
Abstract
Objective:
Clinical trials of cardiac cell therapy have indicated limited benefits in aging patients, even though preclinical studies using young animals consistently reported significant improvements. Animal studies have demonstrated reduced efficacy of donor cells isolated from older individuals. Here, we evaluated the effects of donor age on the function of human mesenchymal stem cells (hMSCs) in the context of cell therapy for ischemic cardiomyopathy.
Methods:
In vitro, we compared the growth and clonogenic potential of hMSCs isolated from young or old patients (1–5 vs. 50–70 years old). In vivo, we injected young or old hMSCs (2.0 × 106) (or medium) into the infarcted myocardia of immunosuppressed rats immediately after coronary artery ligation (myocardial infarction [MI]). We assessed cardiac function (echocardiography) at 1, 2, and 4 weeks after MI, and myocardial matrix metalloproteinase-2 (MMP-2), MMP-9, and tissue inhibitor of matrix metalloproteinase-3 (TIMP-3) levels at 1 week.
Results:
In vitro, growth and colony-forming unit fibroblast (CFU-F) formation were markedly diminished in old hMSCs (p < 0.001 and p < 0.05, respectively, vs. young). In vivo, compared with old hMSCs or medium, young hMSCs best preserved ejection fraction, fractional shortening (p < 0.05), and left ventricular end-diastolic and end-systolic volumes (p < 0.01). Recipients of young hMSCs also exhibited increases in vascular density and TIMP-3 protein levels and activity (p < 0.05), and decreases in MMP protein levels and activity (p < 0.05).
Conclusions:
The regenerative capacity of hMSCs was significantly influenced by age. Transplanting young hMSCs improved functional outcomes after an MI by preventing matrix degradation and promoting angiogenesis. The clinical implication is that aged patients require an optimized source of stem cells for treatment.
Introduction
Human mesenchymal stem cells (hMSCs) have the important properties of self-regeneration and differentiation plasticity. 2,3 Numerous studies have demonstrated that stem cell therapy with MSCs is safe and may be efficacious for patients who have suffered significant cardiac injury. 4,5 Several preclinical studies found that implanted MSCs improved cardiac function after an MI. 4,6 However, the initial clinical trials of stem cell therapy after an MI failed to reproduce the substantial benefits demonstrated in the preclinical animal studies, especially in elderly patients. 7,8 Some reports suggested that donor age was the principal reason for the difference between the preclinical and clinical findings. 9 –11 Indeed, multiple influences of age on stem cell function have been confirmed, including effects on stem cell number, 12 proliferation potential, 13 senescence and apoptosis, 14 replicative capacity, 15 and in vitro differentiation potential. 16 In animal experiments, ex vivo growth was less robust in stem cells from older donors compared to young donors, and old cells did not confer the same benefits as young cells after implantation into an experimental infarct. 11,17 At present, few data exist to confirm that stem cells from elderly human patients will not function as well as those from younger patients. This study directly assessed the effects of age on the function of hMSCs in the context of cell therapy after an MI.
Methods
Animals
Adult male Wistar rats (200–250 g) were obtained from the Animal Central facility of the Second Affiliated Hospital of Harbin Medical University, Heilongjiang, China. Animal procedures were approved by the University's Committee for the Care of Experimental Animals; all animals received humane care in compliance with the Principles of Laboratory Animal Care (National Society for Medical Research) and the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Bone marrow collection
The research proposal for human bone marrow collection was approved by the Second Affiliated Hospital Research Ethics Committee of Harbin Medical University. Bone marrow aspirates were obtained from the sternums of patients undergoing cardiac surgery at the Second Affiliated Hospital of Harbin Medical University after obtaining the patients' written consent according to the principles outlined in the Declaration of Helsinki. Informed written consent was obtained from each adult patient or from the parents of pediatric patients. “Young” bone marrow was collected from patients (aged 1–5) with congenital heart disease, while “old” bone marrow was obtained mainly from patients (aged 50–70) with valve disease. Patients with cyanosis, hepatitis, severe organ dysfunction, or pulmonary hypertension were excluded. All bone marrow aspirates were mixed immediately with heparin.
Cell isolation, culture, and identification
Bone marrow mononuclear cells were separated by centrifugation with a Ficoll-Paque gradient (1.077 g/mL density; GE Healthcare, Kretztechnik, Zipf, Austria), seeded into 75-cm2 culture flasks in Iscove modified Dulbecco medium (IMDM; containing 10% fetal bovine serum [FBS] and antibiotics), and incubated at 37°C in 5% CO2. After 48 h, nonadherent cells were removed by changing the medium. Thereafter, the medium was changed every 2–3 days, and adherent cells were cultured until they reached 80% confluence. The hMSCs were harvested by trypsinization (0.25% trypsin with 0.02% EDTA) and passaged. Passage 3 cells were immunostained with antibodies against CD29, CD90, CD105, CD34, CD45, and CD133 18 to identify hMSCs, and the positive cells were quantified using a flow cytometer (FACSCalibur, Becton Dickinson, Franklin Lakes, NJ and San Jose, CA).
Cell growth characteristics
hMSCs were isolated from equal amounts of young and old bone marrow samples, and then cultured in 75 cm2 culture flasks. The hMSCs were serially subcultured under standard conditions to the third passage. When cells were 80–90% confluent, they were trypsinized and seeded into 96-well plates at a starting density of 1000 cells per well. Cell growth was evaluated with a 3-(4,5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) colorimetric assay at 3, 5, 7, 10, and 12 days after plating. Growth curves were generated for young and old hMSCs and compared. 19
Bone marrow subpopulation clonogenic potential
A colony-forming unit fibroblast (CFU-F) assay was used to evaluate and compare the clonogenic potential of young and old hMSCs. Briefly, both young and old cells were seeded in six-well plates (1 × 106 per well) and cultured for 7 days in MesenCult medium (Stem Cell Technologies). CFU-F was identified using Wright–Giemsa staining, and cloning efficiency was calculated and compared among groups. 11
Myocardial infarction and cell implantation
To prevent immunorejection, all animals received Cyclosporine A (5 mg/kg, given intraperitoneally) each day from 3 days before cell implantation until the end of the experiment. The rats were anesthetized with 10% chloral hydrate (3.0 mL/kg), and respiration was assisted with 100% O2 via a ventilator (Harvard Inspira ASVp, NatureGene Corp., Medford, NJ) and adjusted by weight. Through a left lateral thoracotomy, an MI was created by ligating the proximal left coronary artery (LAD) with a 5-0 polypropylene suture. Myocardial damage was confirmed by a change in regional color (blanching) and a decrease in regional contraction. Fifteen minutes later, young or old hMSCs in suspension (2 × 106/120 μL) were injected into the center of the injured myocardium and into two points around the margin of the infarcted region. The same volume of culture medium was injected into the same locations in the control group. After the incision was closed, 150,000 U/mL of penicillin G was administered intraperitoneally each day.
