
Abstract
Recent studies have shown that the c-Jun N-terminal kinase (JNK) signaling pathway is involved in dopaminergic neuronal degeneration, and direct blockade of JNK by specific inhibitors may prevent or effectively slow the progression of Parkinson disease (PD). Previous studies have revealed that the natural phenolic compound curcumin can reduce inflammation and oxidation, which makes it a potential therapeutic agent for neurodegenerative diseases. In this study, we investigated whether curcumin protects against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine- (MPTP) or 1-methyl-4-phenylpyridnium ion- (MPP+) induced dopaminergic neurotoxicity in C57BL/6N mice or SH-SY5Y cells by inhibiting JNK pathways both in vivo and in vitro. Curcumin treatment significantly improved behavioral deficits, and enhanced the survival of tyrosine hydroxylase-positive neurons in the substantia nigra (SN) in the MPTP-induced PD model mice. Most importantly, curcumin treatment significantly inhibited MPTP/MPP+-induced phosphorylation of JNK1/2 and c-Jun, and cleaved caspase-3. Our study suggests that the neuroprotective effect of curcumin is not related simply to its antiinflammatory and antioxidant properties, but involves other mechanisms, particularly by targeting the JNK pathways.
Introduction
The natural phenolic compound curcumin, isolated from the roots of Curcuma longa (Zingiberaceae) and commonly used as a spice, is well documented for its medicinal properties in the traditional Indian and Chinese systems of medicine. 7 Several epidemiological, clinical, and animal studies have shown that curcumin has a variety of pharmacological activities, including antiinflammatory, antioxidant, anticarcinogenic, and wound-healing effects. 8 –11 In recent years, the potential therapeutic value of curcumin for neurodegenerative diseases, such as Alzheimer disease (AD) and PD, has been increasingly recognized. 12 –14 For example, curcumin has been found to inhibit the formation of amyloid β oligomers and fibrils 15 and reduce oxidative damage and amyloid burden in AD transgenic mouse brain. 16 Curcumin treatment protects against neuronal death in middle cerebral artery occlusion-induced focal cerebral ischemia in rats. 17 An in vitro study has shown that curcumin protects PC12 cells against MPP+-induced apoptosis. 18 Curcumin pretreatment protects brain mitochondria against peroxynitrite in vitro by direct detoxification and prevention of 3-nitrotyrosine formation and in vivo by elevation of total cellular glutathione levels. 19 Furthermore, curcumin has been demonstrated to protect against nigral dopaminergic cell death from 6-OHDA neurotoxicity. 20 Most recently, it has been reported that treatment of ischemic/reperfused cardiac cells with curcumin results in an inhibition of JNK phosphorylation and, hence, protects against ischemia/reperfusion injury. 21 However, whether curcumin could provide protection against dopaminergic cell apoptosis through inhibition of JNK phosphorylation has not been examined.
In the present study, we evaluated the effects of curcumin on PD neuropathology and identified its underlying mechanism. We found that curcumin inhibited JNK-mediated dopaminergic neuronal apoptosis in both an MPTP-induced PD animal model and MPP+-induced cell death in vitro. Our data suggest that inhibition of the JNK pathways by curcumin is a potential strategy for the prevention and treatment of PD.
Materials and Methods
Experimental animals and treatments
Male C57BL/6N mice (purchased from the Experimental Animal Center, China Medical University) weighing 23–25 g (10 weeks of age) were used. Mice were maintained on a standard 12-h light/dark cycle with free access to a standard diet and tap water. All procedures using animals were conducted in accordance with the care and use of medical laboratory animals (Ministry of Health PR China, 1998) and the guidelines of the laboratory animal ethical standards of China Medical University.
MPTP (Sigma, St. Louis, MO) was dissolved in 0.9% saline at a concentration of 3 mg/mL. Curcumin (Sigma) was dissolved in dimethyl sulfoxide (DMSO) at a concentration of 5 mg/mL. Mice were divided into three groups (n = 10 in each group). Group 1 was given the control vehicle; group 2 (MPTP + vehicle) was given MPTP (30 mg/kg, intraperitoneally once a day [i.p.q.d.]) for 5 days, followed by vehicle (intraperitoneally [i.p.]) for 7 days; group 3 (MPTP + curcumin) was given MPTP (30 mg/kg, i.p.q.d.) for 5 days, followed by curcumin treatment. Because the poor bioavailability of curcumin in the diet, we chose 50 mg/kg, i.p.q.d. for 7 days.
Behavioral tests
To assess general motor behavior, the open field test and traction test were applied at day 0 (before drug administration), day 5 (injection of MPTP for 5 days), and day 12 (injection of vehicle or curcumin for 7 days). The open field test was performed as described by Teng et al. 22 Briefly, a mouse was placed in the center of a circular arena (90 cm in diameter, divided into 16 squares, and surrounded by a 40-cm high wall) and observed for 10 min. Locomotion frequencies were quantified by counting the number of intersquare lines crossed. An entry into a square was counted once the mouse had entered a new square with all four paws. Rearing frequencies, the number of times that the mice stood on their hind legs, were also recorded. The impairment of limb movement was assessed by a traction test as described by Wang et al. 5 Briefly, a mouse was hanged from a horizontal wire by its forepaws, and was scored according to how many hindpaws were used to grip the wire over a period of 1 min. Gripping the wire with both hindpaws, one hindpaw, and neither hindpaws was scored 3, 2, and 1, respectively.
