
Abstract
Background:
Adipose tissue is the main energy storage tissue in the body. Its catabolic and anabolic responses depend on several factors, such as nutritional status, metabolic profile, and hormonal signaling. There are few studies addressing the effects of laser photobiomodulation (PBM) on adipose tissue and results are controversial.
Objective:
Our purpose was to investigate the metabolic effects of PBM on adipose tissue from Wistar rats supplemented or not with caffeine.
Materials and methods:
Wistar rats were divided into four groups: control (CTL), laser-treated [CTL (L)], caffeine (CAF), and caffeine+PBM [CAF (L)]. Blood was extracted for quantification of triglyceride and cholesterol levels and white adipose tissues were collected for analysis. We evaluated gene expression in the adipose tissue for the leptin receptor, lipase-sensitive hormone, tumor necrosis factor alpha, and beta adrenergic receptor.
Results:
We demonstrated that the low-level laser irradiation was able to increase the feed intake of the animals and the relative mass of the adipose tissue in the CTL (L) group compared with CTL. Laser treatment also increases serum triglycerides [CTL = 46.99 ± 5.87; CTL (L) = 57.46 ± 14.38; CAF = 43.98 ± 5.17; and CAF (L) = 56.9 ± 6.12; p = 0.007] and total cholesterol (CTL = 70.62 ± 6.80; CTL (L) = 79.41 ± 13.07; CAF = 71.01 ± 5.52; and CAF (L) = 79.23 ± 6.881; p = 0.003).
Conclusions:
Laser PBM decreased gene expression of the studied genes in the adipose tissue, indicating that PBM is able to block the catabolic responses of this tissue. Interestingly, the CAF (L) and CAF animals presented the same CLT (L) phenotype, however, without increasing the feed intake and the relative weight of the adipose tissue. The description of these phenomena opens a new perspective for the study of the action of low-level laser in adipose tissue.
Introduction
The individual body weight is dependent on the mass distribution of different tissue types. Among several different tissues, adipose tissue represents ∼8–21% of men's body weight and 21–33% of women's body weight. 1 This tissue is composed primarily of adipocytes and performs a number of metabolic and endocrine activities. 2 In addition to storing energy in the form of triacylglycerol (TAG), 3 it contains neuronal cells, 4 vascular stroma, 5 and immune system components. 6 Adipose tissue not only responds to signals from the endocrine system and the nervous system, but also synthesizes and secretes hormones with important endocrine functions such as leptin, 7 cytokines, 8 adiponectin, 9 and resistin. 10
The fat adipocytes can be released into the bloodstream to be used as an energy source by the body through lipolysis. Lipolysis is the catalitical pathway responsible for the TAG catabolism stored mainly in adipose tissue in the form of lipid droplets 11 in response to beta adrenergic stimulation. 12 The hormone-sensitive lipase (HSL) promotes an hydrolytic cleavage of TAG and generates free fatty acids in the bloodstream that will later be used as a source of energy, essential precursors for the synthesis of membrane lipids or mediators in cellular signaling processes. 13
Caffeine is a drug with several stimulating properties of the central nervous system widely ingested in the world, present in >60 types of plants, including coffee beans and leaves of various types of tea. 14,15 The main mechanism underlying caffeine psychostimulation is adenosine receptor antagonism, 16 activating sympathetic nervous system responses, 17 and favoring various lipolytic responses. 18 –20
Low-level laser stimulation currently included in the photobiomodulation (PBM) group of effects leads to tissue regeneration of wounds, 21 resolution of inflammation, 22 photodynamic therapy of infections, 23 among others representing the majority of effects promoted by laser irradiation. Increases in cell proliferation also depend very much on the activation of anabolic factors to increase in quantity the cellular machinery. 24
Studies related to lipolytic (catabolic) responses in adipose tissue have been demonstrated. Neira et al. 25 verified the effect of low-level laser in lipoplasty/liposuction procedures. Twelve women subjected to lipectomy were irradiated with a red laser (635 nm) using 10 mW of power at power densities of 1.2 J/cm2 in 2 min, 2.4 J/cm2 in 4 min, and 3.6 J/cm2 in 6 min. The laser irradiation responses in the esthetic procedure showed that after 4 min of irradiation, there was ∼80% of fatty acid release from adipose tissue and 99% when laser was applied for 6 min.