Left ventricular function
Function was evaluated by echocardiography prior to and at 1, 2, and 4 weeks after MI and cell or medium injection in all three groups (recipients of medium, old hMSCs, young hMSCs). Left parasternal images were obtained in the right lateral decubitus position using a 12-MHz transducer (Vivid 7, GE, Oceanside, CA). Short-axis two-dimensional images were stored as digital loops and M-mode images were made at the midpapillary level of the left ventricle. The left ventricular dimensions at end diastole (LVEDD) and end systole (LVESD), left ventricular end-systolic volume (LVESV), and end-diastolic volume (LVEDV) were measured. For each measurement, three consecutive cardiac cycles were traced and averaged by an experienced examiner in a double-blinded fashion, so that any variability due to the position of the probe was equivalent in all groups.
Reverse transcription polymerase chain reaction
Hearts from each group of recipient rats were harvested at 1 week after injection. Tissue RNA levels of TIMP-3, MMP-9, and vascular endothelial growth factor (VEGF) were detected by reverse transcription polymerase chain reaction (RT-PCR). Total RNA was extracted using the TRIzol reagent (Gibco BRL/Invitrogen, Carlsbad, CA), and cDNA was composed using a Reverse Transcription System Kit (Promega) according to protocols listed in the instruction manual. A 20 μL volume (2 μL of cDNA product, 2 μL of 10 × PCR buffer, 0.4 μL of 10 mM deoxyribonucleotide triphosphates (dNTPs), and 1 U of Taq enzyme) was mixed in the presence of 0.1 μL each of 5′ and 3′ primer (10 mM) for PCR. PCR was performed for 30–35 cycles (each cycle: 95°C for 30 s, 53–60°C for 30 s, and 72°C for 30 s), with an additional, 7-min incubation at 72°C after completion of the last cycle. The primers used were as follows: TIMP-3 forward, 5′-GCC GTT TAT GGA GTT GAT-3′, reverse, 5′-AGC ATT GAG CAG GGT AGA-3′, 229 bp; MMP-9 forward, 5′-GCA CGG CAA CGG AGA AGG C-3′, reverse, 5′-CCG TCG CTG GTA CAG GAA GAG-3′, 257 bp; VEGF forward, 5′-CCC ACG ACA GAA GGA GAG CA-3′, reverse, 5′-GCA CAC AGG ACG GCT TGA A-3′, 150 bp. The PCR products were size-fractionated using 2% agarose gel electrophoresis and compared by density intensity.
Protein levels and activity
At 1 week after injection, protein extracts from the rat hearts were analyzed for the levels of MMP-9, TIMP-3, and VEGF by western blot. Briefly, frozen heart tissue samples were processed by grinding and using protein lysis buffer. Equal amounts of protein from left ventricular extracts were applied to 12% acrylamide gels and electrophoresed. The samples were then electroblotted onto polyvinylidene difluoride membranes (PVDF; BioRad, Hercules, CA) and probed with antibodies against TIMP-3 (1:100; ab66022; Abcam, Cambridge, MA) or MMP-9 (1:100; ab58803 Abcam), or VEGF (1:100; sc-7269, Santa Cruz Biotechnology, Inc., Santa Cruz, CA). The antigens were detected using enhanced chemiluminescence. After membrane switch, the levels of MMP-9, TIMP-3, and VEGF were compared among groups by density intensity. The activities of myocardial MMP-2, MMP-9, and TIMP-3 were quantified using gelatin zymography and reverse-gelatin zymography, 20 –22 and then compared among groups by intensity of the strips.
Vascular density and cell survival
To assess vascular density, left ventricular tissue sections were immunostained with an antibody against CD31 (platelet endothelial cell adhesion molecule-1 [PECAM-1], clone MEC13.3, BD Biosciences Pharmingen, San Diego, CA) at 4 weeks after injection. Vascular density in the border zone area was compared among the three groups. The number of capillary structures (PECAM-1–positive vessels) per high-power field (0.2 mm2) was determined in 10 randomly selected fields per section within the scar area, in a blinded fashion, with a Nikon fluorescent microscope.
Myocardial tissue sections were immunostained with an antibody against human mitochondria (MAB1273, Millipore, Billerica, MA) at 4 weeks after injection. Where positive cells were visible in the border zone, infarct, and remote regions, those sections were randomly imaged using the Aperio virtual microscope system. Surviving human cells (positive staining) were quantified as described above for vascular density (number of positive cells per 0.2 mm2).
Collagen and ventricular remodeling
To quantify collagen in myocardial infarct regions at 4 weeks after injection, paraffin-embedded heart sections were subjected to Masson trichrome staining. Areas of collagen in five randomly selected fields from the infarct region were averaged, and the collagen area was expressed as a percentage of the total section area.
Statistical analyses
All data are expressed as mean ± standard deviation (SD). Analyses were performed using the SPSS software package for Windows (version 13.0; SPSS, Chicago, IL). Independent samples t-tests compared the effects of donor age on hMSC surface antigens, human mitochondria positive cell numbers, and CFU-F numbers in vitro. A factorial analysis of variance (ANOVA) compared growth curves among groups, and repeated-measures ANOVA compared the effects of cell treatment (medium, young hMSCs, old hMSCs) on echocardiographic variables. All other data (including vascular density, collagen content, RT-PCR, results from western blot, and zymography) were compared among groups using a one-way ANOVA. If the F-ratio was significant, pairwise tests of individual group means were carried out using the Student–Newman–Keuls test. p < 0.05 was considered statistically significant.
