Previously, two d-enantiomeric amino acid peptides, D1 and D3, which specifically bind to the amyloid-β peptide Aβ(1–42), were identified by phage display selection. To assess the diagnostic and therapeutic potentials of D1 and D3 for the diagnosis and treatment of Alzheimer disease, the blood–brain barrier transport of these d-peptides was quantitatively evaluated in vitro. Our results showed that the apical-to-basolateral transport of D3 was more efficient than that of D1. An active efflux transport mechanism seems to oppose the transport of D1, whereas D3 is likely to be transported through the blood–brain barrier via an adsorptive-mediated transcytosis mechanism.
Introduction
Alzheimer disease (AD) is an age-related progressive neurodegenerative disorder of the brain, which is characterized by cognitive deterioration as well as behavioral, affective, and psychotic disturbances. Pathologically it is characterized by the presence of senile plaques and neurofibrillary tangles in the brain. The senile plaques mainly consist of amyloid-β (Aβ) peptides derived from cleavage of the amyloid precursor protein (APP), with the majority in plaques being Aβ(1–42), which is particularly prone to aggregation. Several lines of evidence suggest a causative role of Aβ species in AD1–3 and thus support it as a promising target for AD drug development and treatment.
Previously, two d-amino acid peptides, D1 (QSHYRHISPAQV) and D3 (RPRTRLHTHRNR), which specifically bind to Aβ(1–42), were identified by phage display selection using the d-enantiomer of Aβ(1–42) as the target.4,5In vitro and in vivo studies showed that D1 was a promising candidate for the diagnosis of AD.4,6,7 D3 was found to modulate Aβ aggregation, reduce Aβ cytotoxicity in cell culture, and drastically reduce the Aβ plaque load and cerebral inflammation in transgenic AD mice,5,6 thus showing a promising therapeutic potential for the treatment of AD. Moreover, as D1 and D3 are composed exclusively of d-amino acid residues, they are thought to be protease resistant and less immunogenic than the respective L-enantiomers, thus being principally suited for in vivo use. However, one major drawback with the use of peptides as drugs in neurological diseases is their expected poor blood–brain barrier (BBB) permeability. The objective of the present study is to examine whether these two d-peptides are able to cross the BBB by using an in vitro BBB model, and if so, to investigate the underlying mechanisms.
Materials and Methods
Peptide lipophilicity and basicity
The relative lipophilicity of each d-peptide was determined by 1-octanol/buffer partition coefficient, as described previously.8 The basicity of d-peptides was represented by the theoretical isoelectric point, which was estimated from the acid–base dissociation constants of ionizable functional groups in each d-peptide.
In vitro BBB model
The BBB model was a co-culture of rat brain microvascular endothelial cells (RBMECs) and rat astrocytes prepared on a collagen/fibronectin-coated polyester Transwell filter (pore size, 0.4 μm; surface area, 1.13 cm2), as described previously.9,10 Chambers containing the RBMECs and the astrocytes are considered as the apical (blood) and the basolateral (brain) compartments, respectively. The barrier formation was examined by measuring the permeability of sodium fluorescein (FLU), a paracellular marker, and propranolol, a transcellular marker. The permeability coefficient (Pe, in cm/min) was calculated as described elsewhere, 11 using filters either coated with endothelial cells or uncoated.
Transport study
Permeability measurements for each compound were run for 150 min at least three times. After preincubation of 30 min in Ringer-HEPES buffer at 37°C, the d-peptide was introduced into the donor chamber (either the apical or the basolateral compartment). At various time points after addition of the d-peptide, the assay buffer was removed from the opposite (receiver) chamber of the well for concentration determination and was immediately replaced by fresh assay buffer to minimize the back diffusion of the d-peptide. Concentration of the d-peptide was analyzed by reversed-phase high-performance liquid chromatography (RP-HPLC). The influence of temperature on the transport of d-peptides was measured with the co-culture system kept at 4°C. The effects of several compounds on the transport of D1 and D3 were determined in the apical-to-basolateral direction.
