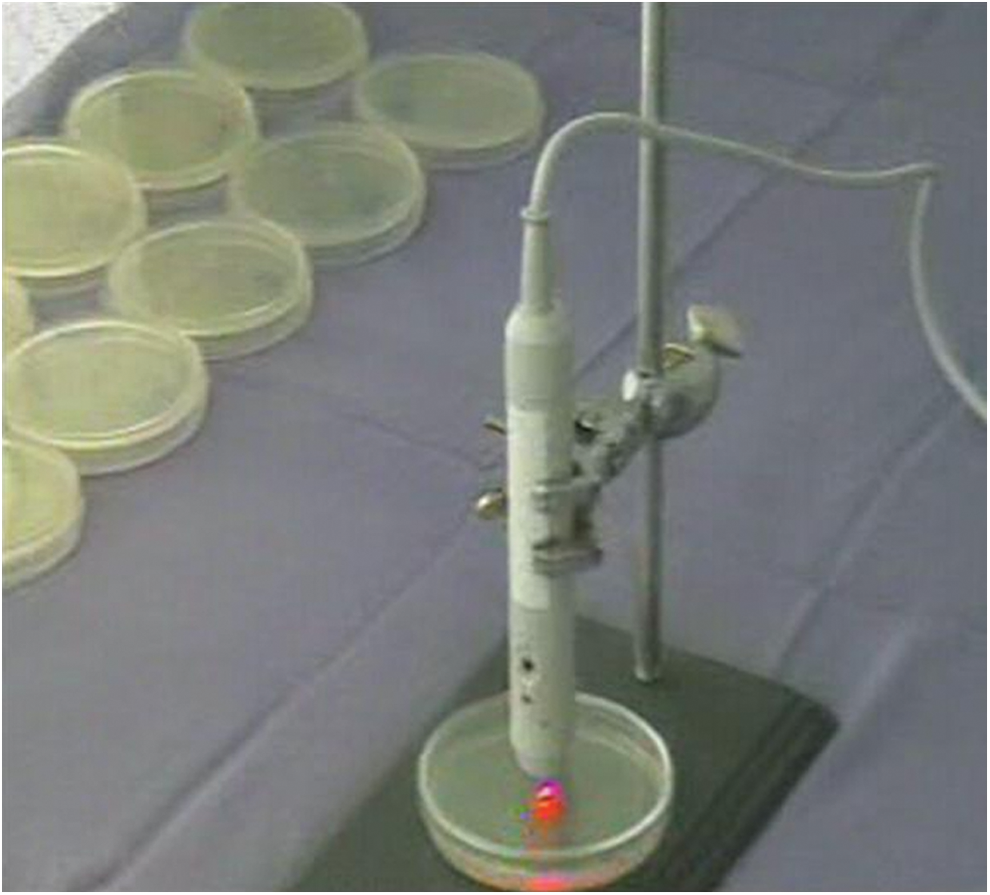
The image of laser application.
The reduction in power density caused by this type of irradiation was calculated to be 50% using a power meter (OPHIR, USA). The treatment groups were irradiated with nine energies: 3, 5, 10, and 20 J at 685 nm (continuous wave [CW], 50 mW) with irradiation times of 150, 250, 500, and 1000 sec, respectively, and 3, 5, 10, 30, and 50 J at 830 nm (CW, 400 mW) for 19, 31, 63, 188, and 313 sec, respectively (Table 1). The area of exposure was held constant, and covered the entire area of the colonies on the culture plates. The control plates were not irradiated.
The irradiation times were calculated in Table 1 by taking into account two points: first, in the continuous mode the device emits two continuous pulses with a 5% delay leading to a 90% CW (Fig. 2), and the second, 50% attenuation of laser power caused by passing through glassy door of the plates. By selecting power and energy density on the control panel of the device, an automatically calculated time was shown on the monitor. Taking into account the 90% CW emission of the BTL laser device and, 50% attenuation of laser power caused by passing through glassy door of the plates, we changed the time duration to slightly more than two times the automatically calculated time duration shown on the monitor of the BTL device.