Tissue preparation
After behavioral tests, mice were anesthetized with sodium pentobarbital (50 mg/kg, i.p.) and decapitated. The brains were immediately removed and split into halves. Half of the brain was paraffin-embedded, and 7μm-thick sections encompassing the entire midbrain were prepared for immunohistochemistry. The other half of the brain was stored at − 80°C for western blot analyses.
Cell culture and drug treatments
Human neuroblastoma SH-SY5Y cells were cultured at 37°C in Dulbecco minimum essential medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum (FBS) in a humidified 95% air, 5% CO2 incubator. The cells were transferred to low serum medium (1% FBS/DMEM) 2 h before drug treatments. MPP+ (Sigma), curcumin, and SP600125 (Sigma) were dissolved in DMSO. Curcumin and SP600125 were added to the medium 2 h before MPP+ treatment.
Cell viability assay
Cell viability was measured in 96-well plates by quantitative colorimetric assay with 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), which is an indicator of the mitochondrial activity of living cells. Briefly, at the indicated time after treatments, 10 μL of MTT (final concentration, 0.5 mg/mL) was added to the medium, which was incubated at 37°C for 3 h. The MTT solution was removed, and 100 μL of DMSO was added to each well to dissolve the colored formazan crystals for 15 min. The absorbance of each aliquot was measured at 490 nm using a Sunrise RC microplate reader (Tecan Group, Maennedorf, Switzerland). Cell viability was expressed as the ratio of the signal obtained from treated cultures and control cultures.
Hoechst 33258 staining
Cells were grown on cover slides in six-well plates. After drug treatments, cells were fixed with 4% paraformaldehyde and stained with Hoechst 33258 (Sigma-Aldrich) for 5 min. Nuclear morphology was examined using a fluorescence microscope (IX51, Olympus). Dead cells and apoptotic bodies were determined by condensed or fragmented nuclei.
Immunohistochemical analyses and stereology
Paraffin sections were dewaxed in xylene and rehydrated through graded alcohol solutions and then rinsed in 0.1 M Tris-HCl-buffered saline (TBS, pH 7.4) and placed in TBS containing 3% hydrogen peroxide (H2O2) for 10 min. After rinsing with TBS, sections were boiled in a TEG buffer for 5 min in a microwave oven. The sections were rinsed in TBS, treated with 5% bovine serum albumin (BSA) for 1 h, and then incubated overnight with rabbit anti-tyrosine hydroxylase (TH, 1:1000; Cell Signaling, 2792), rat anti-dopamine transporter (DAT, 1:1000; Chemicon, MAB369), or rabbit anti-glial fibrillary acidic protein (GFAP, 1:200; Sigma, G4546) at 4°C in a humidified chamber. Control sections were treated with identical solutions but without primary antibody. After rinsing several times with TBS, sections were incubated with biotinylated goat anti-rabbit or rat immunoglobulin G (IgG) for 30 min at 37°C, followed by amplification with streptavidin peroxidase for 30 min at 37°C. The sections were then rinsed in TBS and incubated with 0.025% 3,3-diaminobenzidine (DAB) plus 0.0033% H2O2 in TBS for 5 min. The stained sections were dehydrated through graded alcohol solutions, cleared in xylene, and covered with neutral balsam. All sections were examined, and images were taken with a light microscope equipped with a digital camera. The images were further processed with Adobe Photoshop software.
The numbers of TH-positive neurons and GFAP-positive astrocytes were subsequently counted by unbiased stereology, and the density of DAT-IR fibers was subsequently determined using a computerized image analysis system. Briefly, every seventh section spanning the SN and the striatum was examined for the number of immunoreactive neurons or astrocytes and the density of DAT-IR fibers, and six sections in total were counted per animal. The density of immunoreactive fibers was calculated by averaging three sections per animal. All sections were coded and examined blind. The results were represented as the rate of surviving dopaminergic neurons and reactive astrocytes in the SN.
Western blot analyses
The preparation of lysates and the western blots was performed as described previously. 23 Briefly, midbrain tissues or culture cells were homogenized at 1:5 (wt/vol) in an ice-cold lysis buffer. The resulting homogenate was centrifuged at 12,000 rpm for 30 min at 4°C. The supernatant was collected, and total protein levels were measured using a BCA protein assay kit (Pierce Biotechnology, Rockford, IL). Proteins (30 μg) were separated on 10% sodium dodecyl sulfate (SDS) polyacrylamide gels and transferred onto polyvinylidene difluoride (PVDF) membranes (Millipore, CA). The membranes were blocked with 5% skim milk in TBS containing 0.1% Tween-20 for 3 h and then incubated with a primary antibody overnight at 4°C. The primary antibodies used were rabbit anti-phospho-SEK1/MKK4 (1:1000, Ser257/Thr261; Cell Signaling, 9156), rabbit anti-SEK1/MKK4 (1:1000; Cell Signaling, 9152), rabbit anti-phospho-SAPK/JNK (1:1000, Thr183/Tyr185; Cell Signaling, 9251), rabbit anti-SAPK/JNK (1:1000, Cell Signaling; 9252), goat anti-phospho-c-Jun (1:1000, Ser 63/73; Santa Cruz, 16132), rabbit anti-c-Jun (1:1000; Cell Signaling, 9162), rabbit anti-caspase-3 (1:1000; PharMingen International, 67341), and mouse anti-glyceraldehyde 3-phosphate dehydrogenase (GAPDH, 1:12,000, KC-5G5; Kang Chen, CA, 0811). Bound secondary antibodies were visualized using an enhanced chemiluminescence (ECL) kit (Pierce Biotechnology) using ChemDoc XRS with Quantity One software (BioRad, Hercules, CA). Blots were repeated at least three times for every condition. After development, the band intensities were quantified using Image-pro Plus 6.0 analysis software.