Results
Characteristics of cultured hMSCs from young and old donors
Morphologically, the hMSCs from young and old donors were similar (Fig. 1). Using flow cytometric analysis with antibodies against cell-surface markers to determine the composition of the cells, we found that, regardless of donor age, over 95% of hMSCs expressed CD29, CD90, and CD105, whereas less than 2% of the cell population expressed leukocyte common antigens, CD45, or the hematopoietic lineage markers CD34 and CD133 (n = 6/group). There were no differences between young and old hMSCs (p > 0.05).
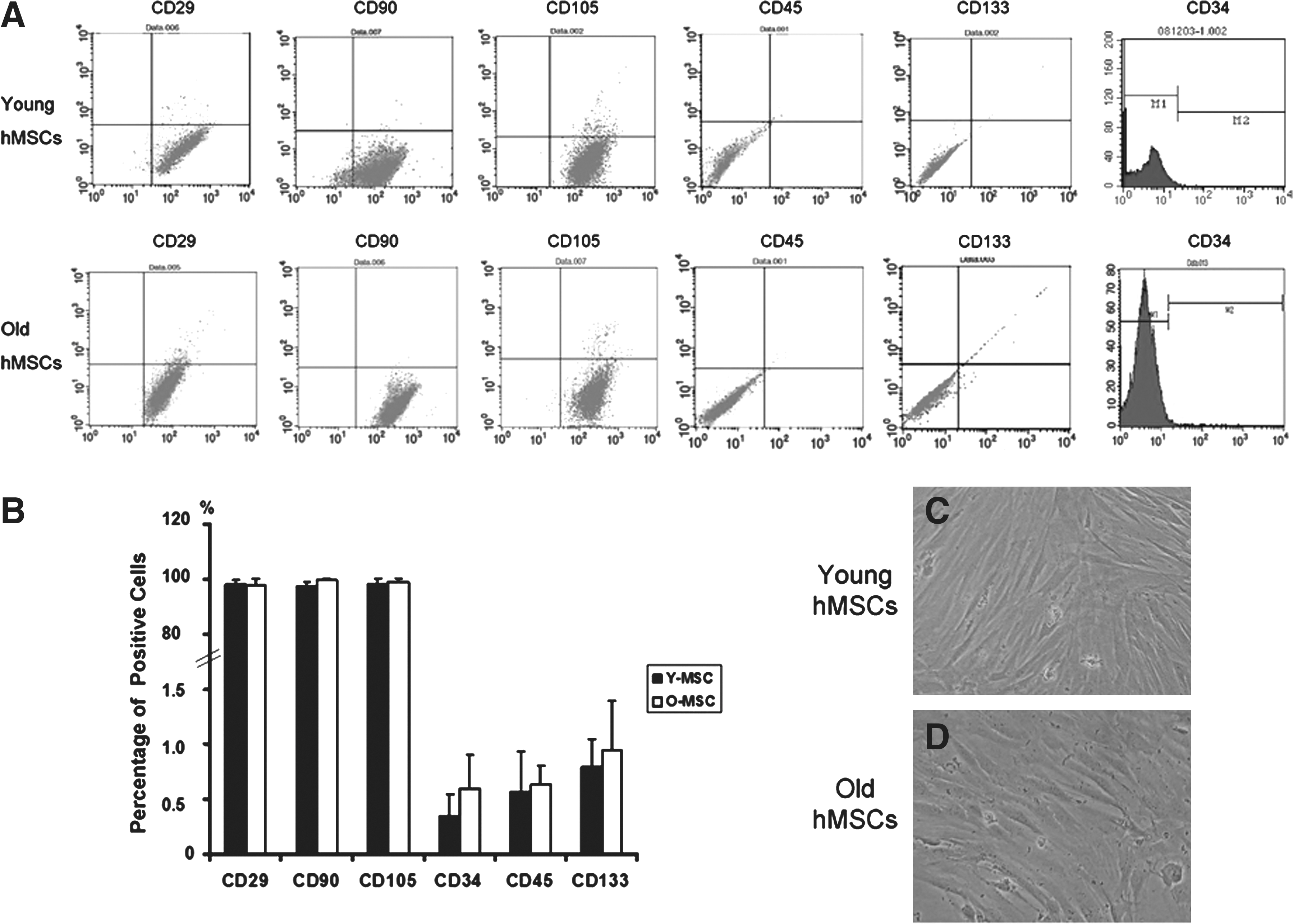
Characteristics of cultured human mesenchymal stem cells (hMSCs) were investigated using a flow cytometric analysis of surface antigen expression (
Although the cell composition was similar for cells derived from young and old donors, cell proliferation differed significantly between the two groups (Fig. 2). Cell cultures (n = 6/group) derived from young donors grew much faster than those from old donors (p < 0.001). We used a CFU-F assay to estimate the number of stem cells in each cell population (Fig. 3, n = 6/group). CFU-Fs were more numerous in cell populations derived from young donors (34.83 ± 5.74) rather than old donors (21.83 ± 3.97, p < 0.01).