Results
D1 and D3 showed quite different physicochemical characteristics. D1 (D = 0.0149 ± 0.0012) was found to be approximately 8 times more lipophilic than D3 (D = 0.0019 ± 0.0004). The isoelectric points of D1 and D3 were calculated to be 9.3 and 13.1, respectively.
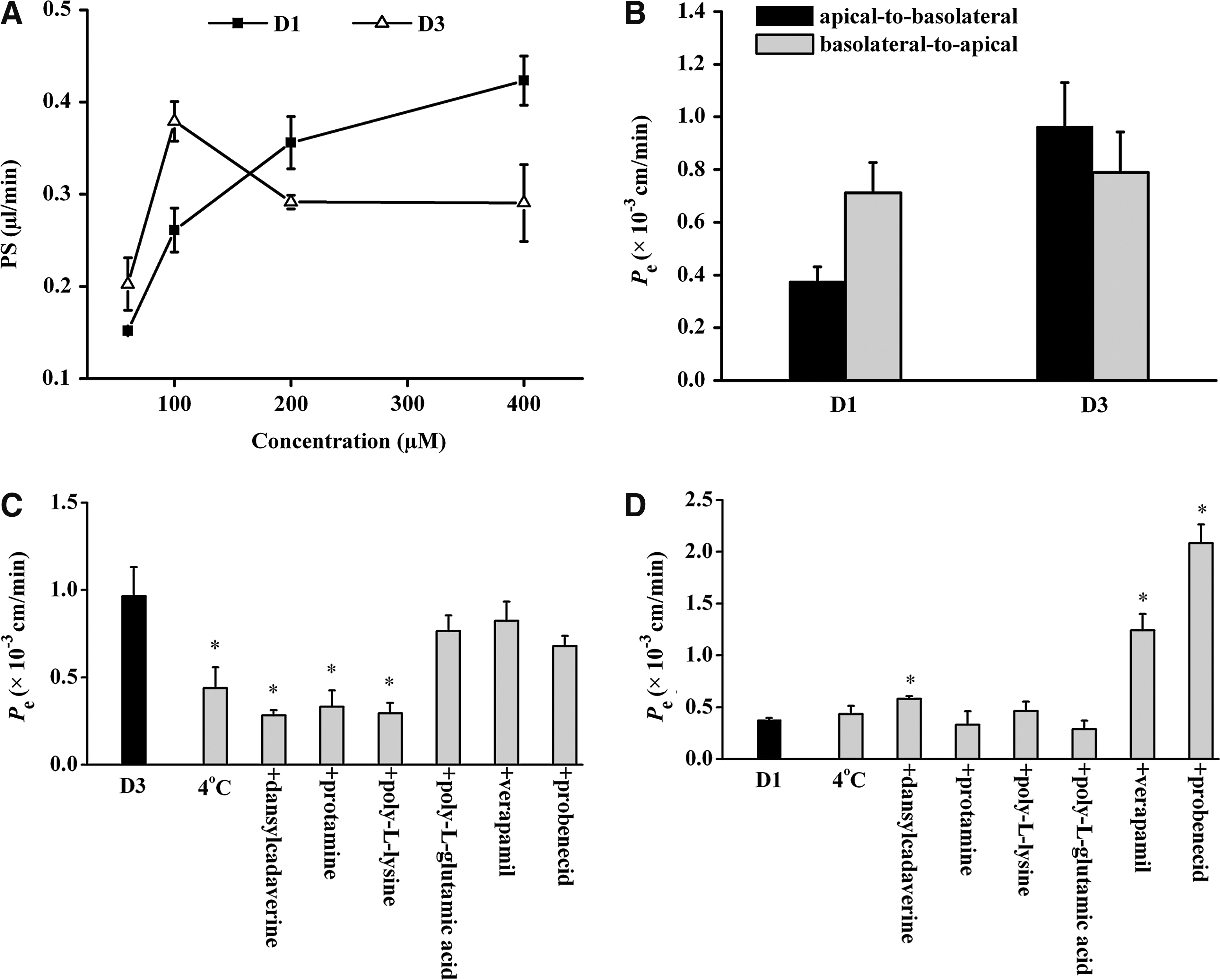
Before the in vitro BBB model was used to measure the BBB passage of d-peptides, the integrity of the barrier was estimated by assessing the permeability of FLU. The permeability coefficient of FLU was very low (0.31 ± 0.03 × 10−3 cm/min), demonstrating that the co-culture system was well intact. In contrast, propranolol, which is known to easily penetrate the BBB, showed a high permeability in the in vitro BBB model (Pe = 1.8 ± 0.2 × 10−3 cm/min). These results were consistent with those of previous reports.10,12 D1 and D3 at varying concentrations were added to the apical compartment of the co-culture system. The results were presented by the perm-eability–surface area product (μL/min) (Fig. 1A). The transport of D1 was found to be concentration-dependent in the assayed concentration range. However, the convex manner of the increase implied the presence of saturation. D3 seemed to achieve a plateau at 200 μM.