Statistical analysis
All values are expressed as mean ± standard deviation (SD). Statistically significant differences were determined by the analysis of variance in SPSS statistical software (SPSS, Inc., Chicago, IL). Differences were considered significant at p < 0.05.
Results
Curcumin alleviates MPTP-induced mouse behavioral symptoms
MPTP administration induced obvious Parkinsonian symptoms in mice as reported previously. 24,25 To examine whether curcumin could alleviate MPTP-induced mouse behavioral changes, the locomotion, rearing frequencies, and traction scores were measured in our PD animal models. The protective effect of curcumin against MPTP infusion in the open-field test is shown in Fig. 1. The MPTP + vehicle group exhibited a significant decrease in locomotion frequencies and rearing frequencies, compared with the vehicle control group on days 5 and 12 (p < 0.01; Fig. 1A,B). On day 12, the MPTP + curcumin group mice showed a significant increase in locomotion frequencies and rearing frequencies compared with the MPTP + vehicle group (p < 0.01; Fig.1A, B). In the traction test, the vehicle control mice gripped the wire with all four paws, whereas on day 12, in the MPTP + vehicle–treated group, mice gripped the wire only by their forepaws. MPTP + curcumin group mice gripped the wire by one or two hindpaws, and the traction scores were increased significantly compared with the MPTP + vehicle group (p < 0.01; Fig. 1C). These results suggest that the Parkinsonian symptoms of MPTP-treated mice were improved by curcumin treatment.
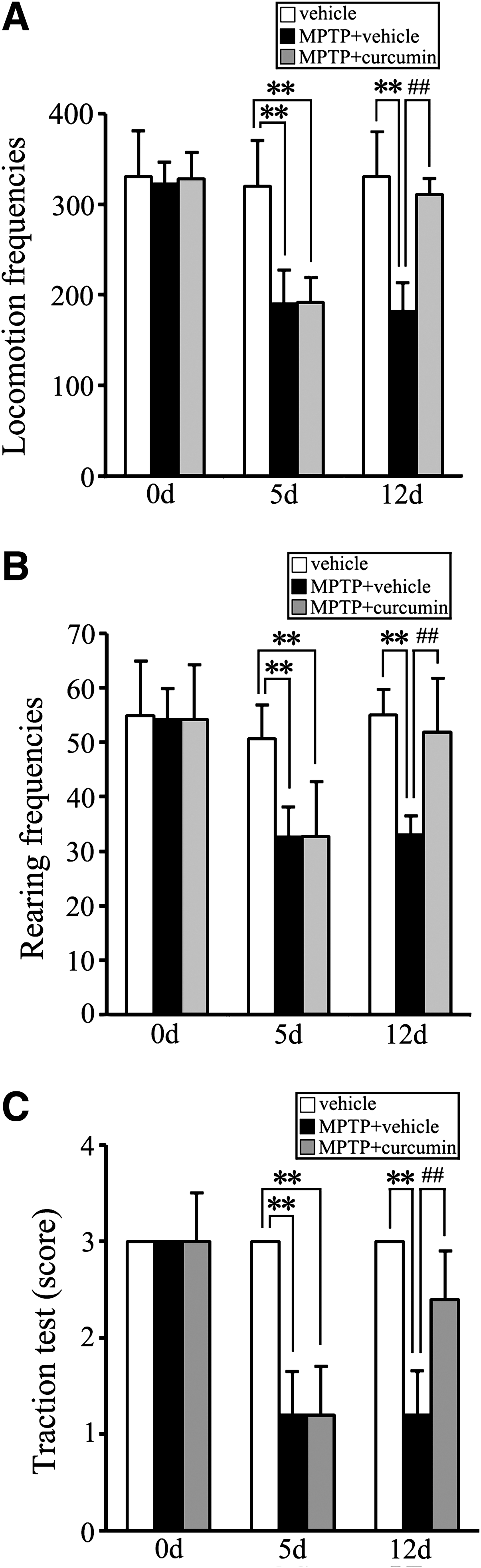
Curcumin improves 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine- (MPTP) induced Parkinsonian symptoms in mice. Open-field and traction analyses of mouse behavior, respectively, at day 0 (before administration), day 5 (injection of MPTP for 5 days), and day 12 (injection of vehicle or curcumin for 7 days). (
Curcumin protects against dopaminergic neuronal degeneration and inhibits astrocyte activation in mice
To explore the effects of curcumin on MPTP-induced loss of dopaminergic neurons, immunostaining was performed to detect the distribution of TH and DAT in the SN and striatum, respectively. As shown in Fig. 2, compared to the vehicle control mice, a significant loss of the dopaminergic neurons and nerve fibers was seen in the SN and striatum after MPTP + vehicle treatment. In contrast, those losses in the MPTP + curcumin group were clearly reduced (Fig. 2A,B). Unbiased stereological and optical density (O.D.) quantification showed that MPTP + vehicle produced a 43% loss of TH positive neurons in the SN and a 91% reduction of DAT-IR fibers, whereas curcumin administered once a day for 7 days improved the TH-positive neurons in the SN and DAT-IR fibers in the striatum to 82% and 36% of MPTP + curcumin-treated mice, respectively (p < 0.01 vs. MPTP + vehicle group; Fig. 2D,E). These results suggest that curcumin can protect against MPTP-induced impairment in the dopamine nervous system.