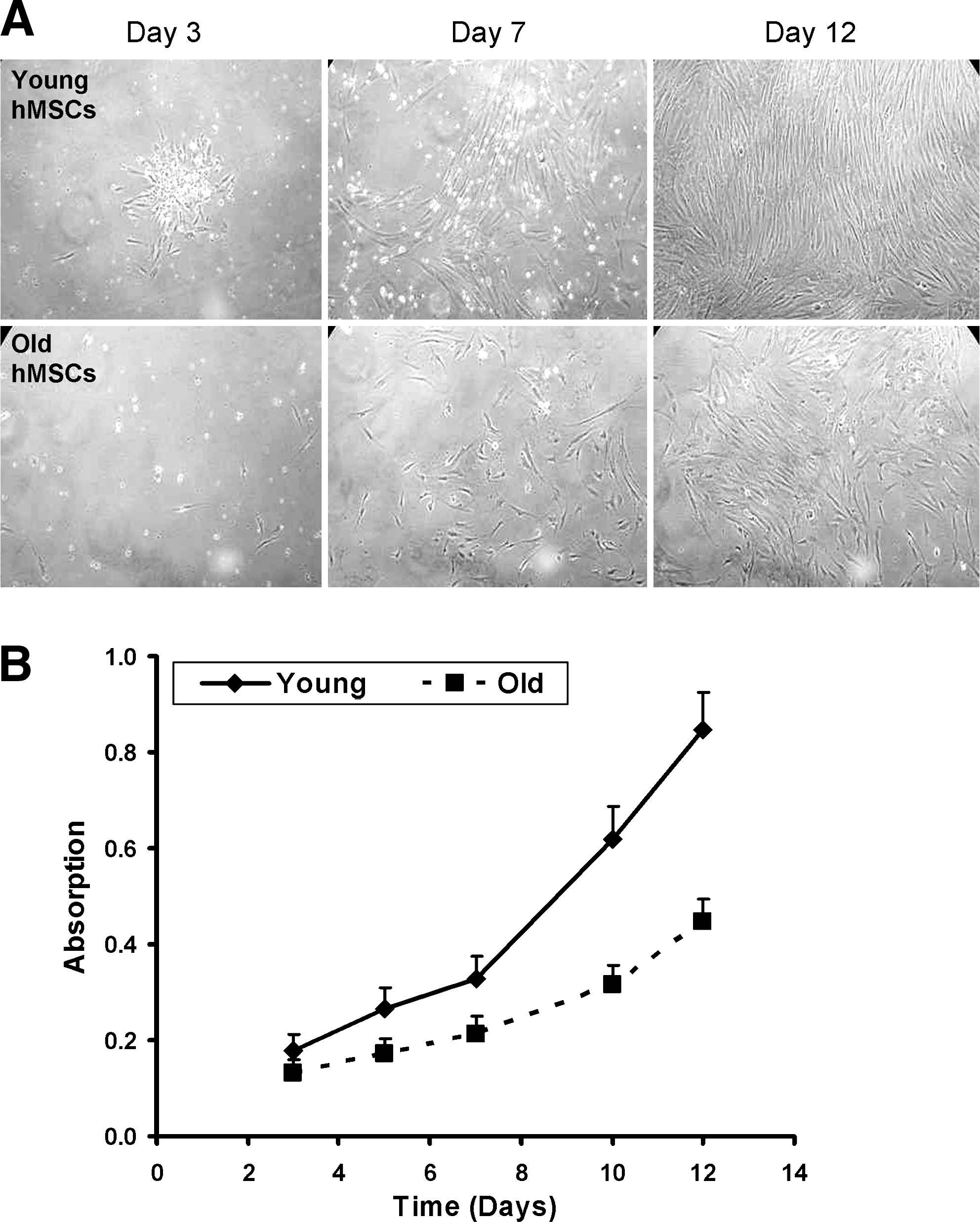
Representative micrographs of day 3, 7 and 12 cultures of human mesenchymal stem cells (hMSCs), isolated from young or old donors (young or old, respectively) from passage 3 (
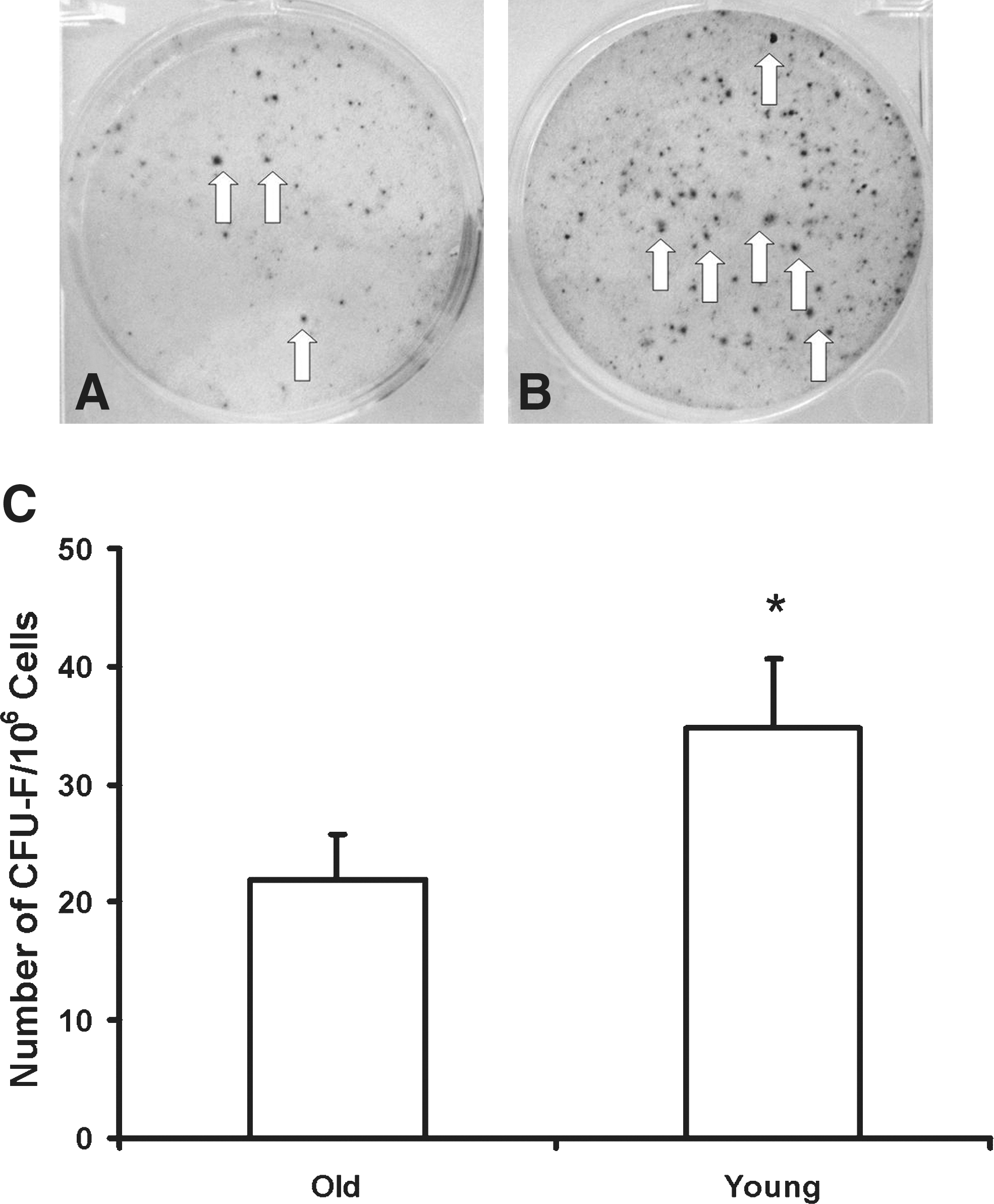
Culture dishes with colonies (visualized with Wright–Giemsa stain, indicated with arrows) produced by human mesenchymal stem cells (hMSCs) isolated from old (
Effect of donor cell age on cardiac function
To evaluate the regenerative capacity of the hMSCs from young and old donors, equal numbers of the cells were injected into the infarcted myocardium 15 min after an MI. Cardiac function was similar among the three groups prior to cell implantation (by echocardiography, Fig. 4, n = 6/group). At 1, 2, and 4 weeks after injection, ejection fraction and fractional shortening were higher in the animals that received young hMSCs than in those receiving old cells or medium (p < 0.05 by a time-varying ANOVA). Recipients of old cells and medium controls did not differ in cardiac function. At these times, left ventricular end-systolic and end-diastolic volumes were significantly lower (p < 0.01 by a time-varying ANOVA) after the implantation of young hMSCs compared to old hMSCs or medium.