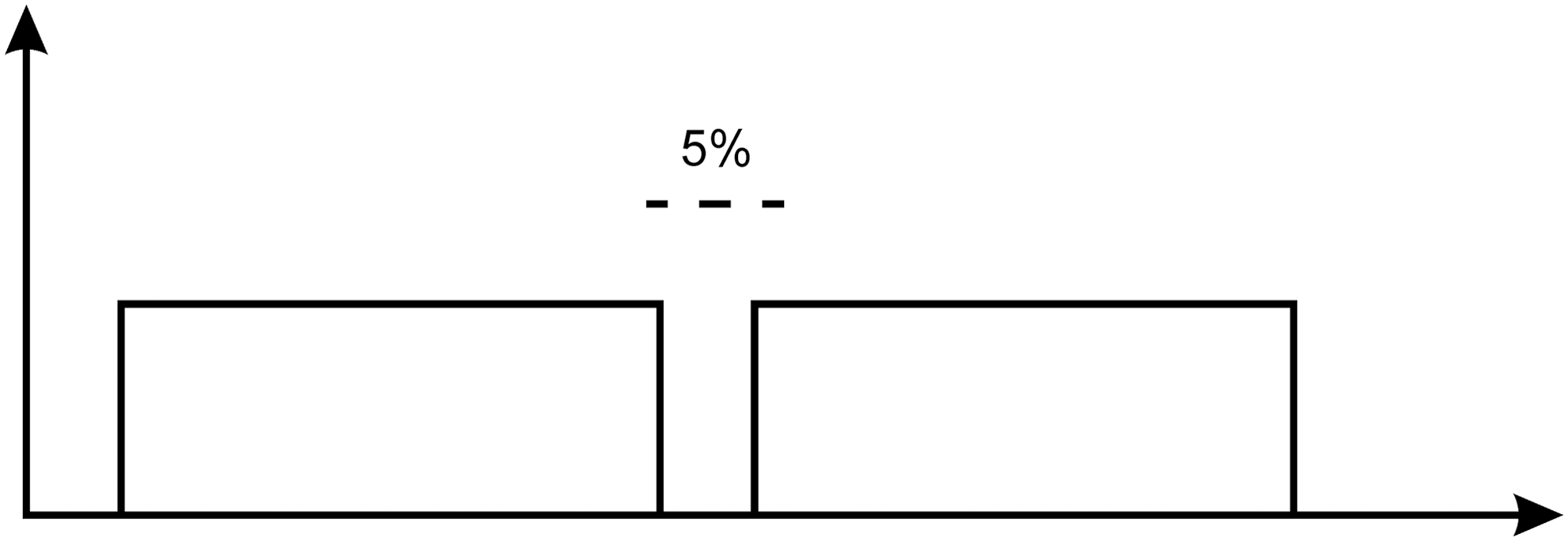
Continuous wave emission of BTL device.
In vitro study
The microscopic and macroscopic morphological and physiological characteristics of the C. albicans isolates were assessed in vitro using semiquantitative and qualitative methods. The macroscopic morphology was determined by directly observing the surfaces of the colonies. The microscopic morphology was investigated by counting the number of yeast cells, using a hemocytometer and evaluating the presence of true mycelium, pseudohyphae, blastospores, and chlamydospores after transferring the yeast to CMA containing Tween 80, and incubating the plates at room temperature for 3 days. The API 20C AUX system was used to assay the assimilation of carbohydrates after 72 h of incubation at 25–30°C.
Quantification of turbidimetric growth
The turbidimetric growth curves of the control and laser-irradiated C. albicans isolates were determined using the procedure described previously. 20 –23 Briefly, the inocula were prepared by diluting an overnight culture with 0.9% NaCl to 1–5×106 CFU/mL. The yeast suspensions were then further diluted in RPMI-1640 (with L-glutamine, without sodium bicarbonate) (Sigma, USA) supplemented with 0.165 M morpholinepropane sulfonic acid (MOPS) to give a final inoculum between 0.5×103 and 2.5×103 CFU/mL. Fungal inocula (100 μL) were added to each well of a sterile 96 well flat-bottomed microtiter plate containing RPMI-1640 (100 μL/well). Inoculations were performed in duplicate for each isolate, and two wells contained only media (background control). After inoculation, the microtiter plates were agitated for 15 sec and incubated at 35–37°C inside a plate reader (Rosys Anthos HT3; Anthos Labtec Instruments GmbH, Salzburg, Austria) for 72 h under ambient conditions. The optical density at 405 nm (OD405) was automatically recorded for each well every 30 min without shaking. The changes in OD over time were used to generate growth curves. The background ODs (the ODs of growth control wells) were subtracted from the ODs of the inoculated wells, and then the growth curve was plotted against time.