(A) Influence of concentration on the apical-to-basolateral transport of D1 and D3 across the in vitro blood–brain barrier (BBB) model. The results were presented by the permeability–surface area product (PS, μL/min). Each point is the mean of three different filters. (B) Directional transport of D1 and D3 at 100 μM across the in vitro BBB model at 37°C. The permeability coefficient (Pe) was calculated from the slopes of the curves for cleared volume of d-peptides versus time. Values represent the mean ± standard deviation (SD) of four measurements. (C and D) Effects of low temperature (4°C), an endocytosis inhibitor dansylcadaverine (0.5 mM), cationic substances protamine (0.5 mg/mL), and poly-l-lysine (0.1 mg/ml), an anionic peptide poly-l-glutamic acid (0.1 mg/mL), a P-glycoprotein inhibitor verapamil (100 μM), and a multidrug resistance-associated protein inhibitor probenecid (100 μM) on the transport of D3 (C) and D1 (D) at 100 μM across the in vitro BBB model in the apical-to-basolateral direction. Statistical analyses were performed using the Student t-test or one-way analysis of variance followed by the Dunnett test. Significance was assumed for p values less than 0.05.
Next, we investigated the directional transport of D1 and D3 at 100 μM. As seen in Fig. 1B, there was a significant directionality in the transport of D1 across the in vitro BBB, with the basolateral-to-apical transport 1.9-fold higher than the apical-to-basolateral transport. In contrast, D3 showed a slightly higher apical-to-basolateral transport than the basolateral-to-apical transport. These results indicated that the apical-to-basolateral transport of D3 was more efficient than that of D1 at 100 μM.
To elucidate the possible mechanisms underlying the transport of d-peptides across the BBB, we examined their transport under various conditions. As shown in Fig. 1C, the apical-to-basolateral transport of D3 was inhibited by 55% at 4°C, but by up to approximately 70% with the addition of dansylcadaverine (an endocytosis inhibitor) and cationic substances (poly-l-lysine and protamine). On the other hand, poly-l-glutamic acid (an anionic peptide), verapamil (a P-glycoprotein inhibitor), and probenecid (a multidrug resistance-associated protein [MRP] inhibitor) had no significant effect. In contrast, the transport of D1 at 4°C was very similar to the transport value obtained at 37°C. No significant changes in D1 permeability were noted in the presence of protamine, poly-l-lysine, or poly-l-glutamic acid. However, pretreatment of the BBB model with dansylcadaverine, verapamil, and probenecid significantly increased the apical-to-basolateral transport of D1 (Fig. 1D).
To validate the data from the experiments with P-glycoprotein inhibition by verapamil, the transport of the P-glycoprotein substrate verapamil in both directions was tested. A 3.5-fold higher basolateral-to-apical permeability of verapamil (7.58 ± 0.69 × 10−3 vs. 2.14 ± 0.49 × 10−3 cm/min) indicated the presence of an apical efflux system.13,14 In a separate experiment, the transport of FLU measured in the presence of verapamil (Pe = 0.36 ± 0.05 × 10−3 cm/min) was similar to that measured for the control, indicating that verapamil did not affect BBB integrity.