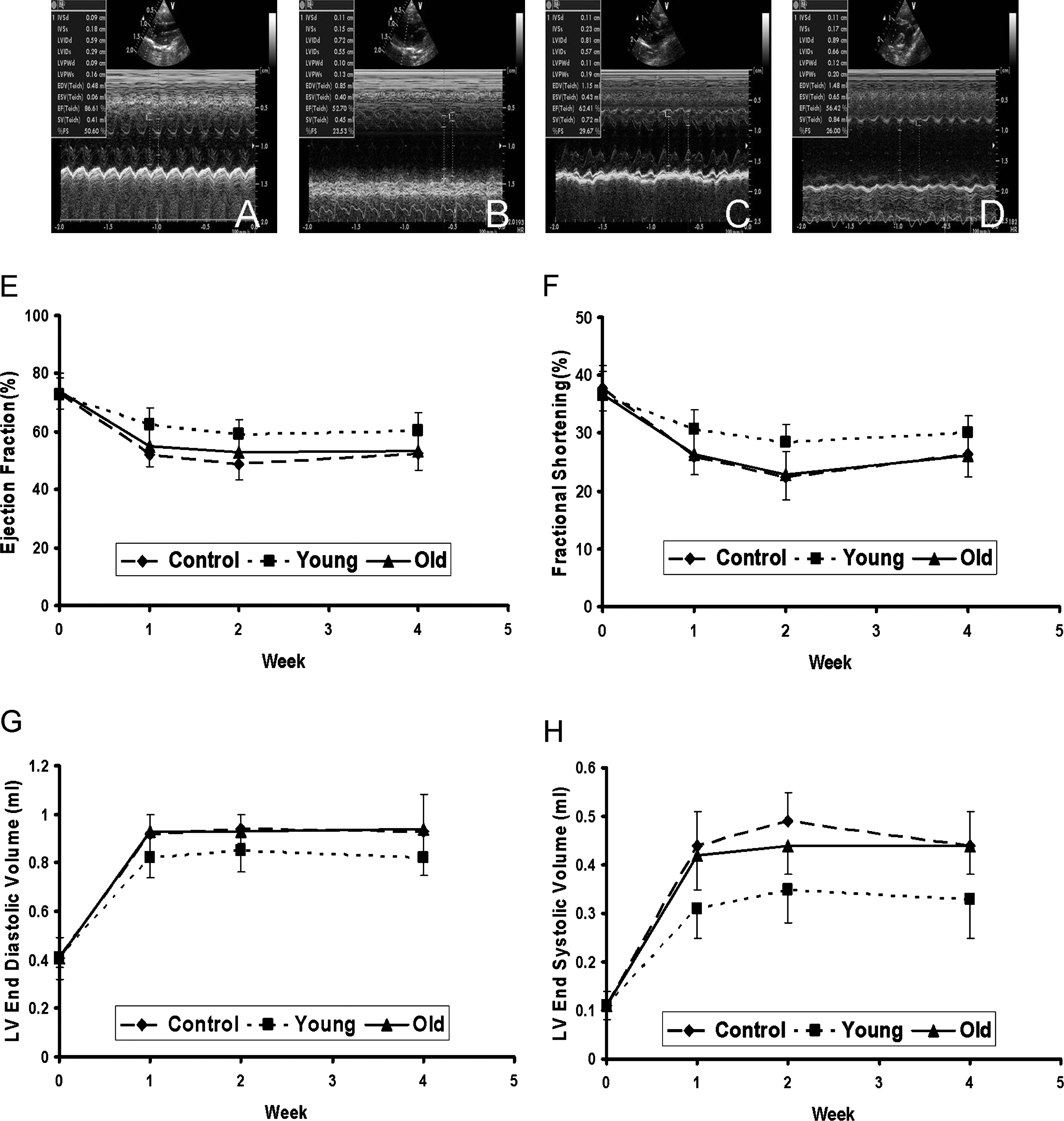
Echocardiographic images of rat hearts before myocardial infarction (MI) (
Possible mechanisms for the enhanced regenerative capacity of young hMSCs
At 4 weeks after injection, we identified more implanted cells within the infarcted myocardium of animals that received young (vs. old) donor cells (Fig. 5). However, there were only a few surviving cells in some of the animals that received old hMSCs. These results suggest that young donor cells have a greater post-implantation survival capacity than old cells.