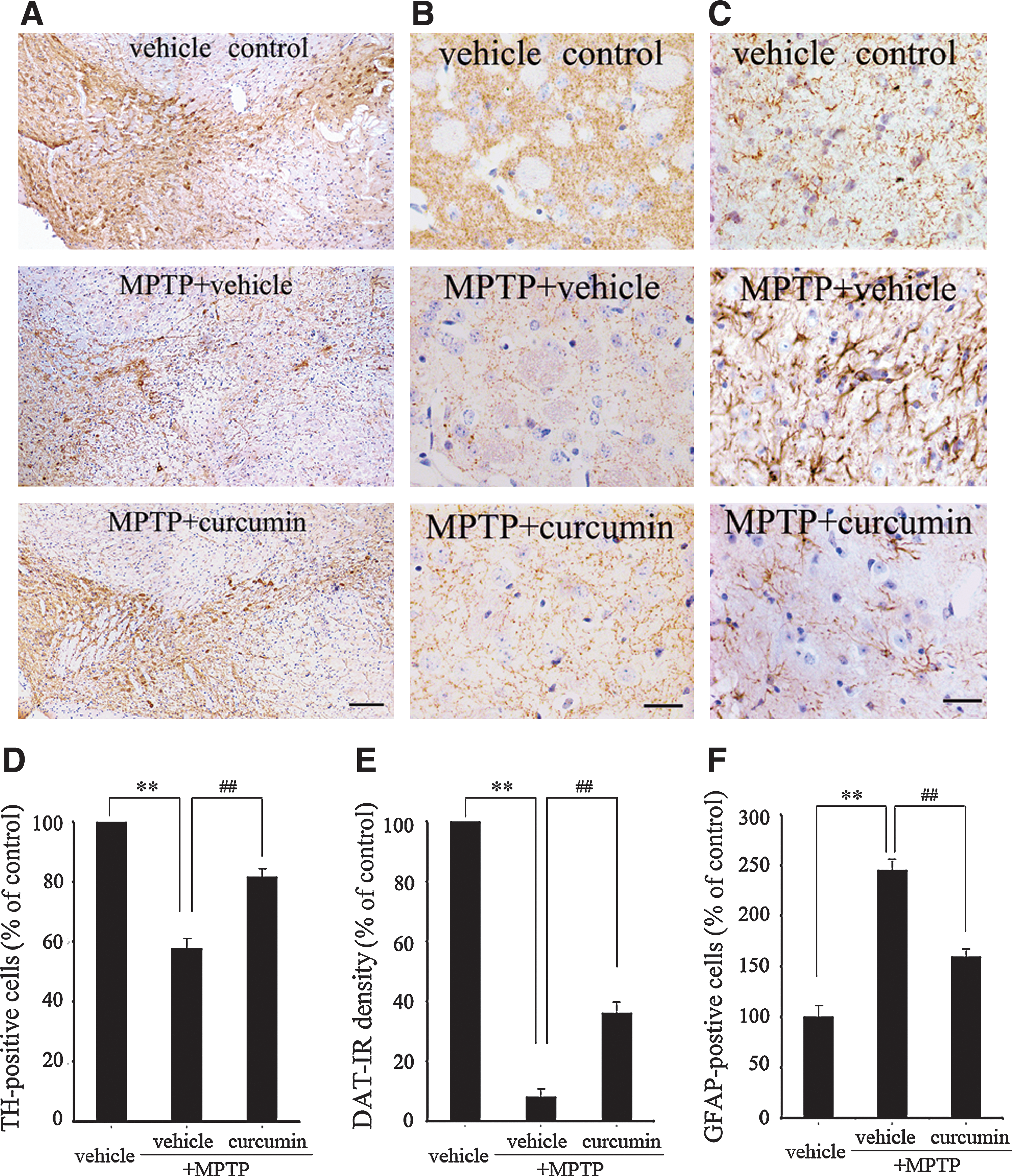
Curcumin protects against 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine- (MPTP) induced reduced expression of tyrosine hydroxylase (TH), dopamine transporter (DAT), and increased expression of GFAP in mouse substantia nigra and striatum. (
Astrocyte activity is mainly characterized by expression of GFAP, a major component of neurofilaments. GFAP overexpression is related to the astrocyte morphological changes in response to injury or inflammation. Thus, in the present study, we used the immunoreactivity of GFAP to represent the activity of astrocytes. Our results showed that MPTP + vehicle–treated mice exhibited 2.5 times more activated astrocytes than the vehicle control mice (p < 0.01 vs. vehicle control group; Fig. 2C,F). MPTP + curcumin treatment significantly attenuated the MPTP-induced increase in the number of activated astrocytes (p < 0.01 vs. MPTP + vehicle group; Fig. 2C,F).