The authors conclude that laser irradiation during the esthetic procedure can be used as a complement to improve the responses obtained in lipoplasty. In contrast, Brown et al. 26 did not observe consistent changes in adipocytes in vitro after 635 nm laser irradiation using a similar 635 nm laser with different methodological procedures.
Another study conducted by Medrado et al., 27 wherein the dorsal fat of normal adult rats (brown adipose tissue) subjected to low-intensity laser irradiation applied locally through intact skin, with four different energy doses (4, 8, 12, and 16 J/cm2), demonstrates a decrease in the amount of lipid droplets in brown adipose tissue, which tended to form a unilocular cell type, indicating that laser irradiation can promote differentiation responses to the formation of white adipose tissue.
Owing to the scarcity in the literature regarding LLLT responses in adipose tissue, we decided to investigate the molecular effects caused by LLLT in adipose tissue, specifically white, and in normal adipose cells.
Methods
Animal models
Male Wistar rats, weighing ∼350 g, from the University of Mogi das Cruzes, were used. The animals were divided into four groups: control (CTL) (n = 10), animals irradiated with supplementation [CAF (L)] (n = 10), animals irradiated without supplementation [CTL (L)] (n = 10), nonirradiated animals with supplementation (CAF) (n = 10). The animals were housed in cages containing wood shavings and with appropriate ventilation. The environment had a temperature control of 22°C + 1°C, with 12-h light/dark cycles with food and water ad libitum. The experimental protocol was approved by the ethics committee of the University of Mogi das Cruzes, under the protocol number 005/2016.
Irradiation protocol
The application of the low-level laser was in direct contact with the skin in the anatomical region where the subcutaneous white adipose tissue is located (posterior aspect of the thigh, right and left side). All animals were enclosed in containment boxes (Insight, Brazil) to avoid their movement. The experimental groups were irradiated once a day, five times a week, for 4 weeks with a 904 nm with the parameters described in Table 1. The CTL group was subjected to the same enclosed procedures and placement of the laser equipment, without the application of the radiation.
Laser Parameters
Caffeine supplementation
Caffeine (Sigma Chemical Co., Germany) supplementation was used at a dose of 6 mg/kg, which has been shown in the literature to be effective in promoting metabolic effects. 21 The caffeine was diluted in distilled water to prepare a stock solution of 200 mg/mL and administered by gavage, once a day, 6 mg/kg, five times a week, for 4 weeks.
Blood samples collection
Blood from the animals was collected through the caudal section at 5 mm from the end of the tail. Approximately 500 μL of blood was collected in a 2 mL tube. Blood was centrifuged at 6000 g at 4°C to obtain the serum.
Animals tissue collection
The animals were sacrificed by the guillotine decapitation method without the use of anesthetics because, since metabolic responses in the adipose tissue were analyzed, many anesthetics commonly used in animal experimentation may interfere in some way with the cellular metabolism of this tissue. 28
All preparation of the environment followed strict control according to the standards described in the “Brazilian Guide to Good Practices for Euthanasia in Animals” (Federal Council of Veterinary Medicine—CFMV—2012). After euthanasia, the animal was cleaned with 70% ethanol (w/w). After removal, the tissues were weighed on analytical balance, separated and packed in 2 mL tubes, one empty tube and another tube containing 500 μL Trizol Reagent (Life Technologies). After conditioning, the tubes were rapidly frozen in liquid nitrogen and stored at −80°C until the moment of their use.
Triglycerides and cholesterol quantification
For quantification of serum triglyceride and cholesterol levels, the monoreagent triglyceride kits (Bioclin, Brazil) and monoreagent cholesterol (Bioclin) were used. Into 96-well (Corning) polystyrene plates were added 5 μL of the serum and 200 μL of the reagent solution in each well and incubated at 37°C for 10 min under constant stirring. The spectrophotometer was read at a wavelength of 500 nm. All reactions were normalized with white and standard reagent values.
