
Abstract
Objective:
To investigate the effects of photobiomodulation therapy (PBMT) at 660 and 810 nm on amyloid-beta (Aβ)42-induced toxicity in differentiated SH-SY5Y cells and to assess its impact on Aβ42 accumulation and cholinergic neurotransmission.
Background:
Alzheimer’s disease (AD) is characterized by the accumulation of Aβ peptides, leading to neurodegeneration, cholinergic deficit, and cognitive decline. PBMT has emerged as a potential therapeutic approach to mitigate Aβ-induced toxicity and enhance cholinergic function.
Methods:
Differentiated neurons were treated with 1 μM Aβ42 for 1 day, followed by daily PBMT at wavelengths of 660 and 810 nm for 7 days. Treatments used LEDs emitting continuous wave light at a power density of 5 mW/cm2 for 10 min daily to achieve an energy density of 3 J/cm2.
Results:
Differentiated SH-SY5Y cells exhibited increased Aβ42 aggregation, neurite retraction, and reduced cell viability. PBMT at 810 nm significantly mitigated the Aβ42-induced toxicity in these cells, as evidenced by reduced Aβ42 aggregation, neurite retraction, and improved cell viability and neuronal morphology. Notably, this treatment also restored acetylcholine levels in the neurons exposed to Aβ42.
Conclusions:
PBMT at 810 nm effectively reduces Aβ42-induced toxicity and supports neuronal survival, highlighting its neuroprotective effects on cholinergic neurons. By shedding light on the impact of low-level light therapy on Aβ42 accumulation and cellular processes. These findings advocate for further research to elucidate the mechanisms of PBMT and validate its clinical relevance in AD management.
Introduction
Alzheimer’s disease (AD) is a progressive neurodegenerative disorder characterized by the accumulation of amyloid-beta (Aβ), particularly the soluble Aβ42 peptide. This accumulation, driven by the activity of secretases cleaving the amyloid precursor protein (APP), is a critical event in AD pathogenesis. 1,2 Amyloid oligomers, the lower-molecular-weight entities of these proteins, are recognized as primary pathogenic species, 3,4 initiating early intracellular Aβ accumulation that leads to cell loss and various pathological changes, including neurite beading and fragmentation. 5 These changes particularly impair cholinergic neurons, resulting in decreased acetylcholine levels and compromised neurotransmission, crucial for cognitive function. 6 –8
In vitro models, especially SH-SY5Y cells, have become invaluable in AD research owing to their ability to mimic neuronal behavior and respond to pathological stimuli such as Aβ. 9 –11 By differentiating these cells into a cholinergic phenotype using retinoic acid (RA) and brain-derived neurotrophic factor (BDNF), researchers can explore the impacts of AD-related proteins on neuronal survival and function. 12 –15 This approach provides insights into the disease’s progression and potential therapeutic interventions.
The therapeutic potential of red and near-infrared (NIR) light has been increasingly explored for its efficacy in ameliorating neuron loss in Parkinson’s disease models and reducing Aβ and tau proteins in AD, both in vitro and in vivo. 16 Photobiomodulation therapy (PBMT), a noninvasive and nonthermal approach, uses low-power light sources such as lasers or LEDs in the red and NIR spectra to stimulate cellular self-repair, enhance energy production, and the prevention of injuries. 17 –19 PBMT has shown promise in various studies; for instance, 670 nm laser irradiation has been demonstrated to clear Aβ42 aggregates albeit at the expense of cellular adenosine triphosphate (ATP) in human neuroblastoma SH-EP cells. 20 Further, 810 nm laser irradiation has effectively reduced reactive oxygen species (ROS) levels in primary cortical neurons, thus protecting against oxidative stress and aiding neurocognitive recovery in conditions of Aβ-mediated neuroinflammation. 21,22
Building upon these insights, our prior research showed that red (660 nm) and NIR (810 nm) wavelengths at energy densities of 1, 3, and 5 J/cm2 could modulate biochemical processes in neural cells, protecting neurons from Aβ42-induced neurotoxicity in a biphasic manner. Treatment with PBMT following Aβ42 exposure could attenuate intracellular ROS generation and calcium influx, potentially alleviating cell death and facilitating neurite outgrowth through enhanced mitochondrial activity. 23 However, the role of PBMT in modulating cholinergic neurons to mitigate Aβ accumulation within neural cells remains not fully understood.
This study aims to address these gaps by investigating the impact of 660 and 810 nm PBMT on in vitro model for AD. We hypothesize that PBMT at these wavelengths will not only reduce Aβ42 peptide accumulation but also bolster cellular defenses against AD pathology. By elucidating the effects of PBMT on Aβ accumulation and associated cellular responses, we hope to uncover new therapeutic mechanisms that could potentially reverse aspects of AD pathology, offering hope for more effective treatments in the future.