Inhibitory effects of curcumin on MPTP-induced activation of JNK, c-Jun, and caspase-3 in mouse SN
Recent studies have demonstrated that JNK is an important transducing enzyme, and alterations in JNK pathways are potentially associated with dopaminergic neuronal death in PD. 26 –28 Therefore, with our PD animal models, we detected the expression levels of JNK and its downstream molecules, c-Jun and caspase-3, to determine if curcumin is able to prevent dopaminergic neuronal death through inhibition of JNK signalling cascades. Our immunoblotting results showed that MPTP induced a robust increase in phosphorylated forms of both JNK1 and JNK2, the key molecules in the JNK pathways, in the SN (p < 0.01; Fig. 3A). Interestingly, curcumin treatment (50 mg/kg i.p.q.d. for 7 days) significantly inhibited MPTP-induced JNK1/2 phosphorylation (Fig. 3A), without alteration of the total protein levels of JNK1/2 (Fig. 3A).
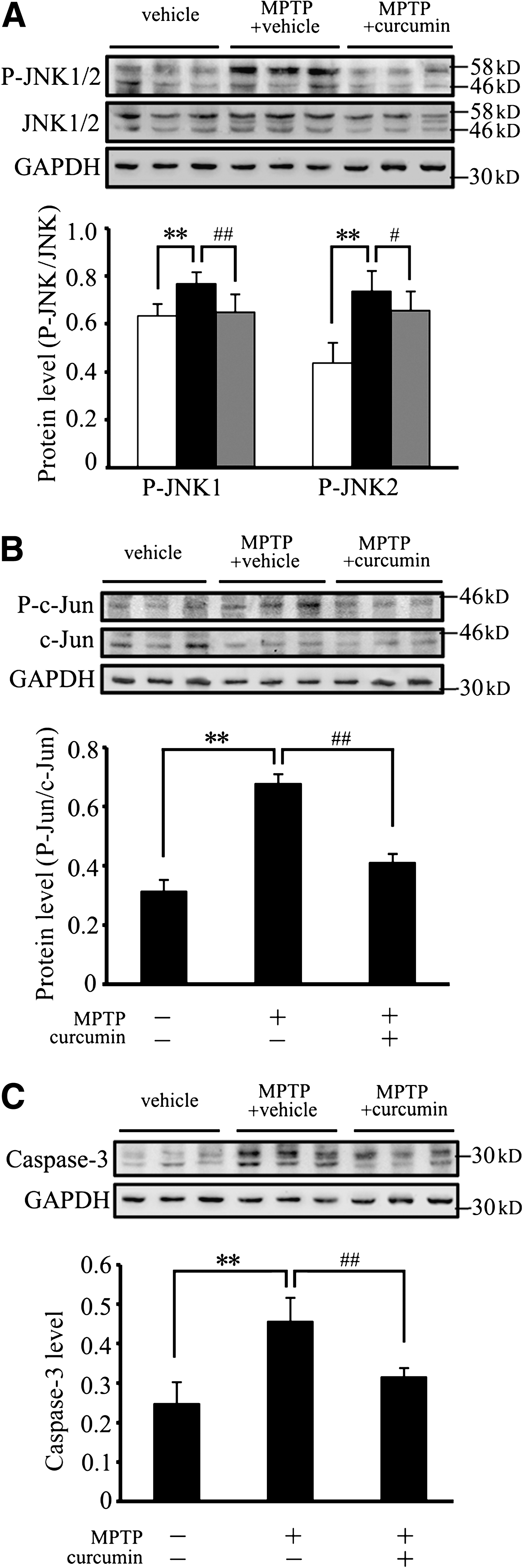
Curcumin reduces 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine- (MPTP) induced c-Jun N-terminal kinase (JNK) and c-Jun phosphorylation, and caspase-3 activation in mouse substantia nigra (SN). The SN from vehicle control, MPTP + vehicle, or MPTP + curcumin-treated mice was lysed and subjected to immunoblotting analysis using SAPK/JNK, P-SAPK/JNK, c-Jun, P-c-Jun, and caspase-3 antibodies. An equal amount of total proteins was loaded into each lane. Quantification showed that P-JNK1/2 (
c-Jun is an important downstream target of JNK, and its activation has been implicated in a wide range of cellular events, including caspase-dependent apoptosis. 29 As shown in Fig. 3, the expression levels of c-Jun and cleaved caspase-3 were increased in the SN of MPTP + vehicle–treated mice (p < 0.01; Fig. 3B,C), whereas curcumin administration inhibited the activation of c-Jun and cleaved caspase-3, which was induced by MPTP (p < 0.01; Fig. 3B,C). Taken together, these results suggest that curcumin markedly inhibits MPTP-induced activation of the JNK pathway, which may result in the reduction of dopaminergic neuronal degeneration.
Curcumin attenuates MPP+-induced SH-SY5Y cell death
To further investigate the inhibitory effect of curcumin on the activation of the JNK pathway and dopaminergic neuronal degeneration, MPP+-mediated human neuroblastoma SH-SY5Y cell damage was used as a model for PD for in vitro study. First, we examined whether the curcumin-dependent neuroprotection in vivo could be extended to MPP+-induced SH-SY5Y cell death using the MTT assay. As shown in Fig. 4, the concentrations of MPP+, curcumin, and SP600125, a well-known JNK inhibitor, for SH-SY5Y cell viability were determined by MTT assay. SH-SY5Y cells treated with MPP+ (0–5 mM) showed a dose- and time-dependent reduction in the levels of cell viability (Fig. 4A). A 50% reduction in cell viability was obtained when cells were treated with 3 mM MPP+ for 18 h. The MTT assay also showed that curcumin (1–5 μM) and SP600125 (5–10 μM) did not exhibit significant toxic effects on the cells (Fig. 4B). In the subsequent experiments, the addition of 3 mM MPP+ for 18 h was used to maintain a 50% reduction in cell viability. When the cells were pretreated with 1 μM curcumin and 5 μM SP600125 for 2 h prior to the addition of MPP+, the cell viability was significantly increased (Fig. 4C).
