
Abstract
The increase in free radicals is hypothesised to cause aging and age-related diseases. The most common source of free radicals is thought to be superoxide. This superoxide is claimed to be released from mitochondria during the enzymatic transformation of oxygen to water by a “leakage” process. This article presents evidence that leakage does not occur. Instead, protonated superoxide radicals are generated by lipid peroxidation processes. In nature, polyunsaturated fatty acids (PUFAs) represent particularly oxygen-sensitive compounds. Apparently, nature uses this sensitivity for signalling processes by producing lipidhydroperoxides (LOOHs) by any change to cell membrane structure. LOOHs easily undergo further enzymatic transformations to compounds which contribute to activation of genes. Bivalent metal ions within the active site of lipoxygenases catalyse LOOH production. The metal ions generate radicals which are transformed within the enzyme complex to non-radical molecules. Thus radicals never leave the enzyme complex except in severe stress situations. In this case the radical intermediates attack bonds, keeping the metal ion in its complex state. Thus, metal ions are released and react in a Fenton reaction with LOOH molecules produced earlier by lipoxygenase to form LO· radicals. Radicals are typically four orders of magnitude more reactive than non-radical molecules. Their action is not under genetic control, they attack nearly all biological molecules, destroying lipids, proteins, nucleic acids, hormones and enzymes until the radicals are quenched by scavenger molecules. The principal degradation routes are outlined in this review. Lysophospholipids are generated in large amounts after stress by activation of phospholipases and are transformed to LO· radicals. These can abstract a hydrogen radical from a lysophospholipid. The radical thus formed adds oxygen and decomposes to a 2-oxolysophospholipid and a HOO· radical (protonated superoxide). HOO· radicals in turn abstract hydrogen atoms from other molecules and produce H2O2. Since any cell preparation method causes membrane destruction, it is inevitable that protonated superoxide is generated, explaining why H2O2 molecules are found as “byproducts” of many reactions.
Theories of Aging
The Hayflick theory postulates that aging and death are programmed in cells and is therefore under genetic control. In 1892, Weismann concluded that the number of cell divisions should be limited. 22 His thesis was confirmed by Hayflick and Moorhead about 70 years later, who detected that cells cannot undergo more than about 50 divisions, 23 confirmed in succeeding experiments for different fibroblast cell lines. 24 Later experiments showed that the limit of cell divisions is not as constant as originally found but lies between 40 and 80. 25
In contrast to the Hayflick theory, it was recognized that the mean life span of experimental animals is increased about 30–40% by food restriction to about two thirds of ad libitum–fed animals, thus relating aging with environmental factors, 26 –28 for instance, by assuming that aging is connected to a restriction in energy intake rather than that of a specific constituent of food. 29 Besides limited food intake, pharmacological interventions, such as rapamycin, 30,31 also increased the life span in experimental animals.
The second main theory of aging was developed already before the Hayflick theory of aging by Harman 32 in 1956. He stated that life span is determined by the increase of oxidative processes. Taking into account the observation that mitochondria are the main source of H2O2, 33 he later modified his theory to the “mitochondrial theory of aging,” emphasizing that oxygen radicals are produced preferentially in mitochondria and that these species induce oxidation of mitochondrial DNA. As a consequence, mutations should occur. 34
Polyunsaturared Fatty Acids Are Among the Most Oxygen-Sensitive Compounds in Nature and Produce Lipid Hydroperoxides Catalyzed Either by Enzymes or by Radical-Induced Processes
Harman had already suspected a connection between the mortality rate of mice and the degree of unsaturated fatty acids in fats when he registered a relationship between uptake of fats containing different amounts of unsaturated fatty acids with different degrees of unsaturation incorporated in fats. 35 He related this observation to a different tendency of unsaturated fats to undergo free radical reactions. This suspicion has been specified in more detail and related to the content of fats in PUFAs. 36,37
PUFAs are characterized by the presence of one or several structural elements of the form: -CH = CH-CH2-CH = CH-. They are among the most oxygen-sensitive compounds in nature, exemplified by the fact that oils stored in sun light become rancid within a short time. 38 The rancid smell is caused by the generation of volatile lipid peroxidation (LPO) products derived from PUFAs. 39 The energy necessary for oxidation can be even reduced by use of lipoxygenases, 40 present in all cells. Most lipoxygenases require free PUFAs as substrate. All PUFAs occur in resting cells not in free form, but are esterified, such as in fats or phospholipids in mammalian tissues or with sugars in plants. 41
PUFAs are liberated by activation of enzymes, for instance by phospholipase A2. Phospholipase A2 42 cleaves phospholipids of cell membranes specifically in position 2 of the glycerol skeleton of a phospholipid where PUFAs are preferentially localized (Fig. 1). This cleavage reaction produces free PUFAs and lysophospholipids, which activate G-protein coupled receptors. 43

Phospholipids are esters of gycerol with phosphoric acid. The alcoholic group in position 1 is usually esterified to a saturated or monounsaturated fatty acid, in position 2 to a PUFA, and in position 3 to phosphoric acid, which in turn carries an amino alcohol, mainly choline (as shown in the figure) or cholamine.
Free PUFAs are substrates for lipoxygenases. The oxidation of PUFAs within these enzymes occurs in one-electron transfer steps catalyzed by bivalent metal ions, in particular iron ions, as elucidated by Vliegenthart. 44 The PUFA molecule is brought within the enzyme in a particular steric position with the activated homoallylic CH2 group close to the iron ion. The Fe3+ ion of the activated enzyme picks up an electron of the activated C-H bond and becomes a Fe2+ ion. The weakened one-electron C-H bond of the PUFA undergoes cleavage to a proton and a carbon-centered PUFA radical, stabilized by resonance. The generated carbon-centered radical adds one molecule of oxygen in a regio- and stereo-specific manner, forming a peroxyl radical. The latter adds an electron from the Fe2+ ion, generating a peroxyl anion and an Fe3+ ion ready to undergo with the next PUFA a new cycle. The peroxyl anion condenses with a proton by producing a molecule of a hydroperoxy PUFA, which is released from the complex. The radicals involved in the reactions never leave intact enzyme complexes before this reaction sequence is completed (Fig. 2).

