
Abstract
In spite of enormous efforts and accumulated knowledge, our capabilities for tackling aging and age-related diseases (ARDs), and ultimately to promote longevity, are still very modest. What is lacking—essential data on key players, efficient analytic tools, or both? Here we discuss how the existing data may be integrated and analyzed in the context of microRNA (miRNA)-regulated protein–protein interaction networks. The proposed model highlighted: (1) The strong molecular links between aging, longevity, and ARDs; (2) the possibility and even the preferability of initiating longevity-promoting interventions in adult life; (3) the potentially important role for miRNA- (or small interfering RNA [siRNA]) mediated targeting of certain genes with features of antagonistic pleiotropy; (4) the superiority of systemic interventions to the common single-target approach in curing ARDs and promoting longevity.
Introduction
Aging was suggested as the main risk factor for the development of ARDs. 5 We believe that ARDs not only are direct consequences of aging but actually represent its diverse manifestations, being an essential component of the “normal” aging process. 1,6 This means that common molecular mechanisms stand behind both aging/longevity and ARDs. These mechanisms could be greatly related to the epigenetic control of gene expression, 6 –10 which includes DNA methylation, histone modifications, and a recently discovered mechanism of RNA interference (RNAi). The effector molecules in RNAi are microRNAs (miRNAs), small double-stranded RNAs that cause gene inactivation at the posttranscriptional level by binding to mRNAs, with their consequent degradation or repression of translation (translational silencing). The rapidly emerging (though still incomplete) data in the field indicate that changes in the patterns of epigenetic regulation or in its components appear to have a profound impact on both longevity and ARDs. 10 –14 For example, the possibility to extend life span by modulating gene expression through RNAi in Caenorhabditis elegans was shown recently. 15
The extremely high complexity of the subject requires appropriate analytic tools. During the last years, a network-based approach has been increasingly used for the analysis of aging, longevity, and ARDs. 2 –4,16 –25 In particular, the analysis of protein–protein interaction (PPI) networks showed numerous molecular links between different ARDs 26 and between ARDs and longevity. 4 Apart from the contribution to a better understanding of the mechanisms of aging, the network approach could be useful for developing new research strategies in the field and searching for potential pro-longevity targets. One of the important conclusions from the network analysis is that multitarget interventions could be much more efficient for combating aging and ARDs than the currently dominant approach, which focuses on highly specific single-target drugs. 4,27 In this work, we analyzed how the miRNA-regulated PPI network common for longevity and ARDs could help in searching for pro-longevity targets.
Materials and Methods
The construction of the networks was described in detail elsewhere.
2,4,28
Briefly, given that the LAGs act in cooperative manner, we have recently constructed the PPI Human Longevity Network (HLN)
2
(for updated HLN, see
Network analysis, including simulations with randomly selected genes/proteins, was performed using YABNA (Yet Another Biological Networks Analyzer), a flexible set of tools developed in our laboratory. The graphical output of networks was generated using Cytoscape 2.6.0. 33
All statistical calculations were performed using the Statistical Package for the Social Sciences Inc. (SPSS; Chicago, IL) software. For comparison of observed and expected values, the chi-squared test was used. The differences were considered significant at p value < 0.05. To compare the average values of connectivity between different groups of genes, the number of PPIs (k) for each gene (protein) was ln-transformed.
Results and Discussion
The Common Gene Signature network is enriched in potentially antagonistic pleiotropy genes
An overlap between HLN and the networks of the major ARDs (Common Gene Signature; CGS) represents a single PPI network consisting of 643 nodes
4
(for the last update see
Detailed analysis of connectivity and functions of the CGS genes was described in our recent paper. 4 Here we would like to stress that among them are many genes that are essential for development and growth. On the other hand, being associated with ARDs, these genes may also have detrimental effects later in life. By definition, this fits very well Williams's idea of antagonistic pleiotropy. 34
More specifically, over one third (39%) of the CGS genes were found as being essential (Table 1). This value is more than two-fold higher than that for the entire human interactome (18%; p < E−25). Furthermore, over half of the LAGs in CGS are essential (56%). This ratio is much higher than that for the LAGs in the interactome (30%). Of note, the high essentiality of the CGS genes is not biased by the relatively large number of essential LAGs in the CGS. This follows from the fact that the percent of essential non-LAGs (genes that have not thus far been identified as LAGs) and the percent for all essential genes in the CGS network are similar (36% and 39%, respectively). Thus, CGS is considerably enriched in essential genes and also in essential LAGs.
Chi-squared test showed a highly significant difference between LAGs and other genes and between all CGS gene sets and their corresponding sets in the interactome; p < E−25.
CGS, Common Gene Signature; LAGs, longevity-associated genes.