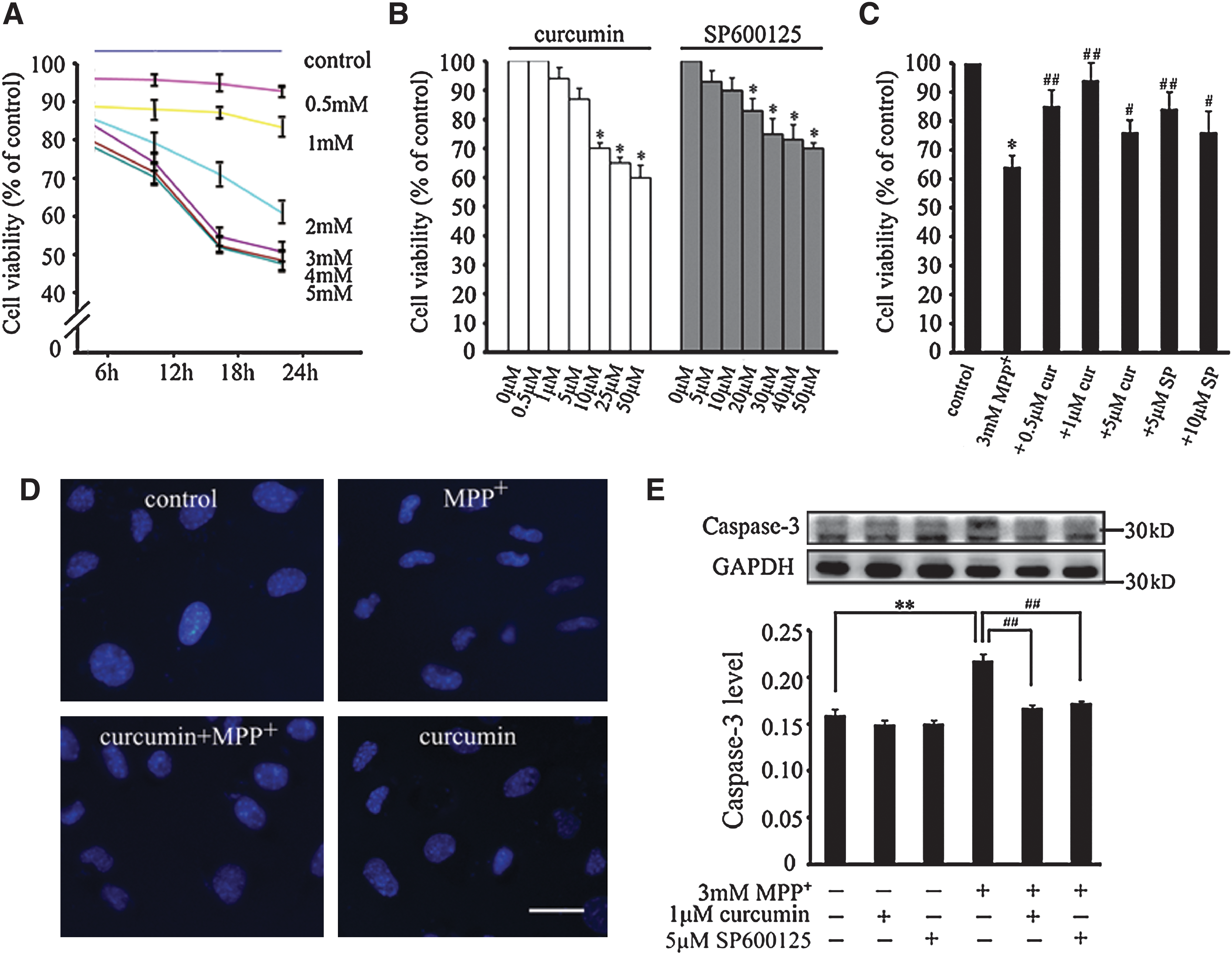
Curcumin attenuates 1-methyl-4-phenylpyridnium ion- (MPP+) induced SH-SY5Y cell apoptosis. (
The protective effect of curcumin on MPP+-induced apoptosis was further verified by Hoechst 33258 staining and caspase-3 immunoblotting. SH-SY5Y cells were pretreated with 1 μM curcumin or 5 μM SP600125 for 2 h, and exposed to 3 mM MPP+ for 18 h. As seen in Fig. 4D, Hoechst 33258 staining showed that distinct nuclear condensation was observed in MPP+-treated cells. This phenomenon was inhibited when curcumin was added prior to MPP+ treatment. It is known that MPP+-induced dopaminergic neuronal cell apoptosis is tightly regulated by caspase-3. 30 Therefore, we performed immunoblotting to analyze the changes in the expression levels of caspase-3 protein in our cell model. As shown in Fig. 4E, MPP+ treatment significantly increased the cleaved caspase-3 protein levels (p < 0.01 vs. control), whereas the pretreatment with curcumin and SP600125 blocked the upregulation of cleaved caspase-3 produced by MPP+ treatment (p < 0.01 vs. MPP+-treated group). Collectively, these results suggest that curcumin serves as a protective factor in MPP+-induced cytotoxic process in SH-SY5Y cells.