The electron transfer reactions in 13-lipoxygenase starts from free linoleic acid and proceeds via formation of radicals to 9-cis,11-cis-octadecadienoic acid.
The reaction in lipoxygenases closely resembles the reaction occurring with PUFA-containing fats when exposed to radiation or sunlight. The sensitive C-H bond of the activated CH2 group of a PUFA is cleaved to form a carbon-centered radical. In contrast to the situation in lipoxygenases, the PUFA is not in a specific steric conformation, and therefore a radical produced by removal of a hydrogen radical from linoleic acid reacts either in position 9 or 13 with oxygen to the corresponding peroxyl radical, resulting in generation of two regioisomers with about equal probability. Each of these regioisomers occurs in two enantioisomeric forms (Fig. 3). Peroxyl radicals abstract an activated hydrogen from a nearby located molecule to form hydroperoxide molecules. Whereas the lipoxygenase-induced LPO reaction is stopped when the amount of free PUFAs is exhausted or by inactivation of the enzyme, nonenzymatic reactions proceed as long as generated radicals are not scavenged. The nonenzymatic reactions can be simulated by exposure of PUFAs to iron ions. 45
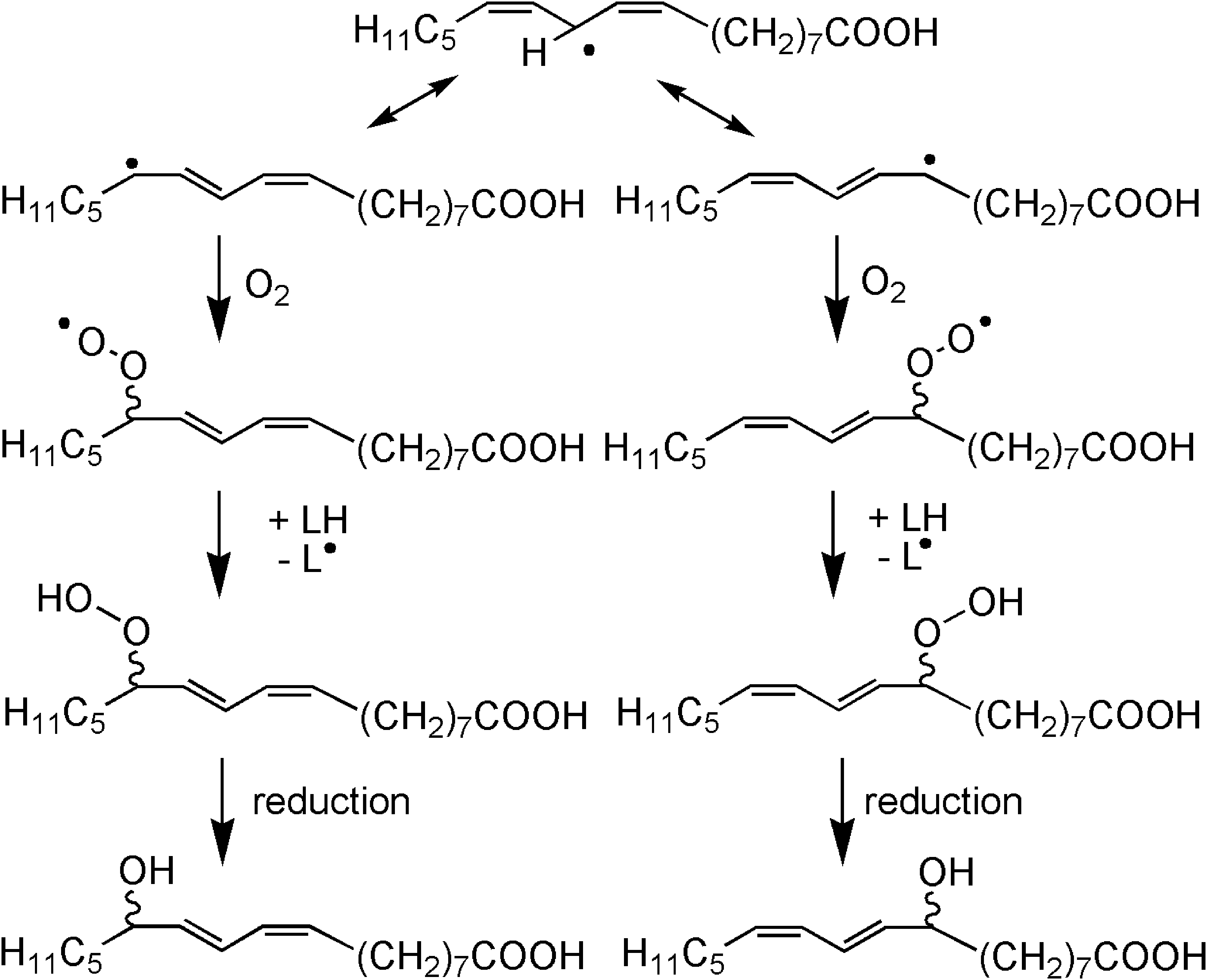
Four isomers are generated by action of radiation on linoleic acid.
Lipid hydroperoxides produced are reduced enzymatically to the corresponding alcohols. The alcohols are, in contrast to the hydroperoxides, sufficiently stable to be separated after appropiate derivatization by gas chromatography and can be distinguished by their mass spectra, allowing us to recognize whether the mechanism of an LPO reaction is enzymatic or nonenzymatic: The observation of a single hydroperoxide resp. its hydrogenation product with a single stereospecific structure indicates that the products are generated by action of lipoxygenases. By contrast, the identification of all possible isomers of PUFAs is typical for nonenzymatic LPO reactions.