Discussion
In the present study, we characterized the transport properties of D1 and D3 across the BBB using an in vitro BBB co-culture system. Using various conditions, the underlying transport mechanisms were studied.
This co-culture model allows the assessment of the BBB permeability of drugs in both directions (influx and efflux) independently. No indication of directional transport of D3 was obtained in the present in vitro assay. The lack of asymmetric transport could be explained by equal active transport in both directions. Our data showed that the apical-to-basolateral transport of D3 through the BBB was partially saturable, suggesting that a more or less specific transport system for D3 might exist. One feature of D3 is the high number of basic amino acids—it contains five arginine residues out of 12 amino acid residues. The high number of positive charges at physiological pH would make passive diffusion through the BBB very unlikely for D3. Adsorptive-mediated transcytosis (AMT) has been proposed for the transport of several arginine-rich peptides, such as basic peptides derived from Antennapedia and the human immunodeficiency virus type 1 (HIV-1) Tat proteins, across the BBB.15,16 In addition, the basicity of peptides is supposed to be an important factor for the transport via the AMT mechanism.17 Therefore, we next investigated the possibility that D3 might be transported via AMT. The apical-to-basolateral transport of D3 showed temperature dependency and was significantly inhibited by dansylcadaverine and polycations, confirming the functional involvement of AMT. In addition, our results suggested that D3 did not seem to exhibit any interaction with P-glycoprotein or MRPs at the BBB. However, other efflux and influx transporters remain to be tested.
Compared to D3, D1 showed lower BBB permeability than predicted from its lipophilicity. This may be due to active transport of D1 by apical efflux transport systems in the BBB. Previously, it was reported that astrocytes increased the functional expression of P-glycoprotein on endothelial cells using a similar co-culture system.13 Herein, our results indicated that the mechanism by which verapamil increased the transport of D1 across the BBB was related to the inhibition of P-glycoprotein–mediated efflux. In addition, probenecid-sensitive efflux transporters18,19 (e.g., MRPs) were also involved in D1 transport. However, it is interesting to note that dansylcadaverine slightly but significantly increased D1 transport. The underlying mechanism is not clear yet.
In conclusion, our study showed that the two d-peptides under study differed in their degree of BBB passage and transport mechanism. An active efflux transport mechanism seems to oppose the transport of D1 across the BBB, whereas D3 is likely to be transported through the BBB via an AMT mechanism.
Footnotes
Acknowledgments
The authors gratefully acknowledge a support from the “Helmholtz–CSC (China Scholarship Council) Junior Scientists Exchange Program.” We also thank Esther Jonas for excellent technical assistance.
Author Disclosure Statement
The authors have no competing financial interests.
WalshDM, KlyubinI, FadeevaJV, CullenWK, AnwylR, WolfeMS, RowanMJ, SelkoeDJ. Naturally secreted oligomers of amyloid beta protein potently inhibit hippocampal long-term potentiation in vivo. Nature, 2002; 416:535–539.
3.
LaFerlaFM, OddoS. Alzheimer's disease: Abeta, tau and synaptic dysfunction. Trends Mol Med, 2005; 11:170–176.
4.
WiesehanK, BuderK, LinkeRP, PattS, StoldtM, UngerE, SchmittB, BucciE, WillboldD. Selection of d-amino-acid peptides that bind to Alzheimer's disease amyloid peptide abeta1-42 by mirror image phage display. Chembiochem, 2003; 4:748–753.
5.
van GroenT, WiesehanK, FunkeSA, KadishI, Nagel-StegerL, WillboldD. Reduction of Alzheimer's disease amyloid plaque load in transgenic mice by D3, A d-enantiomeric peptide identified by mirror image phage display. ChemMedChem, 2008; 3:1848–1852.