RNA extraction
The adipose tissue was homogenized by maceration with magnetic microspheres. Samples were processed in several steps (5 min each step) until complete tissue maceration. After processing, a centrifugation was carried out for 1 min of 6000 RPM at 4°C and then the removal of the supernatant fat. Totally, 200 μL of pure chloroform was then added to the sample, shaken vigorously for 10 sec, and incubated on ice for 10 min. The samples were subjected to Trizol reagent (Thermofisher) RNA extraction protocol, to obtain purified RNA, according manufacture instructions. At the end of the extraction, the samples were eluted in 50 μL milli-Q ultrapure Nuclease-free water.
cDNA synthesis
Totally, 1 μg of extracted RNA was used for cDNA synthesis. The cDNA synthesis reaction was performed with the High Capacity cDNA Reverse Transcription kit (Applied Biosystems) using 10 μL of RNA treated with DNase, 10 × RT-buffer, 25 × dNTP mix (100 μM), 10 × RT random primers, 5 U of multiscribe reverse transcriptase (50 U/μL), 8 U of RNase Out (40 U/μL), and milli-Q water in final volume of 20 μL. The reaction was incubated in thermocycler under the following conditions: 25°C for 10 min, 37°C for 120 min, 85°C for 5 min, and 4°C. At the end of the reaction, the cDNA samples were diluted 1/10 in milli-Q DNase and RNase-free water and stored at −20°C until the time of its use.
Gene expression analysis
The SYBR Green reaction was used with the following conditions for each sample: 5 μL of sample, 5 μM of primers, 10 μÇ of Master Mix SYBR Green previously with ROX (Thermo Scientific), and ultrapure water for final volume of 20 μL. Reactions were all done in duplicates on 96-well real-time PCR plates. The quantified genes were HSL (lipeRN—F: 5′ TGCAAGTTCCCTCTTTACGG 3′; lipeRN—R: 5′ TCCAGAAGGCTTTCCAGAAG 3′), leptin receptor (leprotRN—F: 5′ TATGCTGGGATGTGCCTTG 3′; leprotRN—R: 5′ TAGGGGATGGGGGAGATG 3′), tumoral necrosis factor alpha (tnfRN—F: 5′ CGGGCTCAGAATTTCCAAC 3′; tnfRN—R: 5′ GAGTTCCGAAAGCCCATTG 3′), beta adrenergic receptor (adrb2RN—F: 5′ GCGTCTGGGCACTGAAAG 3′; adrb2RN—R: 5′ GGTGCCAGCAAGAAGTCG 3′).
All expression values obtained were normalized by expression of the endogenous 18s gene (18sRN—F: 5′ GTGCGTCGATGAAGAACG 3′; 18sRN—R: 5′ GCGTTCGAAGTGTCGATG 3′). The relative expression values will be obtained by calculating the 2−ΔΔCt method.
Statistical analysis
The data are presented as mean ± standard error of the mean. For analysis of more than two samples, one-way analysis of variance followed by Tukey's post hoc verification was applied. Statistically significant differences were considered when the p-value was ≤0.05.
Results
After a 4-week period of irradiation, as we can observe in Fig. 1A and B, there was a significant change in the total weight of the animals and no increase in feed intake in the CTL (L) group. It is possible to verify that the irradiated animals had a greater mass of white adipose tissue than the control group (p = 0.04). This difference was not observed when comparing the caffeine intake groups (irradiated or not) (Fig. 1C).

It was found whether there were variations in triglyceride levels and serum cholesterol in animals treated with laser, caffeine, or both. We could observe that there is a statistical difference between the groups, with a slight tendency in the elevations of serum levels of triglycerides and cholesterol in the irradiated groups (Fig. 2A, B).