LPO Reactions Are Started by Enzymatic Reactions and Switch to Nonenzymatic Ones
The investigation of PUFA oxidation products started with the observation that prostaglandins and related eicosanoids are derived from arachidonic acid by oxidation. Therefore, nearly all research efforts concentrated on eicosanoids. 46 Thus, the investigation of linoleic acid oxidation products was neglected, despite the fact that linoleic acid is the most abundant PUFA in mammals, that it is equally well attacked by lipoxygenases, and that it possesses only one activated CH2 group compared to three in arachidonic acid that are prone to oxidative attack and therefore generates much fewer oxidation products than arachidonic acid, simplifying the separation of formed products. Nevertheless, the importance of linoleic acid and its esters has only been recognized in recent years, although the importance of linoleic acid in serving to produce oxidation products has been known for over 40 years, when Galliard 47 detected that linoleic oxidation products are generated in large amounts after injury in plants and Wills 48 observed similar effects by injury of mammalian tissues.
In an experiment to clarify whether the LPO products are generated by enzymes or produced by the action of radical attack, a liver was cut in two parts. One part was directly homogenized in an aqueous solution; the other was first boiled to destroy enzymes and then homogenized. Only the nonboiled sample was rich in LPO products, whereas merely traces of LPO products were detected in the boiled sample. A product analysis of isomeric hydroxyoctadecadienoic acids in the nonboiled sample revealed the presence of all possible isomers, thus pointing to their generation in the course of a nonenzymatic LPO reaction. 49 However, because boiling destroys enzymes, it was concluded that the LPO reaction in the course of homogenization was induced enzymatically, 49 but later switched to a nonenzymatic reaction. Apparently, this switch starts if the amount of free PUFAs exceeds a certain limit. If this limit is reached, peroxyl radicals formed within the enzyme complex attack bonds between the catalytic iron ion and histidine residues, which keep the catalytic iron ion in its complexed state. The histidine gets oxidized, 50 resulting in weakening the binding of the iron ion in the enzyme's active site. As a consequence the iron ion is released 51 and the enzyme is deactivated. The released iron ion reacts with already generated and released lipid hydroperoxy acid molecules in the course of a Fenton reaction to generate LO• radicals (Fig. 4).
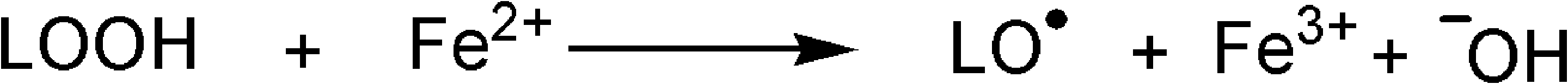
The generation of LO• radicals in the course of a Fenton reaction.
The knowledge that a Fenton reaction stimulates the LPO processes in biological systems allows us to subject a single biological compound to this reaction. The product pattern obtained resembles the products in the biological system but is not disturbed by the reaction products derived by radical attack on the myriad of other biological compounds. If the products formed by attack of radicals on a single biological compound are known, it is much easier to detect these reaction products in tissue samples, because in the whole biological matrix every compound generates a great number of reaction products. 45
Dramatic Differences in the Reactivity Between Nonradical Molecules and Radicals
The radicals discussed here are more reactive than oxidizing molecules by several orders of magnitude. Therefore, LO• radicals cannot be isolated. Their presence can be deduced only by the presence of typical reaction products. Stabilization of LO• radicals occurs either by cleavage, by reaction of the radical site with the adjacent double bond, or by hydrogen abstraction from a nearby molecule. Cleavage of the single carbon–carbon bond adjacent to the C-O• bond produces aldehydes (Fig. 5) and a new carbon radical. The resulting unsaturated aldehydes undergo retro aldol reactions. The newly generated radical usally adds another oxygen atom, forming a peroxyl radical that abstracts a hydrogen from another molecule and is finally enzymatically reduced to the corresponding alcohol. Alternatively, the radical is stabilized by hydrogen abstraction.

Aldehydes are generated by cleavage of LO• radicals. They suffer further decomposition by retro-aldol reactions.
If the radical site of the LO• radical attacks the adjacent double bond, an epoxide radical is produced. This reacts with a second oxygen molecule to generate an epoxy acid after hydrogen abstraction from another molecule (Fig. 6). 45 In the course of hydrogen abstraction by LO• radicals, new radicals are generated. Usually the new radical is formed at a carbon atom. This immediately adds diatomic oxygen to generate a peroxyl radical. This radical may again abstract hydrogen from another molecule, perpetuating the reaction.

Production of epoxy acids by attack of the radical site at the adjacent double bond.
Although the radical reactions are not under enzymatic control, they proceed in a distinct manner, with further hydrogen abstractions being guided by the principle that those processes are preferred that require the lowest amount of energy. Therefore, hydrogens are removed preferentially from activated C-H, N-H, S-H, or O-H bonds. Nevertheless, a tremendous variety of products is generated. Several of the molecules produced may serve after complex formations with proteins to stimulate genes.
Previously the interest of reseachers concentrated mainly on reactions of radicals with C-H bonds of lipids. Thus, for instance, LO• radicals are able to remove the activated hydrogen of a CH2 group from a PUFA in an adjacent phospholipid, resulting in its decomposition either to a truncated phospholipid 52 (Fig. 7) or an epoxy derivative of the phospholipid (analogous to the reaction shown in Fig. 6). In fact, radicals react likewise by hydrogen abstraction from C-H bonds adjacent to an OH or NH group and decomposition to an oxo group under release of a H-O-O• radical, which is a protonated superoxide radical. 53

Generation of a truncated phospholipid by attack of a LO• radical on an adjacent phospholipid.
Lysophospholipids generated by hydrolysis of phospholipids (see Fig. 1) represent substitituted isopropanol derivatives. Because isopropanol reacts in the presence of radicals, as demonstrated for its radiation-induced oxidation to acetone 54 and release of H-O-O• radicals, we have to assume that corresponding oxo compounds are produced by action of radicals with lysophospholipids 53 (Fig. 8).