Materials and Methods
Chemicals and reagents
Cell culture–associated reagents were purchased from Gibco/Invitrogen: Dulbecco’s modified Eagle’s medium (DMEM), Ham’s F-12 Nutrient Mix, 0.25% Trypsin–ethylenediaminetetraacetic acid (EDTA) solution, antibiotic/antimycotic, fetal bovine serum (FBS), and Trypan blue stain. Methyl thiazolyl diphenyl-tetrazolium (MTT) cell viability reagent was purchased from Gibco (Invitrogen Corporation). Human beta amyloid 1-42 (Aβ42) PTD recombinant protein, BDNF, RA, poly-
Cell culture
SH-SY5Y neuroblastoma cells were maintained at 37°C in a humidified atmosphere of 5% CO2. Cells were grown in a mixture medium of 1:1 DMEM and Ham’s F12 supplemented with 10% FBS and 1% antibiotic/antimycotic. Culture medium was aspirated every 3 days, and cells were passaged once they reached 80% confluence using 0.25% Trypsin-EDTA solution.
Differentiation protocol
A differentiation protocol was developed by Forster et al.
10
Briefly, the cells were seeded onto 6-well culture plates coated with collagen type I and poly-
Preparation of Aβ42 oligomer
Aβ42 oligomer was prepared according to the protocol outlined by Stine et al. 24 with slight modification. First, a solution of 1 mg/mL Aβ42 peptide in HFIP was prepared and incubated at room temperature (RT) for 1 h with occasional vortex at moderate speed, followed with a 10 min-sonication in a water bath sonicator. The Aβ-HFIP solution was then dried with nitrogen gas to obtain a clear thin peptide film at the bottom of the tubes. Dried peptide films were stored at −20°C until use. Before each experiment, the peptide films were thawed to RT, resuspended in 100% anhydrous dimethyl sulfoxide (DMSO; Sigma-Aldrich) to a final concentration of 5 mM, vortexed thoroughly, and sonicated for 10 min. The Aβ42 solution was further diluted in ice-cooled DMEM Phenol red-free medium to acquire a final concentration of 1 μM.
Photobiomodulation treatment
An array of 30 aluminum gallium indium phosphide (InGaAlP) LEDs at 660 nm and aluminum gallium arsenide (AlGaAs) LEDs at 810 nm (both from Marubeni) was used. The spot size was 5 × 5 mm2, ensuring uniform delivery of light energy to all target cells. Detailed information on LED parameters, such as full-width at half-maximum and electrical power, is described further in Table 1, and variations in LED intensity across wavelengths are shown in Supplementary Table S1. The PBMT used LEDs at both 660 and 810 nm, with the beam focused from the LEDs’ planar array onto the bottom of the culture plate at a distance of 3 cm, covering the surface area of the cell culture plate (Supplementary Fig. S1). Irradiation was applied to the irradiating plane, delivering a constant power density of 5 mW/cm2 to achieve an energy density of 3 J/cm2. Energy at the bottom of the culture plate was measured using an AvaSpec-ULS2048XL EVO spectrometer (AVANTES, Spectral Technology Instrument Co., Ltd.), calibrated with a National Institute of Standards and Technology’s standard source over the specified wavelength for spectral irradiance (μW/cm2/nm).
Specifications for SMBB660D-1100 and SMBB810N-1100
To assess the neuroprotective potential of PBMT, differentiated neurons were treated with 1 μM Aβ42 for 1 day, followed by additional PBMT treatments (Fig. 1). The culture medium supplemented with 1 μM Aβ42 was changed every 2 days. PBMT involved irradiation at both 660 and 810 nm for 10 min, maintaining a power density of 5 mW/cm2 and an energy density of 3 J/cm2 at the surface of the neural culture plate. Control treatments with Aβ42 were handled similarly, except for the exposure to the LED array. This treatment protocol was based on preliminary studies that demonstrated rapid neurite retraction after 3 days of Aβ42 treatment, which served as a parameter to monitor neurotoxicity against Aβ42.

PBMT treatment protocol. Neuronal cells were incubated with Aβ42 on day 0, and the media containing Aβ42 was changed every 2 days. The PBMT treatment interval was conducted with cells receiving treatment once a day. After treatment with PBMT for 3 days, cells were assayed. The experimental groups include control (without Aβ42 and PBMT), control-treated Aβ42 (without PBMT), PBMT, and Aβ42-PBMT. Aβ, amyloid-beta; PBMT, photobiomodulation therapy.
Cell viability
Based on the determination of metabolic activity, an MTT assay was used to measure cell viability, assessed by the mitochondrial ability to metabolize MTT. Aβ42 peptide was added into cultured medium and incubated for 24 h after that the cells were treated with PBMT for 7 days. The medium was then discarded, and the cells were washed with 100 mL phosphate-buffered saline (PBS) followed by the addition of 50 μL of MTT (ThermoFisher), which measures mitochondrial activity by reducing this salt into formazan (purple) crystals. The mixture was incubated for 3 h at 37°C and 5% CO2. After incubation, 100 μL DMSO was added at RT to resuspend and solubilize the precipitate. The absorbance of supernatant was then measured at 570 nm with a microplate reader (Infinite® M200, TECAN).
Cell morphology visualization
Coomassie brilliant blue staining method for neuroblastoma cell was reported by Shutt et al. 25 This staining method was used to observe morphological changes in the organelles and cytoskeletal systems of cultured cells. Briefly, cells were fixed with 70% ethanol and stained for 20 min with a solution containing 0.02% Coomassie brilliant blue R-250 in a mixture of methanol, acetic acid, and water (46.5:7:46.5, v/v/v). After staining, the cells were rinsed several times with PBS to remove excess dye and then examined under an inverted microscope.