Another remarkable (though expected; see ref. 4) feature of the CGS genes and essential genes in particular is that they have a very high average number of PPIs (ln[k] ± standard error of the mean [SEM] = 2.82 ± 0.04 and 3.08 ± 0.06, respectively; for example, in the interactome these values are 1.27 ± 0.01 and 1.75 ± 0.03, respectively). The combination of a high degree of essentiality and connectivity further strengthens the probability for the CGS genes to have features of antagonistic pleiotropy. In support of this are several lines of observations. Protein connectivity significantly correlates with the degree of essentiality 16 and pleiotropy. 17 Also, the more connected proteins are more likely to be associated with aging, 17,18 diseases, 26 or both. 2,3,25
Not only the essential genes in CGS but also the nonessential ones are highly connected (ln(k) ± SEM = 2.66 ± 0.05). With this in mind, one could ask if in fact there is evidence for their “nonessentiality.” Indeed, the list of essential genes is still far from being complete. Moreover, Zotenko et al. 35 showed that the essential genes tend to be more connected with other essential genes. In CGS, the nonessential genes have numerous connections with essential ones. Therefore, it could be suggested that among these currently considered nonessential genes many in fact will be found to be essential, with potential features of antagonistic pleiotropy.
Expression of many CGS genes is miRNA regulated
As a step toward constructing an integrative PPI–epigenetic network, we included miRNAs that have experimentally validated targets in CGS. The resultant miRNA-regulated PPI network is shown in Fig. 1. Thirty one miRNAs were found to target 77 genes in CGS, which in turn directly interact with other 360 CGS genes. That is, two thirds of the CGS genes could be miRNA regulated, directly or indirectly through PPIs of their target genes. Of note, all of these miRNAs were reported to have an altered expression in at least one ARD. 10,31 The observed involvement of corresponding genes in ARDs could to a great extent be the result of an altered miRNA expression.
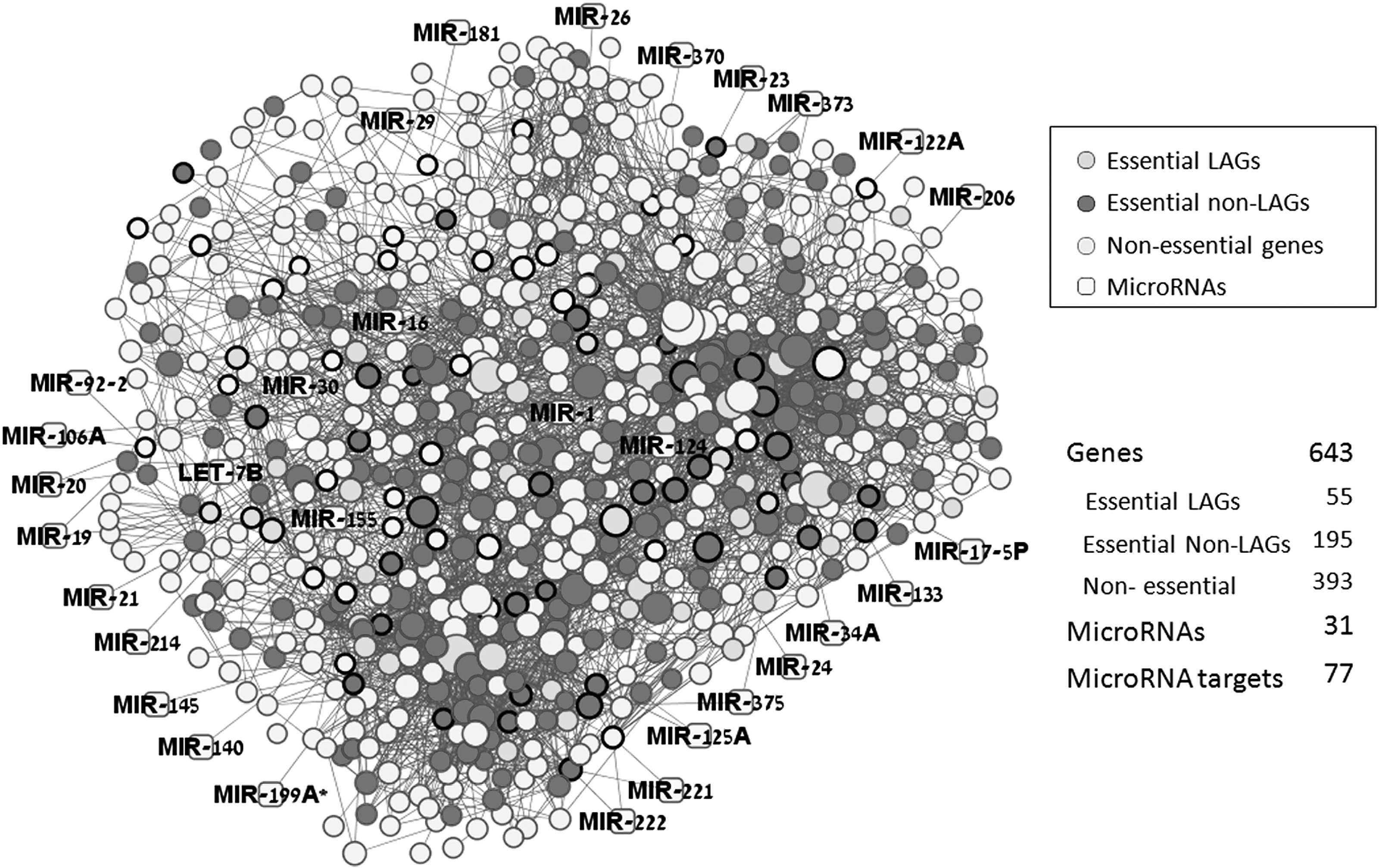
The microRNA (miRNA)-regulated Common Gene Signature (CGS) network. Presented is a graphical representation of the gene overlap between the human longevity network and the networks of major age-related diseases. Node size is proportional with the number of protein–protein interaction (PPIs) in the interactome. Nodes with black borders represent miRNA targets. LAGs, Longevity-associated genes.