In vivo experiments
A total of 110 female inbred BALB/c mice (Institute Pasteur, Tehran, Iran), 8–10 weeks old, weighing 20–22 g were randomized into 10 groups of 11 mice (1 control and 9 treatment groups). The experimental protocols were approved by the ethical review board of the faculty of medicine and medical sciences, Islamic Azad University, Ardabil branch, Iran. The mice were infected by injection of a 0.1 mL inoculum containing 5×105 cells (conidial suspension of C. albicans), corresponding to 10% survival, into the lateral tail vein. In the control groups, the mice were infected with untreated colonies. The treatment groups were infected with laser-irradiated C. albicans isolates (3, 5, 10, and 20 J at 685 nm and 3, 5, 10, 30, and 50 J at 830 nm). Then, the conidia were harvested in 10 mL of sterile phosphate-buffered saline (PBS), and the number of conidia was counted in a hemocytometer. After adjustment to the required concentration, the inocula (the conidial suspensions) were stored overnight at 4°C. The animals were housed under standard conditions, with drink and food supplied ad libitum. The infected mice were examined at least three times daily. The clinical inspections were carried out in order to ensure there are no cases of desiccation, torticollis, staggering, high weight loss (a decrease of 15% or more within 24 h or 20% or more within 48 h), or body temperature drop to <33°C. Mice demonstrating these signs of disease were humanely euthanized. On day 21 post-infection, surviving mice were humanely euthanized with isofloran anesthesia and internal organs were harvested. The survival in days post-infection was recorded and used as end-point.
Data analysis
All data analyses were performed using the software package GraphPad Prism, version 5.0, for Windows (GraphPad Software, San Diego, CA). Mortality data were analyzed using the log rank test. The significance of the differences between groups was determined using one way ANOVA followed by Bonferroni's multiple comparison test. Statistical significance was defined as a p value of 0.05 (two tailed).
Results
Our results indicate that all control and laser-irradiated C. albicans isolates grew well after 24 h of incubation at 35–37°C. There were no significant differences in macroscopic or microscopic morphology, chlamydospore production, or the patterns of carbohydrate assimilation between the treated and control groups. However, the area under the curve and the turbidimetric growth characteristics of C. albicans were significantly different between the control and laser-treated groups for both wavelengths (p value≤0.05). Laser energies ≥10 J resulted in significant growth inhibition compared with the growth of the unirradiated control group. A representative example is shown in Fig. 2, where the mean area under the curve for the unirradiated control group was 1.711, compared with 0.629 and 0.673 for the isolates irradiated with 10 J at 685 and 830 nm, respectively. In addition, the control cultures of C. albicans isolates reached the stationary phase at 22 h; however, the logarithmic phase of the laser-irradiated groups was shorter, and the groups reached the stationery phase at 18 h (4 h earlier than the control group) (Fig. 3). This inhibitory effect on growth was not significantly different between the groups irradiated with energies ≥10 J (either 10 or 20 J at 685 nm compared with each other or with 10, 30, and 50 J at 830 nm). There were no significant differences in the susceptibility profiles of the laser-treated and control isolates based on the in vitro minimum inhibitory concentrations (MICs) of antifungals (data not shown).

Kinetic growth curves of Candida albicans in low-level laser treated (10 J at 685 and 830 nm) and control groups. Changes are shown based on optical density (OD) over time. Curve fitting of data points were performed using nonlinear regression (sigmoidal curve), p value≤0.05.