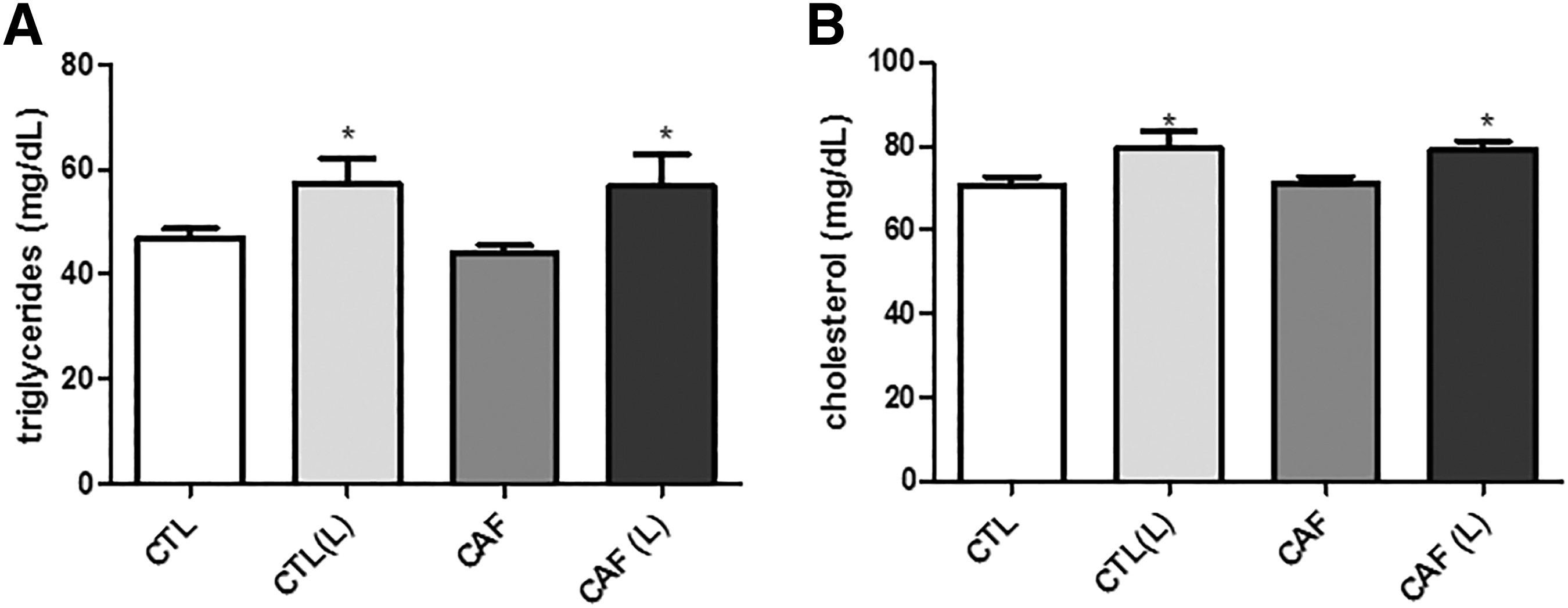
Quantification of serum levels of cholesterol and triglycerides in the experimental groups.
Regarding gene expression in the adipose tissue, it is possible to verify that the group that received infrared laser radiation and caffeine presented a decrease in the expression of all genes analyzed; however, the synergistic effect of a decrease in the gene expression was not obtained in the caffeine with laser (Fig. 3A–D). It is critical to note that these genes described indicate that catabolic responses of adipose tissue are being suppressed by infrared laser radiation. Therefore, this would be a justifiable parameter for the slight increase in the adipose tissue mass of irradiated animals.