When the miRNAs and their targets were “extracted” from CGS, they still formed a continuous network in which the genes are connected via either PPIs or miRNAs (Fig. 2). This is quite an interesting result, in view of the relatively small number of nodes. As seen in Fig. 2, the expression of several miRNAs could be also regulated by genes from the network. 32 For example, miR-21 and PTEN might form a typical feedback loop. Other cases include more complicated relationships. For example, SP1 was reported to influence the expression of miR-106A, which may target SP1 via its neighboring partner RB1. A similar “triangle” loop is observed for CCND1, miR-17, and NCOA3.

The network formed by microRNAs (miRNAs) and their targets in Common Gene Signature (CGS). With only two exceptions, all target genes in CGS are interconnected via either protein–protein interaction (PPIs) or miRNAs connections. Shown in brackets for each miRNA is the total number of targets in the interactome. LAGs, Longevity-associated genes.
Because the list of miRNA validated targets is far from being complete, this network includes a limited number of genes from the CGS. Nevertheless, the constructed network is important in view of possible pro-longevity interventions. Indeed, miRNAs (or small interfering RNAs [siRNAs]) represent a promising tool for controlled modification of gene expression, and the important point is that this could be done postdevelopmentally 15 or even later in life, including the postreproductive period. This approach appears to be much more reasonable, allowing for avoiding hardly predictable and potentially undesirable effects of gene inactivation on growth and development. Then, the essential CGS genes, and particularly those with multiple connections, could be the subject of RNAi. With this in mind, a group of 35 genes from the miRNA-regulated CGS network (Fig. 2), which are neighboring partners of LAGs, could be of particular interest. First, they are essential. Second, most of them were found in association with at least one ARD. This shows that they have features of antagonistic pleiotropy, and therefore could be relevant to aging and longevity. Consequently, these genes could be considered new candidates for pro-longevity interventions with the potential of being modulated through their miRNAs (siRNAs) later in life.
Concluding Remarks
It is reasonable to suggest that a well-balanced activity of CGS nodes including both miRNAs and genes/proteins would assure a favorable healthy status for the whole organism. Besides, the network organization offers the potential for self-compensation, which is very similar to the concept of self-stabilization, a highly desirable property of fault-tolerance systems in distributed computing. 36 A distributed system that is self-stabilizing will end up in a correct state, after a finite number of execution steps, no matter what state it is initialized with and no matter what execution steps it will take. In biological systems, when the network reaches a certain degree of misbalance and self-compensation is not efficient enough, pathological conditions will manifest. It is quite obvious that concomitantly correcting all of the changes in the network is unrealistic. Moreover, it is unnecessary. Instead, we suggest that the restoration of an essential part of the network (“the network fingerprint”) could be sufficient for stabilizing and bringing it to a self-compensatory state.
As a first approximation, the network consisting of miRNAs and their validated targets (see Fig. 2) could be used as such “fingerprint” for CGS. Although the “fingerprint” expression profile may significantly differ among various tissues, it could provide useful information regarding the functional state of the CGS and suggest possible ways for corrections.
Footnotes
Acknowledgments
This work was supported by the European Union FP7 Health Research grant number HEALTH-F4-2008-202047 (to V.F.).
Note Added in Proofs
At the time of receiving page proofs, two related articles, one reviewing network-based approaches in aging research
37
and another one on aging-related miRNAs
38
were in press. Of note, all the four miRNAs described in this article
38
are found in our miRNA-regulated CGS network (Fig. 1) and in the “Fingerprint network” (![]() ).
).