Curcumin inhibits JNK pathway activation in SH-SY5Y cells
Previous studies have demonstrated that JNK is involved in protecting neurons against MPP+-triggered dopaminergic neurotoxicity. Because curcumin suppressed the MPTP-induced JNK activation in our in vivo experiments, we also investigated whether the JNK pathway is the direct target of curcumin in our SH-SY5Y cell model. Meanwhile, SP600125, a specific inhibitor of JNK, was employed as a positive control of curcumin under the same conditions. Our immunoblotting results showed that the phosphorylated forms of both JNK1 and JNK2 were increased significantly in response to MPP+ stimuli (p < 0.01 vs. control; Fig. 5A). The expression levels of JNK1 and JNK2 phosphorylation were inhibited by pretreatment with 1 μM curcumin, and 5 μM SP600125 (p < 0.01 vs. MPP+-treated group; Fig. 5A). The changes in the expression levels of MKK4 phosphorylation, an upstream molecule of JNK protein, were similar to that of JNK protein, i.e., curcumin could also inhibit MPP+-induced MKK4 activation in SH-SY5Y cells (p < 0.01 vs. MPP+-treated group; Fig. 5B).
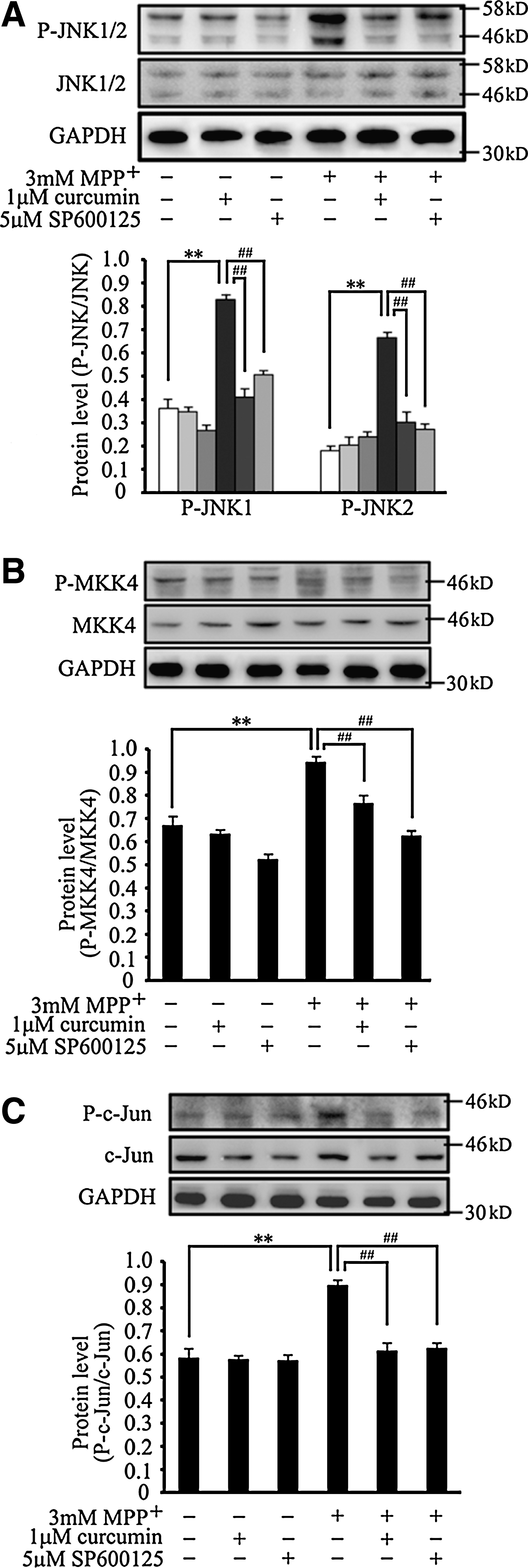
Curcumin inhibits c-Jun N-terminal kinase (JNK), MKK4, and c-Jun phosphorylation induced by 1-methyl-4-phenylpyridnium ion (MPP+) in SH-SY5Y cells. SH-SY5Y cells were pretreated with curcumin or SP600125 for 2 h and then exposed to 3 mM MPP+ for 18 h. Cell lysates were subjected to immunoblotting using SAPK/JNK, P-SAPK/JNK, SEK1/MKK4, P-SEK1/MKK4, c-Jun, P-c-Jun, and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) antibodies. An equal amount of total protein was loaded into each lane. Quantification showed changes in the levels of P-JNK1/2 (
We then examined the inhibitory effect of curcumin on JNK downstream events by detecting the expression levels of c-Jun phosphorylation. Coincident with the result of JNK phosphorylation, MPP+-induced c-Jun phosphorylation was inhibited by curcumin in SH-SY5Y cells (p < 0.01 vs. MPP+-treated group; Fig. 5C), which is similar to the effect of SP600125 on c-Jun activation. These results indicated that curcumin could protect against MPP+-induced cell death through inhibiting the activation of the JNK pathway in SH-SY5Y cells.