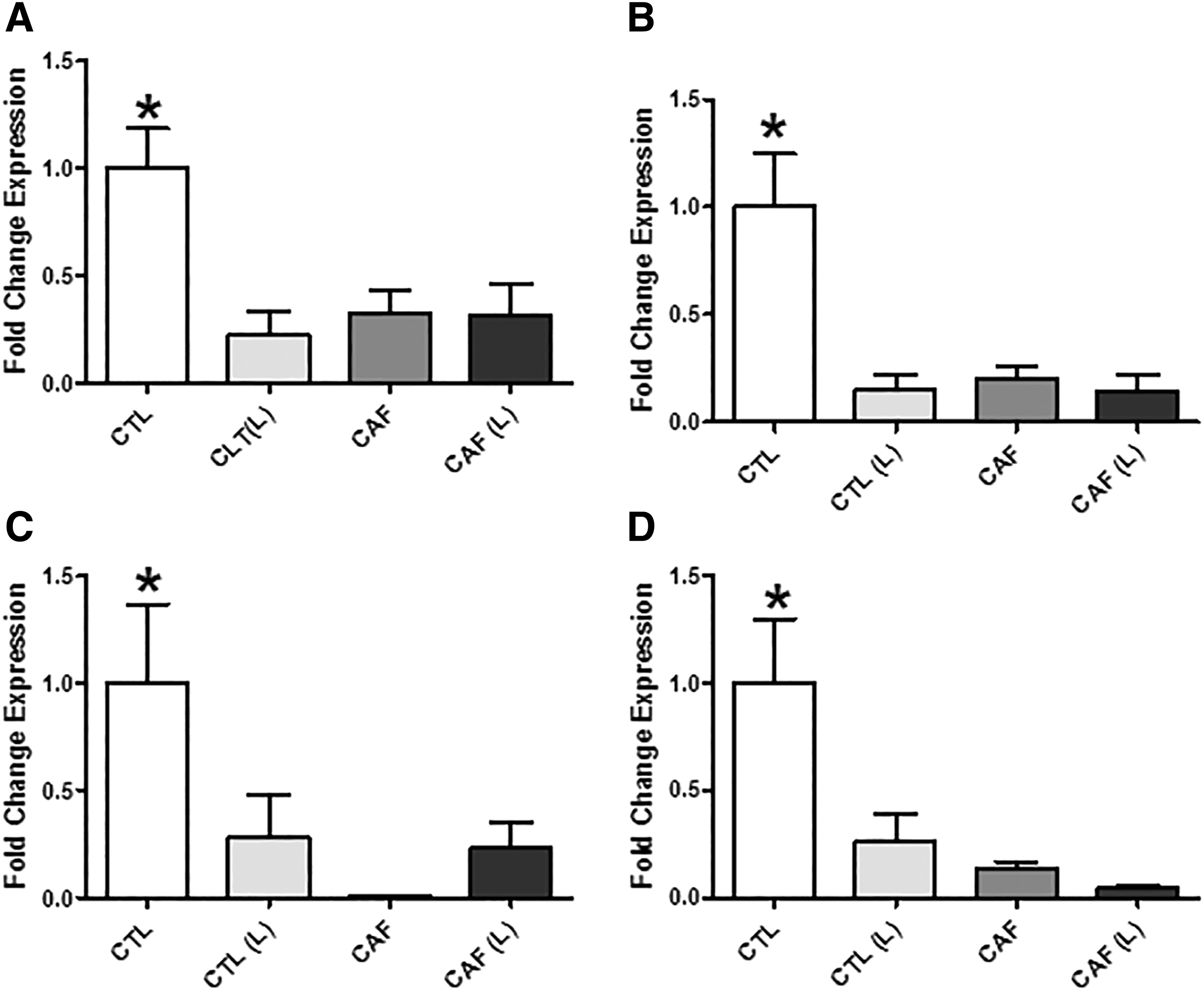
Analysis of the gene expression of adipose tissue products.
Discussion
The literature based on lipolytic responses with the use of low-level laser derives from the association of its application in lipoplasty procedures. 25,26 As there were no studies that verified the laser responses when applied to adipose tissue in a topical way (under the skin), we delineated this study to verify whether their responses had catabolic effects similar to those observed in association with lipoplasty. Curiously, we observed an inverse response, where all the results converged to an inhibition of the catabolic effects. In this context, the explanations of the choices of the models are fundamental for the understanding of the obtained results, besides that several hypotheses must be considered and discussed to give scientific clarity.