Fluorescence imaging for analysis of Aβ42 aggregation
To comprehend the impact of the Aβ42 peptide on neuronal cells, Aβ42 oligomers, commonly labeled with fluorescein isothiocyanate (FITC-Aβ42) at a concentration of 1 µM, were incubated in both differentiated and nondifferentiated cells. The preparation of FITC-Aβ42 was carried out in a manner similar to that used for Aβ42 oligomers, with the important stipulation that exposure to light be avoided during the process. Subsequently, the cells were incubated overnight at 37°C. Fluorescence microscopy (Cytell™ Cell Imaging System, GE Healthcare) was then used for cell imaging. Image processing and analysis were conducted using Java software (ImageJ) to evaluate the area of FITC-Aβ42 covering cell morphology.
Measurement of acetylcholine concentration
To quantify acetylcholine, we used a choline–acetylcholine quantification kit following the manufacturer’s protocol (Quimigen S.L.). In this assay, free choline is oxidized to betaine via the intermediate betaine aldehyde. This reaction generates products that react with the Choline Probe to produce color (570 nm) and fluorescence (Ex/Em 535/587 nm). Briefly, cells were cultured in a six-well plate. After the desired treatment, the medium was removed, and the cells were lysed in choline assay buffer on ice for 10 min and then centrifuged to remove debris. The lysate was directly tested, and fluorescent intensity was recorded using a microplate reader (Infinite M200, TECAN) at the appropriate excitation wavelength.
Image processing
Analysis of neurite outgrowth and neurite complexity was performed with ImageJ software according to the protocol outlined by Boulan et al. 26 and Pemberton et al. 27 with some modification. Initially, all images were prepared by optimizing phase contrast and removing the background. Subsequently, neurite outgrowth was measured by tracing all neurites using the NeuronJ toolbar.
Statistical analysis
All statistical analyses were performed with R program. Differences between the two groups were compared using the t test or Mann-Whitney U test for parametrically and nonparametrically distributed values, respectively. Serial or multiple comparisons were conducted using analysis of variance and post-hoc testing, or the Kruskal-Wallistest and post-hoc testing, for parametrically and nonparametrically distributed values, respectively. A P-value < 0.05 was considered to reflect statistical significance.
Results
Morphological changes in differentiated SH-SY5Y cells
Differentiated cells were obtained by the treatment of RA for 5 days, followed by withdrawal of serum in the culture medium and addition of BDNF for 2 days. After 2 days of BDNF treatment, we replaced the medium with medium containing 1% FBS to maintain cell survival. We observed a relative decrease in cell number after withdrawal of BDNF, which may be owing to S-phase entry and apoptotic cell death. This result suggests that variations in the culture conditions can alter cellular responses.
As shown in Fig. 2, nondifferentiated cells tended to grow in clusters and form clumps of rounded cells on top of one another (indicated by the yellow arrow). At the edges of the clusters, cells began to extend short neurites, and their proliferation rate increased rapidly (see Fig. 3). In contrast, differentiated neurons exhibit extensive neurite outgrowth that connects with adjacent cells, resulting in a loss of rounded morphology. In addition, treatment with RA/BDNF leads to a nearly homogeneous population with enhanced neuron-like characteristics compared to cells maintained in traditional culture conditions.
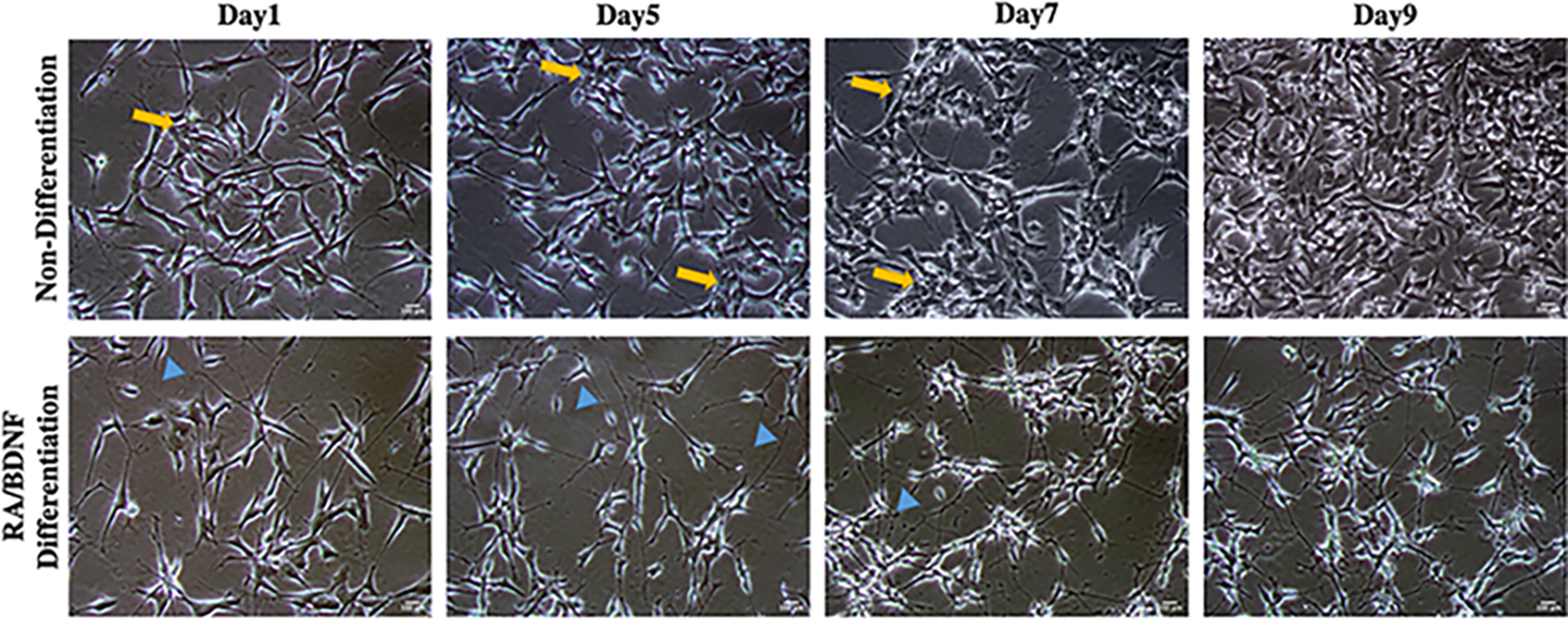
Visualization of differentiated SH-SY5Y cells. Phase contrast image with 10×. Scale bar: 100 μm. Nondifferentiated cells exhibited roundish features with few short processes (yellow arrows). Differentiated cells were more neuronal-like, extending cellular processes or neurites (blue arrowheads).