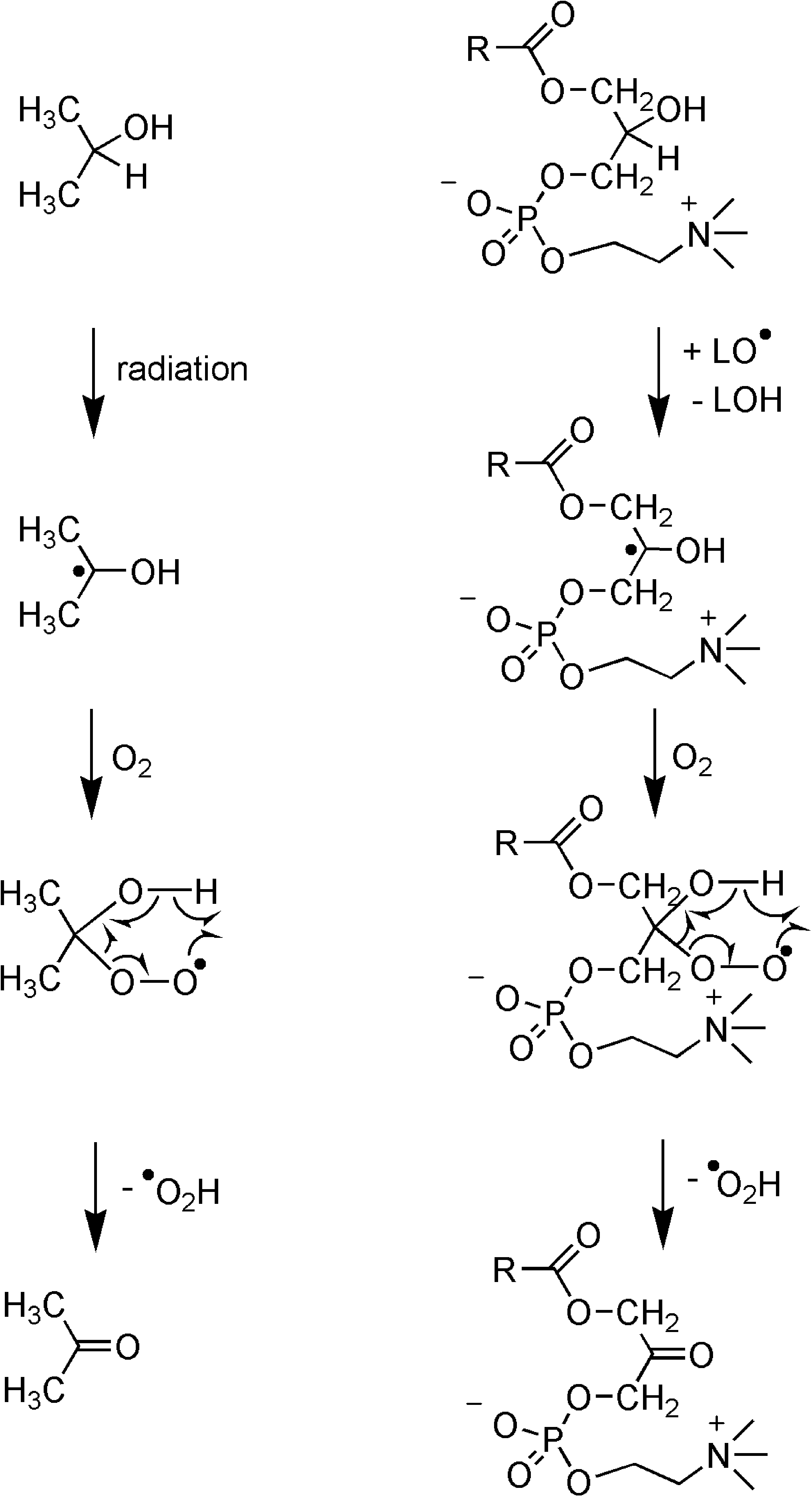
The oxidation of isopropanol to acetone by release of H-O-O• radicals is paralleled by the transformation of lysophospholipids to corresponding oxo compounds by release of H-O-O• radicals.
Similar oxidation reactions of alcoholic groups to keto groups occur in sugars 55 by producing 1,2, 1,3, 1,4, 1,5, and 1,6 dioxo derivatives. 1,4 resp. 1,5 dicarbonyl compounds react with amines and amino groups of lysyl residues by cyclization and water elimination, forming pyrroles resp. pyridine derivatives. Apparently such compounds are constituents of fluorescing lipofuscins. Alternatively sugar can be degraded by a six-membered hydrogen shift reaction (Fig. 9).