HSL is the key enzyme for the release of fatty acids from TAG adipocyte to the circulation. Both beta adrenergic and leptin receptors may be responsible for HSL activation on the fat cells. In lipolytic events, the levels of these proteins are increased. 29 –31 In addition, under anabolic response conditions due to abrupt increase in insulin signaling, TNF-α levels also increase to avoid excessive accumulation of fat in the adipocyte. 32
The variation of these components in the experimental models allows us to analyze the profile of lipolytic responses in our experimental groups. Two important results were observed: the increase in dietary intake by the irradiated group [CLT (L)] in addition to the significant increase in the relative mass of white adipose tissue in comparison with the other groups (Fig. 1B, C); (HSL, leptin receptor, TNF-α, and beta-2 adrenergic receptor) corresponding to lipolytic responses and balance of metabolism 33 in the irradiated [CTL (L)], caffeine (CAF), and irradiated+caffeine [CAF (L)] groups (Fig. 3A–D). Interestingly, although we did not observe an increase in the adipose tissue mass in the caffeine (CAF) and irradiated+caffeine groups (CAF (L) (Fig. 1C), the same expression phenotype was observed in both groups in relation to the irradiated group [CTL (L)].
As for the irradiated animals [CTL (L)], the higher feed intake (Fig. 1B) may partially explain these anticatabolic responses. The increase in the supply of nutrients without an increase in caloric expenditure necessarily implies the activation of carbohydrate and lipid anabolism. 34 This increase in consumption mediated by low-level laser stimulation may be closely related to a decrease in leptin receptor production, as observed in our results (Fig. 3D). The reduction of leptin receptor signaling in adipose tissue may promote hyperphagia, 32 increase adipogenic responses, 35,36 and decrease the expression of beta-2 adrenergic receptors, 37 corroborating the findings of our study.
According to Huan et al., 36 the responses of decreased leptin receptor production may also trigger dyslipidemia and increase hepatic insulin resistance, being a negative for metabolism if the inducing mechanism of this process occurs resistant. However, it may be that the laser will trigger this process of decreasing leptin receptor expression without necessarily causing insulin resistance, since we observed a decrease in TNF-α expression (Fig. 3C), which is a key factor to trigger hepatic insulin resistance, 32 as well as a description in the literature of a study that verified the decrease of insulin resistance in diabetic rats with low-level laser stimulation in the salivary glands. 38
We also observed an increase in the serum concentrations of total cholesterol and triglycerides in irradiated animals [CTL (L)] (Fig. 2A, B). In a study by Aquino et al., 39 it was shown that animals exposed to infrared laser (830 nm) with a total energy of 9.4 J did not have a significant increase in the lipid profile, despite having its weight increased (being consistent with our results). However, our experimental protocol used an infrared laser of 904 nm and a total energy dose of 18 J, which could be a justification of results that were inconsistent with what has already been observed in the Aquino et al. study, although our animals were not subjected to aerobic training, which, according to the author, is also responsible for promoting the loss of fat mass in the experimental models. 39
The increased concentration of these components could perhaps be explained by anabolic actions. Anabolic signals, mainly mediated by insulin and IGF1, 40 are underway for energy storage when there is a large supply of energetic substrates or protein synthesis in various cell types. 41 In addition, with an increase in the supply of energetic substrate from the glycolytic pathway, hepatocytes increase glycogen synthesis. 42 After reaching a maximal concentration, citrate 43 and acetyl-CoA 44 —the two major components for the triglycerides 45 and cholesterol 46 synthesis—are increased.
As observed in the decrease in lipolytic gene expression (Fig. 3A–D), it can be logically inferred that anabolic effects of lipid metabolism were increased, which would justify an increase in serum concentration of both total cholesterol and triglyceride. Probably, serum triglyceride levels are increased to take the excess lipids to adipose tissue, which, as observed in our results, showed an increased mass (Fig. 1A), as well as a decrease in leptin receptor expression (Fig. 3D). It is possible that this anticatabolic process is transient due to the activation of anabolic pathways. Anabolism can be justified in parallel by studies that demonstrated that the low-level laser therapy can activate signaling pathways that are clearly anabolic, such as cell proliferation in the case of wounds 47 and differentiation and proliferation of cells. 48
Our intent with caffeine was to use it as a positive control of lipolytic responses, since there are already indications of these effects in the literature. 18 –20 Caffeine supplementation (CAF) and caffeine+laser [CAF (L)] had the same gene expression profile as the laser group [CTL (L)]; however, there was no significant difference in adipose tissue mass when compared with the control group. Equivalent to the gene expression condition, in the experimental groups supplemented with caffeine, both irradiated [CAF (L)] and nonirradiated (CAF) groups presented the same phenotype of the blood lipid profile without a significant increase in the mass of white adipose tissue.