Discussion
Several epidemiological studies indicate that there is a reduced prevalence of AD and PD in India, where about 60–200 mg/day of dietary curcumin is consumed. 31 The novelty of using curcumin as a neuroprotective component is its relative lack of toxicity and lack of major side effects. 32,33 Although a number of studies have shown that the mechanisms underlying the neuroprotective effects of curcumin are multifactorial, such as antiinflammatory and antioxidant actions, our results lend support to the view that curcumin protects against MPTP/MPP+-induced dopaminergic cell death through inhibition of JNK phosphorylation. These results suggest that curcumin is a JNK inhibitor, and curcumin treatment is a potential strategy for the prevention and treatment of PD.
Several studies have shown that administration of MPTP in mammals can replicate the selective degeneration of dopaminergic neurons in the SN, 24,25 thereby mimicking the behavioral and pathological features of PD. Also, MPP+ can induce dopaminergic neuronal death in several culture cell lines. 34 –36 In the present study, we found that curcumin could increase in locomotion frequencies, rearing frequencies, and limb movement compared with the MPTP-treated mice. Curcumin could also inhibit the MPTP/MPP+-induced reduction in dopaminergic neurons, as revealed by detection of the immunostaining of TH and DAT in mouse SN and striatum, MTT assay, and Hoechst 33258 staining in cultured SH-SY5Y cells. Furthermore, our immunoblotting assay showed that curcumin could inhibit the activation of caspase-3 both in vivo and in vitro. Collectively, these data indicate that curcumin can directly protect against the toxic effects of MPTP/MPP+ on dopaminergic neurons.
JNK is a member of the mitogen-activated protein kinase (MAPK) superfamily and is an important kinase mediating neuronal cell death induced by cell stress-inducing signals. 26,27,29 In the nigrostriatal pathway, the JNK signaling pathway is activated by MPTP through increasing the levels of JNK phosphorylation, and this increased phosphorylation is inhibited by the indolocarbazole CEP-1347/KT-7515, an inhibitor of the JNK pathway. 37 CEP-1347/KT-7515 also attenuates MPTP-mediated nigrostriatal dopaminergic cell loss in vivo. 38 More recently, a selective JNK inhibitor, SP600125, was found to protect dopaminergic neurons from MPTP/MPP+-induced neurotoxicity. 4,5 Consistent with the evidence from the MPTP-induced PD model, nigrostriatal dopaminergic neuronal death is also associated with activation of the JNK signaling pathway in the 6-OHDA lesion model of PD. 6 Therefore, it is reasonable to speculate that blockade of JNK by specific inhibitors may prevent or effectively slow the progression of PD. To further clarify whether the neuroprotective effects of curcumin on MPTP/MPP + -induced dopaminergic neuronal loss are through inhibition of the JNK pathway, in the present study, the changes in the expression levels of key molecules in the JNK pathway have been examined in our in vivo and in vitro PD models.
Our results showed that JNK1/2 phosphorylation was increased in response to the selective nigrostriatal dopaminergic neurotoxin MPTP/MPP+ treatments. However, curcumin treatment significantly inhibited MPTP/MPP+-induced JNK activation in dopaminergic neurons in mouse SN and in human neuroblastoma SH-SY5Y cells, respectively. It has been reported that the MPTP-mediated activation of JNK is accompanied by an increase in MKK4 phosphorylation, the upstream signaling of the JNK protein. 39 Our results showed that MKK4 was upregulated in MPP+-treated cells, and this increased phosphorylation was inhibited by pretreatment with curcumin. We also found that the MPP+-induced phosphorylation of c-Jun, the downstream target of JNK, was inhibited by pretreatment of curcumin in SH-SY5Y cells. It is worth noting that parallel experiments involving pretreatment with curcumin and SP600125, a specific JNK inhibitor, 40 were performed in our PD cell model, and the inhibitory effects of both curcumin and SP600125 on JNK were always quite similar. Taken together, the present data indicate that curcumin is an inhibitor of the JNK pathway and, hence, it protects dopaminergic neurons from death induced by MPTP/MPP+.
In addition, our results showed that curcumin could inhibit the MPTP-induced increase in GFAP-positive astrocytes in mouse SN. It has been reported that curcumin is an antiinflammatory and antioxidant agent . 41 Several lines of evidence have indicated that inflammation is a mechanism involved in dopaminergic neuronal degeneration, 42 and astrocytes have been suspected of being instrumental in this process. 43 –45 Our findings that curcumin reduced the MPTP-mediated increase in astrocytes indicate that curcumin may inhibit the activation of astrocytes and the release of neurotoxic molecules from reactive astrocytes, thereby protecting dopaminergic neurons after MPTP insult. Further studies are needed to validate this notion.
In summary, the present study indicates that curcumin has neuroprotective effects against MPTP/MPP+-induced dopaminergic neuronal death by a mechanism believed to inhibit the JNK pathway, which suppresses the increase in caspase-3 in dopaminergic neurons. Our data provide experimental evidence to explore the daily consumption and pharmaceutical use of curcumin, a relatively nontoxic and convenient natural compound, as an efficient approach to the prevention and treatment of PD.
Footnotes
Acknowledgments
The study was supported by the Natural Science Foundation of China (30770680), the Program for New Century Excellent Talents in University (NCET-04-0288), the China Postdoctoral Science Foundation (2005037008), and the Specialized Research Fund for the Doctoral Program of Higher Education (SRFDP-20060159001).
Author Disclosure Statement
No competing financial interests exist.