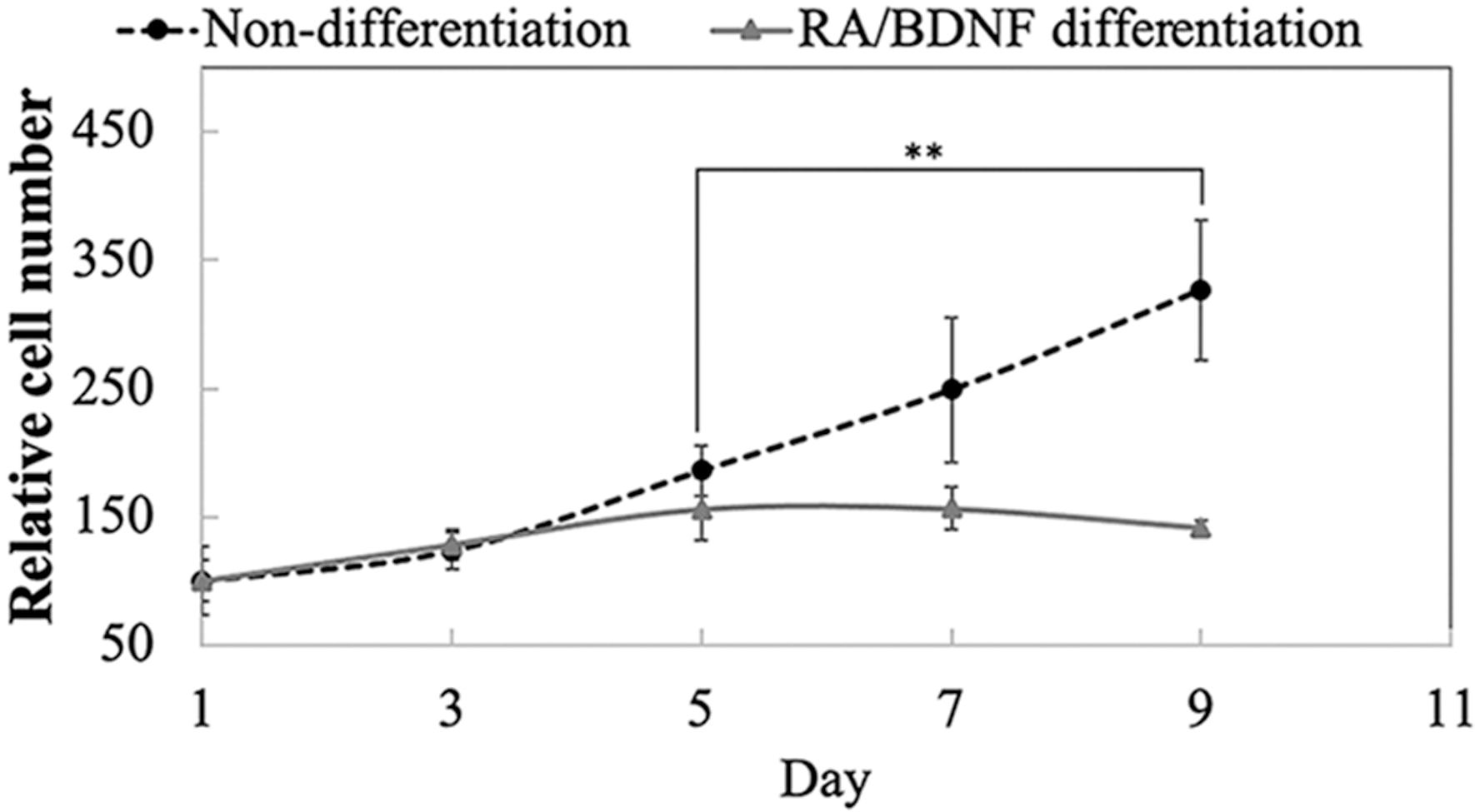
Relative cell number after 9 days of differentiation. The figure displays the mean ± SD; n = 5. **p < 0.01. SD, standard deviation.