The consumption of caffeine in the dose administered to the animals (6 mg/kg) performs lipolytic events and activation of energy metabolism. 47 However, what is commonly observed in the literature is that caffeine is administered acutely rather than chronically. 49 –51 However, a study by Yu et al. 52 analyzed the effects of chronic consumption on spontaneously hypertensive Wistar rats. Curiously, sympathetic responses have decreased, suggesting the hypothesis that its chronic use may decrease its effect on metabolism and provide us with a basis for explaining our findings. 53 The adenosine receptor, when suppressed chronically, may undergo upregulation 54 –56 and decrease the levels of other receptors acting on lipolytic responses, such as adrenergic and dopaminergic receptors, 56,57 but not necessarily in the components related to the appetite. 58
Animals that were supplemented with caffeine had no increase in feed intake (Fig. 1B), probably because the initial action of caffeine promoted an anorexia that persisted throughout the treatment. The fact that the caffeine (CAF) and laser [CTL (L)] groups present the same effects at the level of gene expression, but not in the increase of feed intake and adipose tissue mass, may be that the laser and caffeine act in some similar signaling pathways competing for the same intracellular messengers, as the synergism between them in the caffeine+laser group [CTL (L)] was not observed.
In our study, we applied 18 J of total energy with laser at 904 nm wavelength. The standardization of the PBM with low-level laser and its wavelength can show different biological effects, similar to a drug therapy, where the exposure time and dose offered are defined for a specific therapeutic condition. 59,60 It has been widely demonstrated that a dose of 18 J (100 mW/cm2 power) per session of a laser at the wavelength of 810 nm (corresponding to the red laser) is capable of promoting improvements in tendinitis. 61
The variations in the wavelengths of each type of laser can also be considered as key factors for the determination of a specific dose of radiation in a therapeutic scheme. 59,61,62 In our study, we verified the variation of the gene expression of HSL, leptin receptor, beta-2 adrenergic receptor, and TNF-α in adipose tissue at wavelength 904 nm. However, since there may be different responses at different wavelengths, it is crucial to test other wavelengths and other laser parameters in future conditions to evaluate the reproducibility of our results.
All these findings corroborate with our previous study. 63 Even with all the results, this study presents some biases. The same laser parameters can have different effects when the pigmentation of the skin is darker, when the light is more absorbed in this circumstance, and there is no equivalent absorption when compared with a white skin. 64
In our experimental model, we used an albino rat (Wistar), in which we can only infer these results for conditions wherein the skin is white. Future experiments should be conducted in other animal models in which there is a darker pigmentation with the intention of verifying the reproducibility of our method. Another key point is that low-level laser PBM may have triggered direct and indirect effects on adipose tissue.
As it is well described in the literature that the low-level laser is capable of promoting cell proliferation and differentiation of mesenchymal cells, 65 it is possible that in our model there has been a differentiation of these cells into adipose tissue and, consequently, through hyperplastic responses. In addition, as we demonstrate that the catabolic efficiency of adipose tissue has decreased, it is also possible that the volume of these cells has increased through hypertrophy of adipose tissue by fat deposition. For future studies, the histological analysis of adipose tissue is necessary.
Finally, some results obtained with a minimally invasive procedure, laser acupuncture (LA), demonstrated by Tseng et al., 66 in humans showed a slight decrease in adipose tissue mass. As previously mentioned, the issue of standardization and laser protocols is very specific for each situation, so it is not always possible to perform extrapolations in relation to the obtained results.
In conclusion, at least in our study, low-level laser irradiation presented significant effects on adipose tissue metabolism. With these findings, a new field of study is opened to evaluate new effects of low-power laser on adipose tissue.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP No. 2015/06502-3).