The survival curves for the control groups indicated a mortality of 100% and a median survival time of 2 days, as shown in Fig. 2. The maximum energy (20 J, 50 mW) at 685 nm LLLI resulted in 82% survival, compared with 91% when mice were infected with the C. albicans isolates that were already exposed to 50 J at 830 nm/400 mW (Fig. 4). There were no significant differences in the rate of survival between the groups infected with the C. albicans isolates exposed to energies ≥10 J (either 10 or 20 J at 685 nm compared with each other or with 10, 30, and 50 J at 830 nm). In contrast, the graph clearly shows statistically significant differences in the survival rates between the control and laser-treated isolates irradiated with energies above 10 J at both wavelengths. Differences were found in both the median survival time and the survival rate.

Survival curves of various low level diode laser irradiated Candida Albicans isolates: 3, 5, 10, and 20 J at 685 nm–50 mW; and 3, 5, 10, 30, and 50 J at 830 nm–400 mW. Control group received no treatment. For all groups, n=11.
Discussion
In the present study, we evaluated in detail the effects of LLLI on the in vitro growth characteristics and in vivo pathogenicity of C. albicans in a murine model of disseminated candidiasis in the absence of a photosensitizer. C. albicans cultures were exposed to common clinically applied energies of LLLI: 3, 5, 10, and 20 J at 685 nm/50 mW and 3, 5, 10, 30, and 50 J at 830 nm/400 mW. Significant effects were observed in vitro on the turbidimetric growth kinetics following LLLI, and in vivo on the median survival time and survival rates of mice (p value≤0.05). In both the in vitro and in vivo studies, the statistically significant effects started at the energy of 10 J for irradiation at both wavelengths. Therefore, this energy could be considered a threshold for clinical investigation and it can be proposed that a direct laser-based approach without using a photosensitizing agent can significantly reduce the pathogenicity of C. albicans.
In our in vitro study, there were no significant changes in the macroscopic morphology or biochemical phenotype of the C. albicans in the laser-irradiated groups compared with the unirradiated controls. However, a significant decrease in the growth rates over 24 h was observed. These effects were in agreement with the decreasing pathogenicity of irradiated isolates in survival using a murine model. The mice infected with colonies irradiated with energies ≥10 J using 685 and 830 nm lasers survived longer during the observation period, whereas all controls died by day 7. It can be hypothesized that LLLI affects endogenous chromophores present in the fungi, resulting in the inhibition of adherence or dimorphism, 24,25 thus reducing the virulence of C. albicans. 26,27
One possible limitation of the experimental design used in the current study is that the effects were observed in clinical isolates instead of a standard reference strain. However, the use of four different clinical isolates helped us to reach firmer conclusions, despite the possible risk of nonstandard behaviors. 28 There are several reports regarding the use of photodynamic-based eradication strategies with the application of different photosensitizers to treat infections with a variety of microbial species. 26 Teichert et al. indicated that the effect of methylene blue in combination with the use of a diode laser on buccal candidiasis in immunosuppressed mice was directly proportional to the photosensitizer concentration. 29 Junqueira et al. evaluated the effects of photodynamic therapy on buccal candidiasis in rats and verified that the rats treated with a laser and methylene blue developed a greater number of discrete candidiasis lesions than the control groups did. 30 Therefore, further research is required to determine the photodynamic effects of LLLT on C. albicans in the absence of a photosensitizer.
Many in vitro and in vivo investigations have evaluated new methods to reduce the number of C. albicans, including microwave disinfection, ozone exposure, and photodynamic antimicrobial therapy. 29 –35 In such studies, the in vitro effects of 685 nm laser radiation on the viability of different species of Candida genus have been investigated in the presence of photosensitizers, 13,34,36 and it was concluded that it is possible to destroy C. albicans colonies with light from a low-power laser when the yeast is sensitized with low concentrations of various dyes. 13 A few studies have focused on the effects of LLLT on the growth of bacterial colonies in the absence of a photosensitizer. One such study investigated the effects of different wavelengths of low level laser (630, 660, 810, and 905 nm) on various genus of bacteria. 10,11 Interactions were found among the species, wavelength, and radiant exposure in the overall effects on bacterial growth. A wavelength of 630 nm appeared to be most commonly associated with the inhibition of bacterial growth. The laser doses that we used in our study were selected based on the results of the study by Nussbaum et al., 11 who evaluated the bactericidal capacity of LLLI at various wavelengths and observed beneficial bactericidal effects. Despite the promising results achieved in vitro, the biostimulatory effects of LLLI were not remarkable.