Effect of oligomeric Aβ42 accumulation on cell differentiation
The aggregation of amyloidogenic Aβ-peptides on cell surfaces is a well-known phenomenon in cultured cells. We explored the distribution of Aβ42 using FITC-conjugated imaging (Fig. 4). Our findings reveal a pronounced accumulation of extracellular Aβ42 in a time-dependent manner in differentiated cells, whereas nondifferentiated cells displayed minimal changes over time. After a 2-day exposure to 1-μM Aβ42, differentiated cell bodies were completely enveloped by significant Aβ42 aggregates, leading to neurite shrinkage. Quantitative analysis indicated a notable increase in the area covered by FITC-Aβ42 aggregates over differentiated cells, increasing significantly to 1.86 ± 0.04, 1.62 ± 0.12, and 4.83 ± 0.97 folds after 2, 3, and 4 days of treatment, respectively. This interaction between cell bodies and Aβ42 is linked to neurite retraction and subsequent cell death, with a significant decrease in cell viability after 7 days of Aβ42 exposure (Fig. 5). These results highlight the detrimental effects of Aβ42 on cellular integrity and underscore the influence of differentiation status on cellular responses to Aβ-induced toxicity.
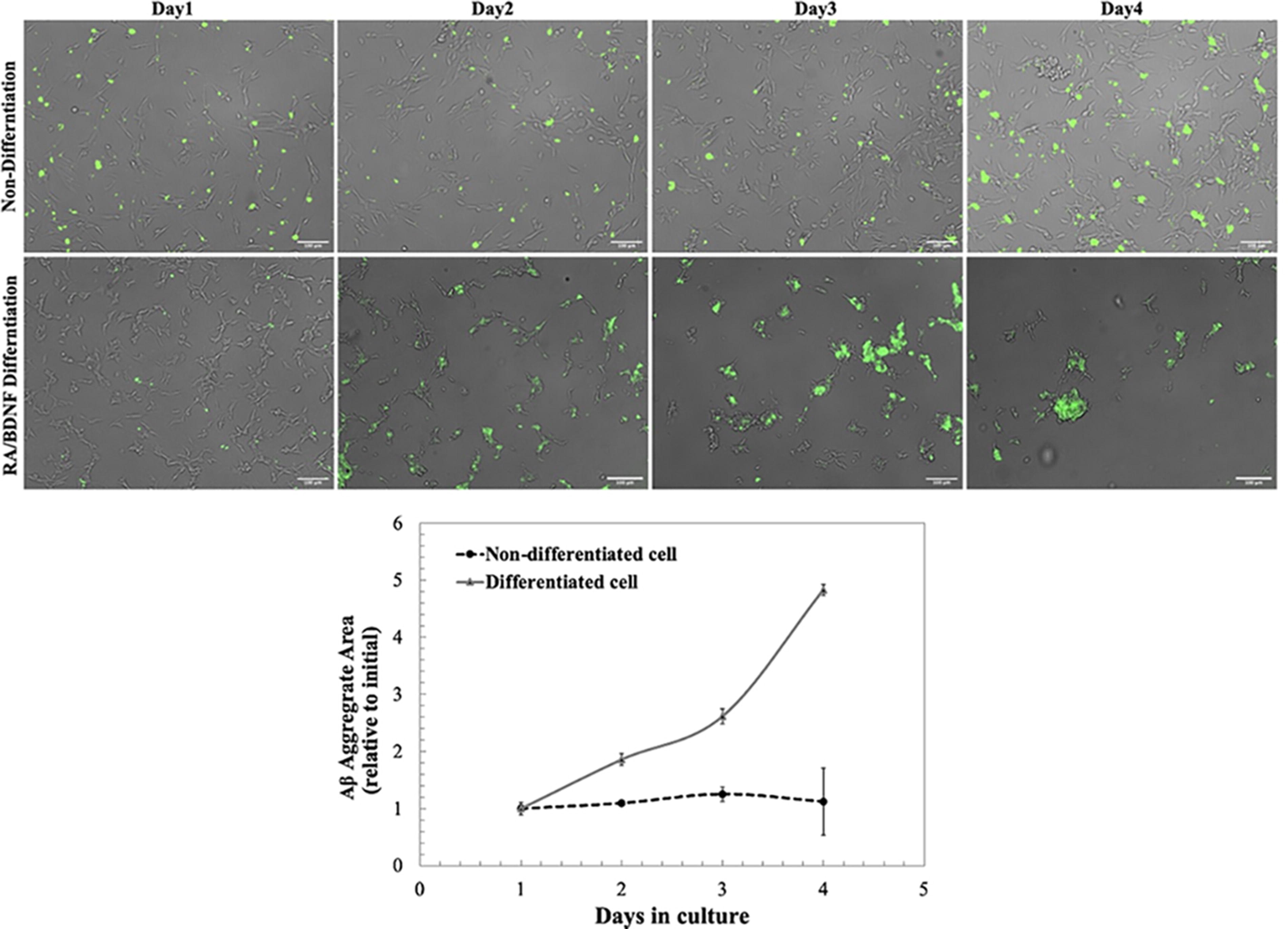
FITC-Aβ42 covered cell bodies and induced pathological changes in neural morphology after 1, 2, 3, and 4 days of incubation. Scale bars represent 100 μm. The size of FITC-Aβ42, reported as a relative value from day 1 of incubation, was determined using Fiji ImageJ. Values represent mean ± standard error; *p < 0.05. In this example, the sample size was 5, and there were two replicates for each condition. FITC, fluorescein isothiocyanate.