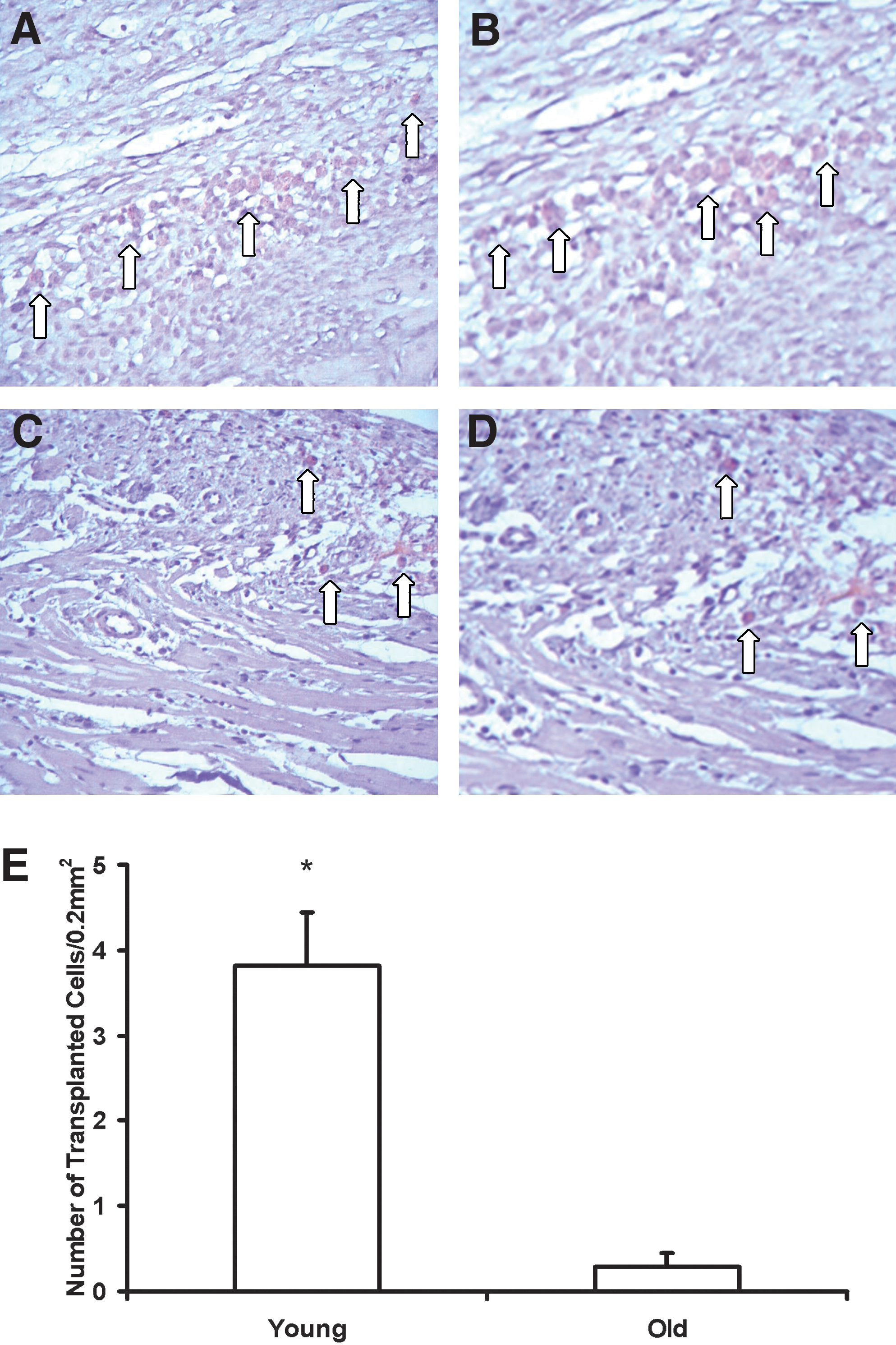
At 4 weeks after myocardial infarction (MI) and cell injection, representative micrographs of myocardial sections from animals that received human mesenchymal stem cells (hMSCs) isolated from young (
Consistent with the echocardiographic evaluation, morphological analysis showed that ventricular diameters were smaller in the animals receiving young (vs. old) cells (data not shown). These results suggest that the implanted cells altered matrix modulation and prevented ventricular dilation. Using the RT-PCR technique, we quantified the balance between MMP and TIMP gene expression. We found that rats receiving young hMSCs had significantly greater TIMP-3 expression and less MMP-9 expression than those receiving old cells or medium (Fig. 6A,B, n = 6/group, p < 0.05 for all groups). Meanwhile, animals that received old hMSCs expressed more TIMP-3 and less MMP-9 than controls (p < 0.05).
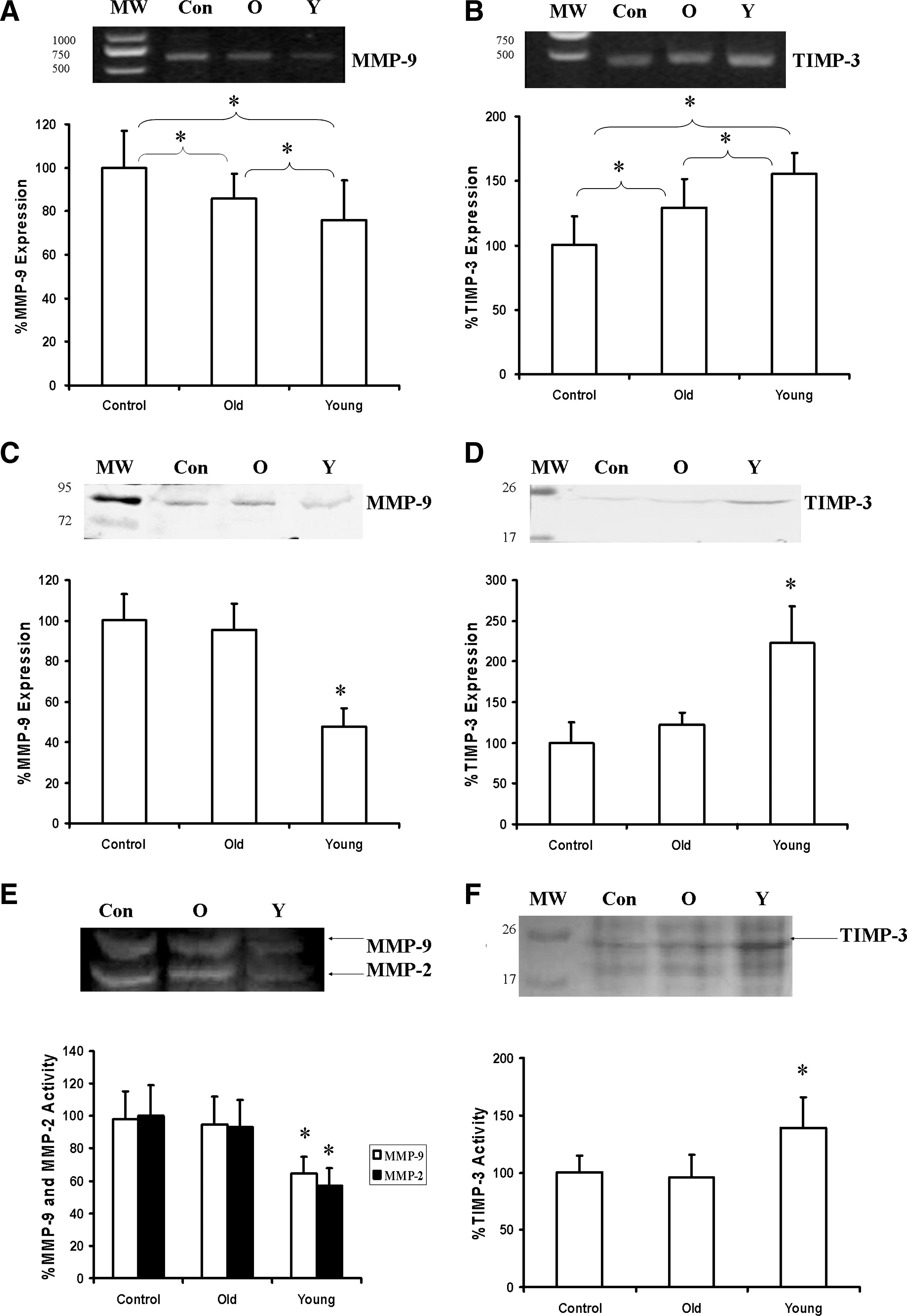
Gene expression (
In agreement with the gene expression data, protein levels of MMP-9 and TIMP-3 also differed among groups (Fig. 6C,D). Specifically, MMP-9 protein levels were significantly reduced in animals that received young hMSCs, whereas TIMP-3 protein levels were significantly increased by a factor of 1.5 (n = 6/group, p < 0.05) compared to those that received old hMSCs or medium. In contrast to the gene expression data, MMP-9 and TIMP-3 protein levels were similar in old cell recipients and controls. Zymography and reverse zymography were used to assess the activities of MMPs and their inhibitor. Compared to old cell and medium recipients, young hMSC recipients exhibited significantly lower MMP-9 and MMP-2 activities (p < 0.05) (Fig. 6E) and significantly higher TIMP-3 activity (p < 0.05) (Fig. 6F).
Collagen accounted for 48.7 ± 6.6%, 46.1 ± 6.6%, and 32.5 ± 7.7% of the infarct area in the medium, old hMSC, and young hMSC recipients, respectively, indicating a significant reduction in the percentage of collagen in the infarct border zone after the implantation of young hMSCs (vs. old hMSCs or medium) (p < 0.001).
Effect of donor cell age on angiogenesis induced by implanted cells
VEGF gene and protein expression were evaluated in all three groups using an RT-PCR assay and western blotting, respectively. We found that VEGF gene and protein levels were significantly elevated in the myocardium after young hMSCs (vs. old hMSCs or medium) were implanted (Fig. 7A,B). Histological study showed that young hMSCs also had the greatest vascular density (Fig. 7C–F, n = 6 per group, p < 0.05 vs. old hMSCs and medium). Vascular density did not differ between the recipients of old hMSCs and medium.
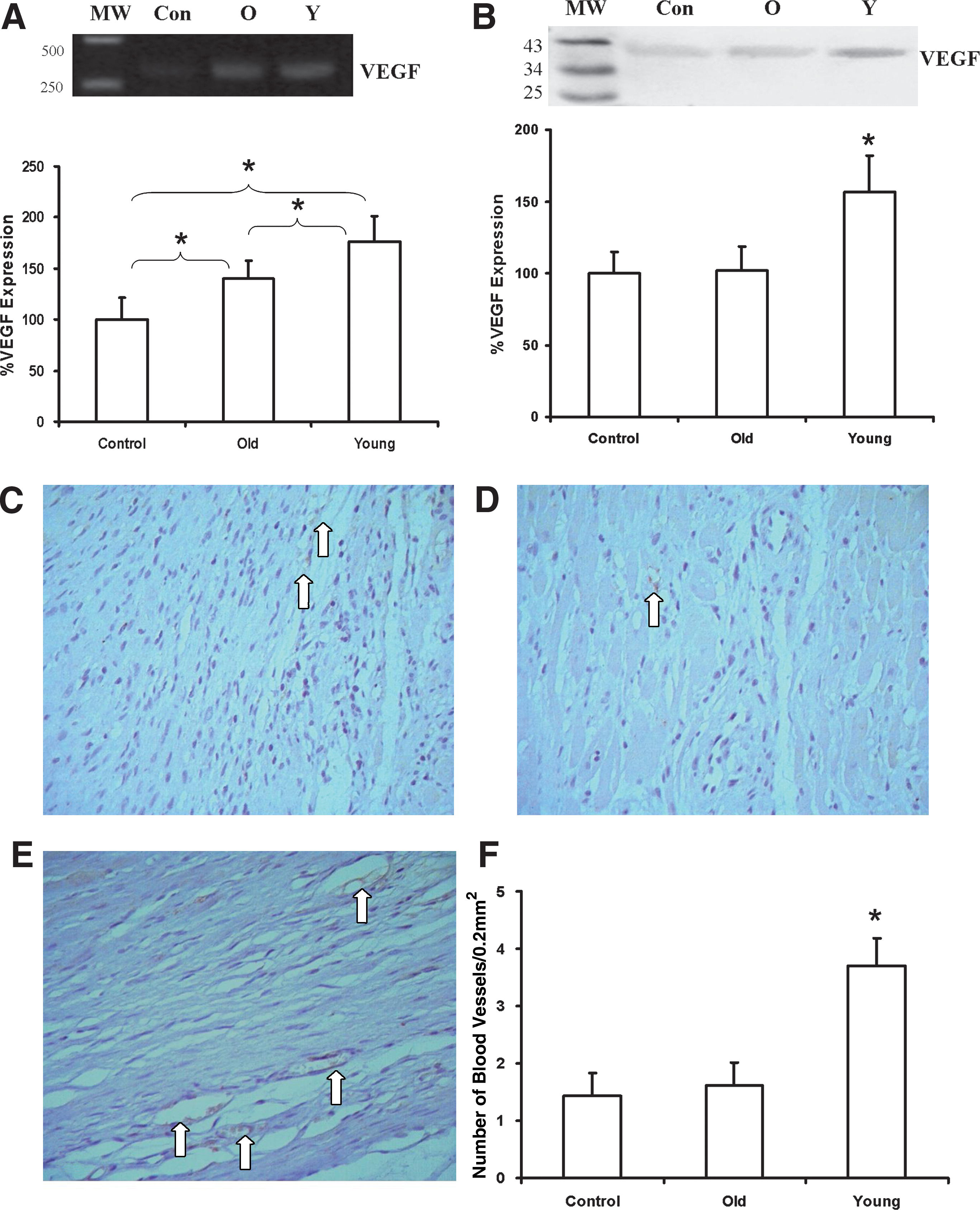
Densitometric analysis of vascular endothelial growth factor (VEGF) RNA levels (by reverse transcriptase polymerase chain reaction [RT-PCR],
Discussion