Similar to our findings, laser energy has been shown to have the potential to eliminate T. rubrum, 12 C. albicans, and A. niger, 14 suggesting that a direct laser-based approach without using a photosensitizing agent can eradicate superficial fungal infections. Saltarelli et al. investigated the responses of four strains of C. albicans to light irradiation, and concluded that the light-induced changes in the morphology of all strains; however, the viability was not affected. 37 Maver-Biscanin et al. studied the effects of diode laser irradiation on denture stomatitis, and a fungicidal effect was observed in the laser-treated groups. These researchers found that C. albicans can be destroyed by light from a semiconductor diode laser in the absence of a photosensitizer. 38,39 They compared the significant effects of two wavelengths of LLLI and observed that the fungicidal effect was greater for 830 nm, whereas the wavelength of 685 nm had a greater anti-inflammatory effect. 38 In our study, we did not observe such a significant difference between these two wavelengths.
Regarding the possible mechanism of action of LLLT, evidence is accumulating that LLLT influences both microorganisms and the cells of the target organ. 40,41 A recent review by Bjordal et al. indicated that LLLT modulates the inflammatory process by exerting neurophysiological, local anti-inflammatory, and biochemical effects in addition to cellular and soft tissue effects. The light delivered by lasers has a stimulatory effect on human lymphocytes, in particular on initially low-activated lymphocytes. 42 In addition, the effects of the co-stimulation of the immune response with adenosine triphosphate (ATP) production in mitochondria should be considered. However, all these results apply to the evaluation of the effects of LLLT on C. albicans in in vivo or in a clinical setting, where the laser light is absorbed by both C. albicans and the host immune cells at the same time. In our study, laser light was absorbed only by C. albicans in our in vitro setting, and, therefore, the effects on the immune system should be negligible.
With regard to the intrinsic mechanism of action affecting microorganisms, in a recent article it was reported that laser light modulates transmembrane convection, which may result in the destruction of the treated cells. This effect is explained by the interplay between the expansion and contraction of the intracellular water volume and fluidity, which generates bidirectional flow across the cell membrane. 43
C. albicans is the main etiological agent of superficial and systemic candidiasis. 1 Despite the use of antifungal drugs to treat vulvovaginal and oral candidiasis, infections are often re-established. 44 This behavior is of great importance, as the rate of recurrence is high, and an optimal treatment modality has not yet been found. Other problems with classical antifungal agents include their side effects. These side effects may be exacerbated after long-term use, as indicated in relapse prevention or in the treatment of recurrent infections. Long-term use may also encourage the development of resistant strains.
There is growing interest in new approaches, including photodynamic antimicrobial therapy with various wavelengths of laser light. However, there is still a need for safe, efficient, easily performed and cost-effective treatment methods without the disadvantages of resistance to topical or systemic antimycotic agents. Because the lesions caused by superficial candidiasis are readily accessible to laser beams and because, unlike treatment with antiseptics or antibiotics, there is no need to maintain high drug concentrations in the infected area, it can be concluded that the concomitant use of LLLI could be useful in the management of recurrent superficial or cutaneous infections caused by resistant C. albicans.
Although more research utilizing C. albicans reference strains is needed to support our conclusions, our in vitro and in vivo findings indicate that LLLI has potential beneficial uses in the treatment of fungal infections. LLLI affects C. albicans by reducing its turbidimetric growth rate over time, which is in line with our in vivo results indicating a significant decrease (p value≤0.05) in the pathogenicity of the irradiated isolates.
Conclusions
Translating our data into the clinical setting, it can be concluded that a direct laser-based approach without using a photosensitizing agent may be a promising novel treatment approach for superficial and mucocutaneous C. albicans infections.
Footnotes
Acknowledgments
The work of Dr. Seyedmousavi was supported by a research grant from the Faculty of Medicine and Medical Sciences, Islamic Azad University, Ardabil Branch, Ardabil, Iran. The authors thank Ms. Maliheh Salami for her technical assistance with the laser irradiation. The authors also thank Afshan Shirkavand for her invaluable contribution to laser energy calculations.
Author Disclosure Statement
No competing financial interests exist.