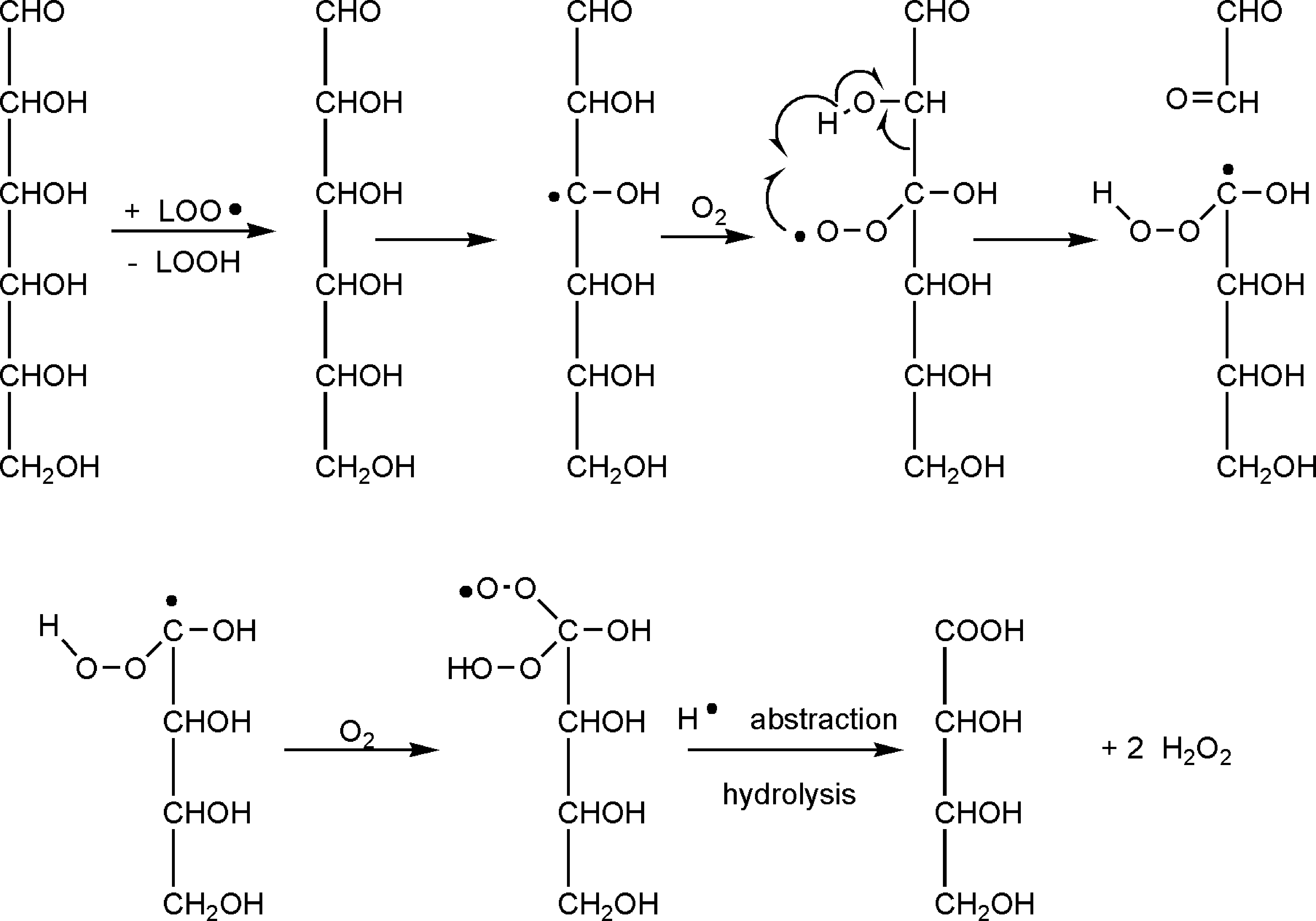
Hydrogen rearrangement via a six-membered intermediate causes degradation of sugar molecules.
These reactions are probably responsible for a loss in immune responses and production of Maillard products. 53 Many other biological alcohols, such as sterols or steroids, and amino acids, such as serine and threonine, contain CH(OH) groups and are prone, like the CH(OH) group in lysophopholipids (Fig. 8), to radical attack. Their probable oxidation partly destroys these biological important molecules, too. Amines such as putrescine or spermidine and the basic amino acid lysine are likewise oxidized to carbonyl derivatives (Fig. 10). 53
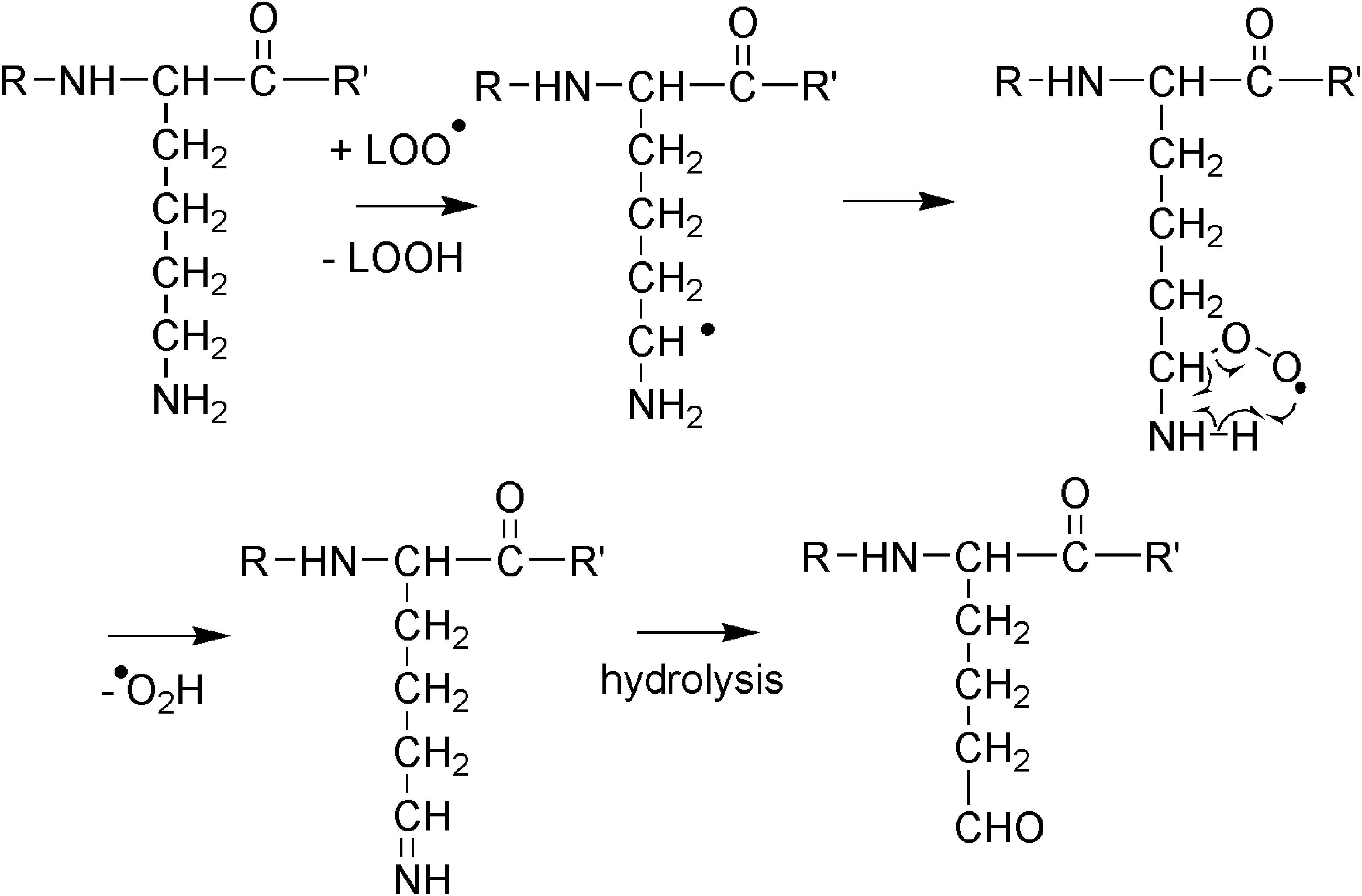
LO• radicals transform lysine to an aldehydic amino acid.