Although cardiac stem cells may induce cardiomyocyte regeneration after an MI, 23,24 bone marrow-derived MSCs have received extensive evaluation because of their efficacy in preclinical studies, their ready availability, and their ease of expansion and storage prior to clinical use. 5 The implantation of allogeneic or autologous MSCs has been demonstrated to improve damaged heart function in animals. 25,26 However, post-MI cell therapy with autologous hMSCs was less effective in clinical trials than in the preclinical animal experiments. 27 Reviewing studies performed using animals and those using patients, we found that the age of the recipient—and thus, the cell donor—was the main difference between the two types of studies. 11 Bone marrow stem cells from aging patients are known to lack the angiogenic potency of cells from young patients, 7 but this study is among the first to demonstrate that hMSCs from old patients lacked the regenerative capacity of hMSCs from young patients in a post-MI cell therapy model. In our rodent model, age diminished the benefits of hMSC transplant therapy. Specifically, old cells were less effective than young cells in preventing ventricular remodeling or inducing new vessel formation, and the in vitro growth characteristics predicted the in vivo performance of the cells.
Using echocardiography, we found that implanting hMSCs from young donors significantly preserved systolic function and ventricular volumes, whereas hMSCs from old donors were associated with a limited reduction of left ventricular end-systolic volumes. Thus, young hMSCs are likely the best candidate cells for patients who require cellular therapy to prevent heart failure. The ability of hMSCs to prevent cardiac dilatation and failure was as affected by the aging process as that of stem cells examined in previous animal experiments, which may explain the fact that autologous stem cell implantation was ineffective in the initial clinical trials.
The exact mechanism by which hMSCs improve myocardial function remains unknown; however, possible explanations include induction of angiogenesis to improve the blood supply to the infarct border zone, paracrine modulation of myocardial fibrosis and remodeling, and transdifferentiation of stem cells into smooth muscle cells or cardiomyocytes. 27,28 Myogenic transdifferentiation of the implanted cells seems unlikely in this case, considering the low engraftment rate and the fact that double immunostaining for human mitochondria protein and cardiac-specific myosin heavy chain revealed that none of the hMSCs expressed the cardiac protein at 4 weeks after implantation. However, we identified more blood vessels in the hearts of animals that received young hMSCs (vs. old hMSCs or medium), and so the functional effect of implanting young hMSCs might be principally due to improved myocardial perfusion. The increased vascular density may have resulted from the VEGF upregulation demonstrated with both RT- PCR and western blot. However, angiogenesis happens mainly at the infarct border zone, and the blood supply to the infarct area plays a very small role in myocardial regeneration without cell transplantation. 27