6.
van GroenT, KadishI, WiesehanK, FunkeSA, WillboldD. In vitro and in vivo staining characteristics of small, fluorescent, Abeta42-binding d-enantiomeric peptides in transgenic AD mouse models. ChemMedChem, 2009; 4:276–282.
7.
WiesehanK, StöhrJ, Nagel-StegerL, van GroenT, ReisnerD, WillboldD. Inhibition of cytotoxicity and amyloid fibril formation by a d-amino acid peptide that specifically binds to Alzheimer's disease amyloid peptide. Protein Eng Des Sel, 2008; 21:241–246.
8.
LiuHM, LiuXF, YaoJL, WangCL, YuY, WangR. Utilization of combined chemical modifications to enhance the blood-brain barrier permeability and pharmacological activity of endomorphin-1. J Pharmacol Exp Ther, 2006; 319:308–316.
9.
DehouckMP, Jolliet-RiantP, BreeF, FruchartJC, CecchelliR, TillementJP. Drug transfer across the blood-brain barrier: correlation between in vitro and in vivo models. J Neurochem, 1992; 58:1790–1797.
10.
GaillardPJ, VoorwindenLH, NielsenJL, IvanovA, AtsumiR, EngmanH, RingbomC, de BoerAG, BreimerDD. Establishment and functional characterization of an in vitro model of the blood-brain barrier, comprising a co-culture of brain capillary endothelial cells and astrocytes. Eur J Pharm Sci, 2001; 12:215–222.
11.
PardridgeWM, TrigueroD, YangJ, CancillaPA. Comparison of in vitro and in vivo models of drug transcytosis through the blood-brain barrier. J Pharmacol Exp Ther, 1990; 253:884–891.
12.
BararJ, OmidiY. Bioelectrical and permeability properties of brain microvascular endothelial cells: Effects of tight junction modulators. J Biol Sci, 2008; 8:556–562.
13.
GaillardPJ, van der SandtIC, VoorwindenLH, VuD, NielsenJL, de BoerAG, BreimerDD. Astrocytes increase the functional expression of P-glycoprotein in an in vitro model of the blood-brain barrier. Pharm Res, 2000; 17:1198–1205.
14.
MegardI, GarriguesA, OrlowskiS, JorajuriaS, ClayetteP, EzanE, MabondzoA. A co-culture-based model of human blood-brain barrier: Application to active transport of indinavir and in vivo-in vitro correlation. Brain Res, 2002; 927:153–167.
15.
DrinG, CottinS, BlancE, ReesAR, TemsamaniJ. Studies on the internalization mechanism of cationic cell-penetrating peptides. J Biol Chem, 2003; 278:31192–31201.
16.
TanabeS, ShimohigashiY, NakayamaY, MakinoT, FujitaT, NoseT, TsujimotoG, YokokuraT, NaitoM, TsuruoT, TerasakiT. In vivo and in vitro evidence of blood-brain barrier transport of a novel cationic arginine-vasopressin fragment 4–9 analog. J Pharmacol Exp Ther, 1999; 290:561–568.
17.
TamaiI, SaiY, KobayashiH, KamataM, WakamiyaT, TsujiA. Structure-internalization relationship for adsorptive-mediated endocytosis of basic peptides at the blood–brain barrier. J Pharmacol Exp Ther, 1997; 280:410–415.
18.
SugiyamaD, KusuharaH, ShitaraY, AbeT, MeierPJ, SekineT, EndouH, SuzukiH, SugiyamaY. Characterization of the efflux transport of 17beta-estradiol-d-17beta-glucuronide from the brain across the blood-brain barrier. J Pharmacol Exp Ther, 2001; 298:316–322.
19.
ZelcerN, HuismanMT, ReidG, WielingaP, BreedveldP, KuilA, KnipscheerP, SchellensJH, SchinkelAH, BorstP. Evidence for two interacting ligand binding sites in human multidrug resistance protein 2 (ATP binding cassette C2)J Biol Chem, 2003; 278:23538–23544.