Further investigations revealed that LOO• radicals are—apart from their ability to abstract hydrogen from other molecules—extremely powerful oxidizing reagents. They attack, for instance, any double bond, leading to epoxidation. Thus, the double bond of oleic acid reacts to 9,10-epoxyoleic acid. 56 This epoxy acid is detectable in foods stored for long periods in air, such as cheese. 57
Peroxyl radicals, derived by attack at the activated CH2 group of a PUFA in a plasmalogen (abundant constituents of brain and heart tissues), find the closest available next double bond in the same molecule and therefore attack the enolic double bond. The resulting epoxides are not very stable; they undergo hydrolysis to corresponding diols and further to α-hydroxyaldehydes (Fig. 11). 58

LOO• radicals generated from PUFA-containing plasmalogens oxidize the enolether bond in an intramolecular reaction and easily undergo hydrolysis. Subsequently, they produce α-hydroxyaldehydes.
The knowledge of these degradation products allowed, for instance, detection of the accumulation of these oxidation products in the brain of aged humans 59 and also as a consequence of an artificially induced myocardial infarction in porcine hearts. 60 Peroxyl radicals attack not only single bonds of acids and enols but also those of sterols and terpenes, abundant compounds in all living matter. This induces severe alterations in the structure of biologically important compounds such as cortisone and other steroidal hormones.
Is leakage the Source of Superoxide?
H-O-O• radicals are abundant LPO products, generated, for instance, from alcohols such as lysophospholipids and amines (see Figs. 8 and 10). H-O-O• is a protonated superoxide (O2 •−). Superoxide is transformed to H2O2 by the action of superoxide dismutase. 61 H2O2 was recognized by Boveris et al. 62 to be the main oxidation product in isolated mitochondria. The isolation of mitochondria requires homogenization of tissue. Tissue homogenization is inevitably connected with activation of phospholipases and lipoxygenases, as demonstrated for mammalian tissue by Wills 48 and for plant tissue by Galliard. 47 These enzymes transform PUFAs to lipid peroxidation products. Evidently, Boveris et al. 62 were not aware of these findings and therefore assumed that H2O2 is derived from superoxide (O2 •−) generated in the respiratory system where oxygen is transformed in a four-step electron transfer reaction finally to water. It was further assumed, although never experimentally proven, that the first step of this reaction sequence might occur at an inappropriate location within the respiratory system, from which the generated superoxide (O2 •−) might escape by a “leakage” process. This idea became very popular. 63 Subsequent generations of biochemists continued isolation of mitochondria and other cell constituents as demonstrated by Boveris et al. 62
Unfortunately, the analytical methods usually applied, such as reaction to a colored compound by UV absorption 64,65 or the more sensitive method of reaction to a fluorescent compound, 64,66 or even by spin trapping with electron spin resonance (ESR) spectroscopy, 67 do not allow us to distinguish between superoxide (O2 •−) and its protonated form, H-O-O•, because in an aqueous solution the anion superoxide readily picks up a proton.
Applying these analytical methods the main sites of generation of O2 •− resp. H2O2 were recognized to be complex I 68 and III 69 of the electron transport system. Nevertheless O2 •−and H2O2 were found also to be “byproducts” 69 of a very great number of redox enzymes, such as nicotinamide adenine dinucleotide phosphate (NADPH) oxidase, 70,71 xanthine oxidase, 72 –74 cyclooxygenase, 75 3-phosphate dehydrogenase, 76 ketoglutarate dehydrogenase,, 77 or glycerophosphate dehydrogenase. 78
The author of this review finds it quite unusual that enzymes should produce “byproducts” and even more unusual that different enzymes should generate identical “by-products,” because enzymes act very efficiently without allowing side reactions. These considerations raise the question whether the production of H2O2 might be caused by a reaction connected with the preparation of the cells, as suggested by the investigation of radical production by endothelial cells in the course of reperfusion injury. 79 The suspicion that H2O2 might be generated via release of H-O-O• radicals according to analogous processes, as outlined in Figs. 8 –10, and that superoxide might not derived by a leakage process of the respiratory system is further supported by the following considerations: many enzymes assumed to produce H2O2 suffer like lipoxygenases self-deactivation, for instance, cytochrome c, 80 cytochrome P-45, 81 cycloxygenase, 82,83 NADPH oxidase, 84 3-phosphate dehydrogenase, 85 and also ketoglutarate dehydrogenase. 86 In all redox enzymes, single-electron transfer reactions occur catalyzed by metal ions. Moreover, evidence was provided that the true reagents that cause generation of LPO products are H-O-O• radicals. 87
It is tempting to speculate that the deactivation process resembles the deactivation of hemoglobin and myoglobin by release of iron ions from the enzyme, 88 also demonstrated to cause inactivation of lipoxgenases. 89 In this case, the iron release is induced by an excess of free PUFAs as a consequence of cell wounding. Apparently, the radicals generated within the enzyme attack the bonds, 53 keeping the catalytic metal ions in their complexed form. If the complex is destroyed and if, as a consequence, the bivalent metal ion is released from the enzyme, it can react with LOOH molecules already generated by action of lipoxygenases. Thus, in all cases in which redox enzymes are involved, similar reaction sequences might cause the production of H2O2. All redox enzymes contain bivalent metal ions, which catalyze redox reactions by single-electron transfer reactions and generation of radicals. It is likely that they are, like lipoxygenases, sensitive to an excess supply of substrate and release the metal ion. Because, phospholipases and lipoxygenases are also activated, parallel to the activation of these redox enzymes, LOOHs are generated that serve as substrate for the released metal ions to undergo Fenton reactions and therefore contribute to enhance LPO processes by production of H2O2.