Diminished myocardial fibrosis and remodeling may also have improved cardiac function in the animals that received young hMSCs. An imbalance between TIMP-3 and MMP-9 has been shown to contribute to progressive ventricular dysfunction in human and experimental heart failure. 29,30 Here, we found that TIMP-3 levels (both RNA and protein) were higher in the hearts of animals implanted with young hMSCs (vs. old hMSCs and medium), and MMP-9 levels were lower (by RT-PCR and western blot).
Finally, differences in the characteristics of stem cells derived from patients of different ages can affect the outcome of hMSC therapy. 16 In this study, cell growth and clonogenic potential were both diminished in hMSCs from old donors. In addition, more young than old hMSCs survived at 4 weeks after implantation into the hostile environment of the myocardial infarct. There are a number of factors related to cell senescence that can limit the survival and regenerative potential of aging cells. 31,32 For example, telomere attrition or dysfunction increases cells' sensitivity to apoptotic stimuli 33 and induces environmental changes that restrict stem cell function and engraftment potential. 34 In future, a microarray analysis could be used to determine exactly which genes are differentially regulated by old and young hMSCs.
The present data indicate that young hMSCs are superior to stem cells derived from old patients as donor cells for cardiac cell therapy. It would be interesting to test whether the outcome with old hMSCs could be improved after correcting for the difference in clonogenic potential by identifying colony-forming cells in young and old populations prior to implantation. Of course, the cell recipients in this study were young animals, and it is possible that the functional effects associated with the young hMSCs reflected a strong host response to the implanted cells or specific interactions between the implanted cells and the host cardiomyocytes and extracellular matrix elements. 6,7 Our future studies will examine the success of post MI hMSC therapy in recipients of different ages.
In summary, we found that hMSCs derived from old patients were less robust than those from young donors. Further, old hMSCs exhibited significantly less plasticity than young hMSCs and conferred weaker functional benefits after implantation into an experimental infarct. The effects of age on stem cell function in vitro predicted the effects of age on the regenerative capacity of implanted hMSCs within the infarcted myocardium. In future, allogeneic cells or stem cell rejuvenation may be required to prevent congestive heart failure in patients who have suffered an extensive MI.
Footnotes
Acknowledgments
This research was supported by the Heart and Stroke Foundation of Ontario (T6604 to R.K.L.) and a Heilongjiang Provincial Award (GB06C40304). R.K.L. is a Longjiang Scholar and holds a Canada Research Chair in cardiac regeneration. We thank Heather McDonald Kinkaid for editorial assistance.
Author Disclosure Statement
No competing financial interests exist.