Cell viability was measured with the MTT test 7 days after treatment with Aβ42 peptides. The figure displays the mean ± SD; at least n = 3; *p < 0.05; #p < 0.05 vs. control. MTT, methyl thiazolyl diphenyl-tetrazolium.
PBMT mitigates Aβ42-induced toxicity in differentiated SH-SY5Y cells
Given that RA/BDNF differentiation increases the vulnerability of SH-SY5Y cells to Aβ42 aggregates and toxicity, these cells were exposed to Aβ42 for 24 h and then subjected to PBMT. This treatment was evaluated for its impact on morphological changes and cytoskeletal integrity using Coomassie brilliant blue staining. The findings demonstrate that PBMT positively influences neural functions, maintaining neurite morphology in differentiated cells amid Aβ42-induced toxicity, compared to control cells without PBMT (Fig. 6A). Viability significantly improved after 7 days of exposure to 660 and 810 nm light, reaching 48.10 ± 2.15% and 53.22 ± 0.85%, respectively, whereas in the absence of PBMT, viability decreased to 29.11 ± 2.22% (Fig. 6C).

Significant enhancements were observed in the mean length and complexity of neurites, with notable increases after treatment with 660 and 810 nm light for 3 days (Figs. 6D and F). The mean neurite length for the control group was 1.62 ± 0.28, whereas cells treated with Aβ alone exhibited a substantial decrease in neurite length, with a mean of 0.48 ± 0.23. However, when Aβ-treated cells were exposed to 660 and 810 nm light, the neurite lengths recovered to 1.43 ± 0.16 and 1.81 ± 0.34, respectively, indicating a reversal of Aβ-induced damage.
Moreover, both wavelengths of PBMT demonstrated significant efficacy in reducing the area of Aβ aggregates in an in vitro model, as evaluated by observing changes relative to initial conditions (Fig. 6B). Initial measurements showed a mean area of 1.88 ± 0.341 for Aβ aggregates, whereas PBMT at 660 and 810 nm significantly mitigated this increase, reducing the values to 0.95 ± 0.17 and 0.76 ± 0.09, respectively. After 3 days of PBMT, the 660 nm treatment reduced the mean value to 0.64 ± 0.07, and the 810 nm treatment further reduced it to 0.44 ± 0.06, whereas the control group’s condition worsened further to 3.75 ± 0.51. These results underscore the consistent and effective capability of PBMT, particularly at 810 nm, to counteract and reverse the adverse effects induced by Aβ, highlighting its potential therapeutic significance.
Impact of PBMT on acetylcholine levels in differentiated neurons exposed to Aβ42
The neurotoxic effects of Aβ42 peptides are linked with a decline in acetylcholine levels, a critical marker in AD because of the degradation of cholinergic neurons. Our study aimed to determine the impact of PBMT on acetylcholine levels following Aβ42-induced toxicity. The results indicated a significant reduction in acetylcholine levels to 2.35 ± 0.41 μM owing to Aβ42 exposure. However, following 3 days of irradiation with 810 nm light, there was a significant increase in acetylcholine levels in differentiated neurons to 4.03 ± 0.51 μM (Fig. 7). This suggests that although Aβ42-induced toxicity notably decreases acetylcholine levels, PBMT can effectively reverse this effect, highlighting its potential therapeutic benefits.