In most cell preparations, mannitol and glucose are added to keep the osmotic conditions constant and to avoid rupture of cells. Mannitol and glucose are sugars, known to produce H-O-O• radicals in large amounts in the presence of LO• radicals. 55 Therefore, they should also contribute to the generation of H2O2. Often, in cell preparation, a minute amount of serum derived from bovine brains is added. Bovine serum contains all of the prerequisites to generate LOOH molecules and certainly contributes also to increasing the concentration of H2O2. Therefore, the generation of O2 •− and H2O2 is probably not induced by a variety of redox enzymes such as cytochrome c 62 or NADPH oxidase, 70,71 but is a consequence of the wounding of cells.
This assumption is further corroborated by a recent investigation using ESR spectroscopy. It revealed that O2 •− is generated only in mitochondria that have experienced a shock, but not in intact ones. 90 Considering that PUFAs are particularly oxygen-sensitive compounds, it is hypothesized that peroxidation of PUFAs is the initiating process of all biological oxidation reactions. Usually it was assumed so far that LPO reactions are initiated by superoxide and not by the reaction of lipid hydroperoxides produced by cell injury as outlined above.
Is the Generation of Radicals Caused by an Alteration of the Cell Structure?
As pointed out by Park, 91 resting neutrophils are dormant and become active only when exposed to the appropriate stimuli. These requirements are probably true for all cells. Generation of LPO products such as H2O2 is not restricted to homogenation, but is recognized also by less severe cell woundings, such as the attack of bacteria 92,93 and viruses. 94 Moreover, activation of phospholipases and lipoxygenases was induced by treatment of cells with ionophores. Thus, addition of the ionphore A23187 42,95 causes the liberation of arachidonic acid by cleavage of phospholipids and its conversion by similarly activated cyclooxygenase to prostaglandins. 12-O-tetradecanoylphorbol-13-acetate (TPA) and other phorbol esters 96 as well as bacterial-derived lipopolysaccharides, 97 including zymosane, 98 stimulate cells to induce enzyme activation. Moreover, the nonapeptide antibiotic leucinostatin A acts as an ionophore. 99 In addition, a multitude of other antibiotics 91 or alkaloids, 100 polar organic compounds such as paraquat, 101,102 and also nonpolar compounds such as polycyclic aromatic hydrocarbons, 103 some solvents, such as carbon tetrachloride, 104 and detergents, 105,106 as well as inorganic compounds such as salts of heavy metal ions such as Hg2Cl2 107 or quartz dust, 108 even excercise, 109 or heat stress, 110 induce activation of enzymes.
Activation of superoxide resp. production of H2O2 is not restricted to enzymes of mammalian cells but was also recognized in those of other animals, e.g., in snail hemocytes 111 and especially in plant cells. Their membranes consist mainly of phospholipids and phosphoglycosides. In response to stimuli or stress they activate phospholipases and lipoxygenases like mammalian cells and produce similar LPO products as mammals. For instance, LPO product generation is reported by stress caused by bacterial attack, 112 by water, 113,114 drought-, 115 salt-, 116 heat-, 117 chill-stress, 118 or by addition of salts containing a bivalent metal ion, such as CuSO4. 119 LPO products are also generated when plants are treated with ozone 120,121 or come into contact with peptides 122 or by light 123 ; even germination induces LPO processes. 124
The very different compounds and events that cause the generation of peroxides seem to have a common root. Severe wounding of cells by homogenation, and also less severe wounding by attack of bacteria, changes the membrane structure of cells. Such a change in cell structure also occurs by contact of the cell membrane with detergents or solvents that dissolve the phospholipid layer, such as carbon tetrachloride. The polar membrane also reacts by addition of metal salts or polar organic compounds such as alkaloids. Changes in the membrane structure also change the ion channels crossing the membrane. Therefore, a relation between activation of ion channels, Ca2+ ions, generation of LPO products, and membrane interactions is recognized. 125 As a result of a change in membrane structure, Ca2+ ions influx into the cell is observed as the earliest event occuring after injury. 126,127 Ca2+ ion influx into the cell causes activation of enzymes such as phospholipases and lipoxygenases 128 and consequently induces lipid peroxidation processes that might be stopped by Ca2+ ion antagonists. 129 The activation is not restricted to these enzymes. A great number of degrading enzymes is activated by cell structure alterations, for instance, deoxyribonucleases, 130,131 peroxidases, 132 proteases, 133,134 as well as spingomyelinase, 135 polygalacturonase, and cellulase, 136 and many others.
Injury induces the hypersensitive response in plants by degrading attacked cells to kill the invader by starvation. 137 Considering that injury is connected with the activation of degrading enzymes, it is tempting to assume that this might also be the process that induces the release of those enzymes that attack the telomeres at the caps of DNA. Thus, the Hayflick limit seems to be largely caused by the induction of DNA degradation by radicals. Therefore, the Harman and Hayflick theories have apparently identical roots, the activation of degrading enzymes by cell membrane alterations.
The degradation reactions induced by cell membrane alterations apparently proceed in two steps. First, the reaction proceeds in a controlled manner, resulting in the programmed cell death by cleavage of the DNA at selected bonds to a ladder of DNA fragments. 134 When the severity of injury exceeds a certain limit, a noncontrolled radical process is induced by cleavage of any bonds that connect the different DNA building blocks. This resembles the switch from enzymatic to nonenzymatic LPO.
After severe cell damage activation of degrading enzymes such as phospholipases and lipoxygenases induced by the influx of Ca2+ ions, and the switch to a nonenzymatic reaction by production of radicals at the surface of cells, the resulting radicals come into contact with the phospholipids of adjacent cells and react there by oxidation of PUFAs incorporated into phospholipids. Thus, the initiated radical reaction is able to proceed like an infection from one cell to the next, until finally a radical is scavenged.
Radical Scavengers
Usually a radical reacts with an adjacent molecule by hydrogen abstraction, producing a new radical. This chain reaction is only interrupted if the resulting radical has a lifetime long enough to meet a second radical to generate a new nonradical molecule. Radicals with a long lifetime are often generated by hydrogen abstraction from the SH group of thiols, for instance glutathione, which is usually present in large amounts in cells.
Because plants are exposed much more than animals to radiation, they have developed a great variety of scavenger compounds, especially with phenolic skeletons, such as flavonoids, 138 because the generated radicals of phenols are resonance stabilized. The scavenging property of simple phenols is enhanced by additional phenolic groups, such as in resveratrol. 139 The phenolic compound may react in its different resonance structures with a second radical of this type by forming oxygen–carbon or carbon–carbon bonds, giving rise to lignification. These molecules enhance the cell walls and help to prevent the attack of predators. Red wine is known to contain resveratrol. Therefore, it has been recommended to drink red wine, in which resveratrol is enriched. 140 Because resveratrol seems not to be absorbed in the blood, however, the ability of red wine and other phenolic compounds to act as scavengers seems questionable.
On the other hand, all plants generate furan fatty acids, which also act as radical scavengers. 141 These compounds, instead of PUFAs, are incorporated into the phospholipids of mammals 142 and therefore seem to fulfill the requirements to act as potent scavengers in humans. Another scavenger molecule with these properties containing an enlarged resonance system is vitamin E.
In recent years, several compounds, such as plasmalogens, 143 have been denoted as scavengers because they disappeared in the presence of radical-producing compounds. In fact, as shown above (Fig.11), they are transformed by radicals into reactive compounds (α-hydroxy aldehydes) 58 that suffer dehydration to α,β-unsaturated aldehydes. These react with nucleophiles, e.g., glutathione, by a Michael condensation 144,145 and thus reduce the body's own supply of scavenger compounds. Likewise ascorbic acid reacts as a sugar derivative by generation of aldehydic degradation products 55 and is therefore also not able to scavenge radicals efficiently.
Aging and Age-Dependent Diseases
As pointed out above, any change in cell structure induces enzymatic LPO reactions, which switch by severe alterations to nonenzymatic ones. These nonenzymatic LPO reactions generate peroxyl radicals, which not only attack phospholipids but also important proteins, sugars (for instance, those responsible for immune reactions), hormones, and even DNA. These changes in cell structures and enzymes, which are responsible for regeneration, slowly decrease the function of organs and contribute to normal aging, recognizable by an increase of LPO marker compounds, 2 including products of the Maillard reaction. 13
The spread of radicals from cell to cell induces the events observed in inflammatory diseases. Thus, LPO products were detected in increased amounts in atherosclerosis, 146,147 diabetes, 148,149 rheumatoid arthritis, 150 asthma, 151 cataractogenesis, 152 psoriasis, 153 Alzheimer disease, 154,155 Parkinson disease, 156 and multiple sclerosis. 157,158 Dramatic increases in LPO marker compounds were also observed after sudden events combined with severe cell damage, e.g., after shock, 159 burn injury, 160,161 myocardial infarction, 60 or stroke. 162 Nevertheless, such findings must be assessed with care, especially if tissue was homogenized without considering that this manipulation induces severe LPO oxidation processes. For instance, an issue to consider is if workup of tissues is carried out by homogenization using aqueous solvents or instead by adding an organic solvent, which prevents activation of degrading enzymes.
In the above list of the relationships between age-dependent diseases and LPO, cancer is deliberately not included. There are hints that cancer might be induced by a deficiency in production of radicals that contribute to the generation of LPO signals that stop growth. Thus, it was expected to find increased amounts of LPO markers in cancer tissues. In contrast, a comparison of the LPO markers in unaffected and cancer-carrying breast tissues revealed a lower amount of markers in the affected parts of a single breast than in the unaffected parts. 163 This effect increased with the aggression of the disease. In this connection, it seems useful to mention that compounds with an α,β-unsaturated carbonyl system are used as drugs to fight against cancer, 164 and α,β-unsaturated aldehydes are the first degradation products in the radical-induced decay of hydroperoxides, which react further in Michael-type reactions. 144
Conclusions
The increase in products derived by generation of radicals seems to cause aging and age-related diseases. In contrast to previous assumptions, evidence is provided that radicals are apparently not produced by a “leakage” process by which superoxide is released from the respiratory chain, but that radicals are derived by oxidation of PUFAs in a two-step reaction, first by enzymatic LPO, which switches to a nonenzymatic one as a response to any changes in cell structure. It is hypothesized that this is, in fact, the only way in which radicals are generated. The initially generated LO• radicals transform other molecules into peroxyl radicals, which release protonated superoxide radicals and attack other molecules, thus destroying lipids, sugars, nucleic acids, hormones, and proteins, leading to a few typical degradation mechanisms and contributing to a slow decrease in body functions. Thus, the different theories of aging, including the Hayflick limit, can be explained.
Footnotes
Acknowledgments
I am very grateful to my former collaborators A. Batna, S. Bauer, A. Dudda, C. Fuchs, P. Gorst-Alman, K. Hannemann, M. Herold, M. Jande, M. Jahn, W. Jira, W. Kern, U. Kießling, W. Knörr, P Kreil, A. Loidl- Stahlofen, A. Lutz, C. Mayer, W. Meyer, A. Mlakar, V. Puchta, J. Reiner, J. Schmidt, R. Schödel, H. Spreitzer, my three sons Dieter, Michael, and Peter Spiteller, U. Walther, H. Weiss, M. Weisser, and H. Ziegler, who contributed to my research about lipid peroxidation and furan fatty acids. I also thank the Deutsche Forschungsgemeinschaft, Fonds der Chemischen Industrie, Schering AG, Henkel AG, and Fischer Stiftung for many years of generous financial support.