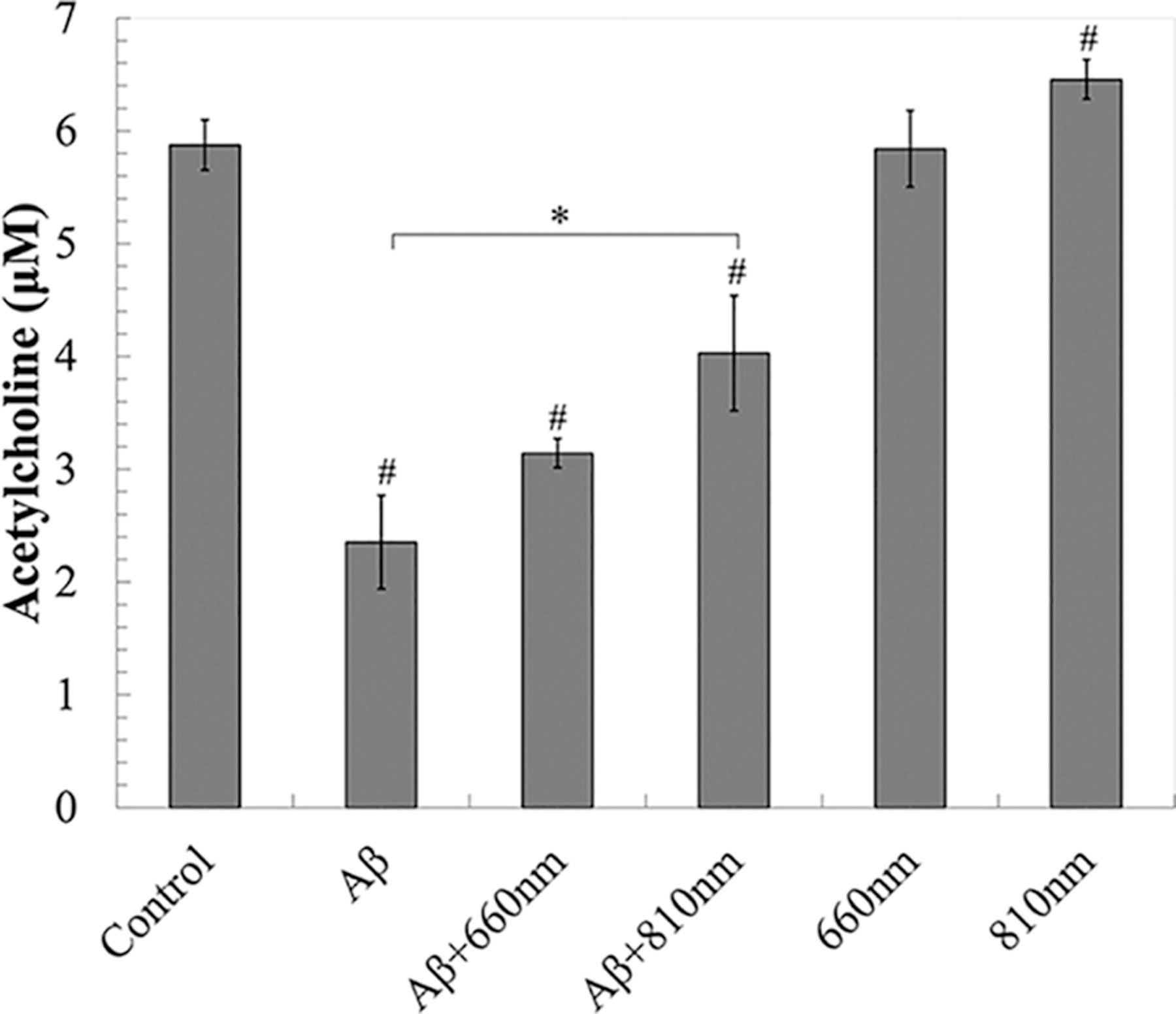
PBMT effect on acetylcholine levels in differentiated neurons after 3 days of irradiation. Data were expressed as mean ± SD; n = 3. *p < 0.05, #p < 0.05 vs. control.
Discussion
Over the past few decades, the amyloid hypothesis has been a central focus in AD research, prompting numerous studies using diverse cellular and animal models. These investigations have aimed to establish the toxic effects of Aβ peptides and explore potential alternative treatments. Several studies have reported the efficacy of PBMT in addressing Aβ-induced reactive gliosis, inflammation, tau hyperphosphorylation, and neuronal apoptosis in vivo, extending earlier findings about its protective effect in suppressing Aβ-induced oxidative stress in vitro. 20,28 –32 Despite therapeutic potential suggested by several studies on PBMT, the utility of PBMT for patients with AD remains elusive. It is unclear how red and NIR PBMT mitigates Aβ accumulation and translates into effective therapy for AD.
The molecular mechanisms underlying PBM’s effects on AD have been extensively studied, progressing from basic research to preclinical and clinical trials. Previous studies have demonstrated PBM’s ability to reduce amyloid-β aggregates in human neuroblastoma cells 20 and to alleviate Aβ-induced oxidative stress and inflammatory responses in rat primary cortical astrocytes. 33 In addition, PBM has been shown to affect microglia-mediated neurotoxicity via the tyrosine-protein kinase Src/Syk signaling pathway. 34 PBMT has also been observed to impact cell apoptosis pathways, exhibiting prosurvival effects through the protein kinase B/glycogen synthase kinase 3 beta/beta-catenin (Akt/GSK3β/β-catenin) pathway and upregulation of BDNF via CREB-dependent mechanisms. 35,36 These studies underscore the diverse cellular signaling effects of PBMT in mitigating AD pathology.
PBMT offers a promising alternative and potentially more effective method for minimizing oxidative stress crucial in the context of neurodegenerative diseases. Previous literature has demonstrated that PBMT induces the dissociation of nitrous oxide from Complex IV (cytochrome c oxidase [CCO]) in the mitochondrial electron transport chain, leading to an upregulation of CCO activity and consequently enhancing electron flow, proton gradient, and ATP production. 37 This enhancement in mitochondrial function is accompanied by an increase in mitochondrial membrane potential, triggering a transient surge in ROS generation and activation of nuclear factor–kappa B (NF-kB) in the cytoplasm. NF-kB then exerts its cytoprotective effects by translocating to the nucleus and modulating oxidative stress and neuroinflammation. Interestingly, this initial burst of ROS production induced by PBMT appears to mitigate future mitochondrial dysfunction and excessive ROS levels, indicating a hormetic response. 38
The key mechanism of PBMT involves ATP activation and the promotion of distal axonal growth and neuronal activity aligns with previous studies. 39 PBMT at 810 nm has been shown to promote neurite growth of trigeminal ganglion neurons, working at relatively low energy densities at 1 and 2 J/cm2. 40 In addition, NIR wavelengths have been shown to provide positive effects on neuronal growth in the peripheral nervous system. 41 –43 Previous studies have reported that 810 and 660 nm are optimal wavelengths for transcranial PBM, exploiting the maximal penetration depth achieved at 808 nm. 44,45 Moreover, a literature review supports that wavelengths in the far red and NIR range could be beneficial in combating AD. 46
Our investigation deepens our understanding of Aβ42 peptides’ pathological effects using an in vitro model, shedding light on the potential therapeutic role of PBMT in mitigating Aβ-induced toxicity. Our results validate the positive impact of PBMT on neuronal functions, as demonstrated by the maintenance of neurite morphology and mitochondrial activity through enhanced cell viability.
Our findings align with previous research that revealed extracellular Aβ42 oligomers cover cell bodies and whole neurites in differentiated cells, causing neurite retraction. 47 The amphipathic nature of amyloid oligomers is proposed to enhance their ability to interact with membranes, exhibiting penetration, insertion, surface coating, or potential cell-penetrating peptide activities. Previous studies reported that Aβ42 binds to the plasma membrane and forms aggregates there, which are subsequently taken up and accumulated in endocytic vesicles. This process correlates with metabolic inhibition, indicating that the formation of Aβ42 aggregates is a prerequisite for Aβ42 uptake and cytotoxicity. 48
Although Aβ can be internalized into cells through membrane disruption and receptor-mediated uptake, contributing to the intracellular accumulation of Aβ within neurons, 49 –51 we found that PBMT could reduce extracellular Aβ aggregates. This reduction in Aβ42 aggregates could be attributed to the interaction between the photons and chromophores within mitochondrial complexes, potentially enhancing mitochondrial function and reducing oxidative stress, thus mitigating Aβ-induced cellular damage. 52
Moreover, our findings revealed that Aβ42 lead to proportional decrease in acetylcholine level. Aβ peptides are involved in the regulation of acetylcholine homeostasis either through allosteric activation of acetylcholine-degrading cholinesterases or by inhibiting the high-affinity choline uptake transporter. 53 Interestingly, we observed a significant increase in acetylcholine levels following 810 nm PBMT but not 660 nm PBMT. These findings align with our previous study, which demonstrated that 810 nm PBMT following Aβ42 exposure activated mitochondrial membrane potential effectively than 660 nm. 23 The exact mechanism of how the PBMT affects cholinergic neuron remains unclear; however, a previous study has been reported that it could maintain the amount of acetylcholine receptor. 54 It could be suggested that although Aβ42 may impair the acetylcholine biogenesis process, causing neuronal loss by reducing neurite outgrowth, 810 nm PBMT offers a promising avenue for mitigating Aβ-induced neurotoxicity and presents PBMT as a preventative strategy against AD progression.
Considering the primary effect of PBMT on cellular metabolism and mitochondrial function, it is conceivable that PBMT’s therapeutic benefits extend beyond Aβ42 clearance to encompass broader cellular repair mechanisms. The restoration of neuronal morphology, preservation of neurotransmitter levels, and reduction in Aβ42 aggregates collectively underscore PBMT’s multi-faceted neuroprotective effects. 55 Our study confirms the comparative efficacy of 660 and 810 nm PBMT on neuronal viability and outgrowth, with 810 nm showing superior effects in increasing neurite length and mitigating Aβ42 burden. This substantiates the potential therapeutic impact of PBMT. The significant increase in acetylcholine levels in differentiated neurons following 810 nm PBMT further supports its role in restoring acetylcholine levels diminished by Aβ42.
Conclusions
In conclusion, our study establishes a connection between Aβ42-induced toxicity, neuronal morphology, and acetylcholine level in differentiated SH-SY5Y cells. The finding not only contributes valuable insights into the intricate mechanisms of Aβ42-induced toxicity in a cholinergic neuronal model but also highlights the potential of PBMT, particularly at 810 nm, as a therapeutic intervention to counteract AD progression. However, future studies are needed to elucidate the underlying molecular mechanisms and optimize the parameters of PBMT to enhanced efficacy for treatment of patients with AD.
Footnotes
Acknowledgments
The authors want to thank Dr. Ngamkham K. and Assoc. Prof. Hemrungrojn S., M.D., for their helpful comments and recommendations, which improve the quality of the experiments.
Authors’ Contributions
The authors confirm contribution to the article as follows: S.T., K.P., K.T., and A.K.: Conceptualization; A.K.: Funding acquisition and resources; S.T.: Methodology and data collection; S.T. and P.N.: Analysis and interpretation of results; K.T. and A.K.: Supervision; S.T., P.N., and K.T.: Validation and visualization; S.T.: Original draft article. All authors reviewed the results and approved the final version of the article.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This work was supported by Scholarship for the Development of High-Quality Research Graduates in Science and Technology Petchra Pra Jom Klao Ph.D. Research Scholarship (KMUTT-NSTDA) and National Research Council of Thailand (NRCT).
Supplementary Material
Supplementary Figure S1
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.