
Abstract
Age-related changes in immunity have been shown to highly influence morbidity and mortality. The aim of the present work was to study the effects of environmental enrichment (EE) (8–16 weeks) on several functions and oxidative stress parameters of peritoneal leukocytes, previously described as health and longevity markers, in mice at different ages, namely adult (44 ± 4 weeks), old (69 ± 4 weeks), and very old (92 ± 4 weeks). Mortality rates were monitored in control and enriched animals, and effects on survival of long-term exposure to EE until natural death were determined. The results showed that exposure to EE was efficient in improving the function (i.e., macrophage chemotaxis and phagocytosis, lymphocyte chemotaxis and proliferation, natural killer cell activity, interleukin-2 and tumor necrosis factor-α levels) and decreasing the oxidative–inflammatory stress (i.e., lowered oxidized glutathione content, xanthine oxidase activity, expression of Toll-like receptors 2 and 4 on CD4 and CD8 cells, and increased reduced glutathione and glutathione peroxidase and catalase activities) of immune cells. These positive effects of EE were especially remarkable in animals at older ages. Importantly, long-term exposure to EE from adult age and until natural death stands out as a useful strategy to extend longevity. Thus, the present work confirms the importance of maintaining active mental and/or physical activity aiming to improve quality of life in terms of immunity, and demonstrates that this active life must be initiated at early stages of the aging process and preserved until death to improve life span.
Introduction
Another regulatory system that suffers severe age-related alterations is the immune system. Indeed, dysfunction in immunity along with the aging process exerts a great influence on age-related morbidity and mortality, 5 and thus several age-related changes in immune functions have been correlated with increased mortality, such as low lymphoproliferative response to mitogens and natural killer (NK) cytotoxicity. 6 –9 Moreover, the recently proposed oxidative–inflammatory theory of aging suggests a key role for the immune system in accelerating the aging rate of the organism by persistent oxidative–inflammatory stress affecting leukocytes, leading to damage of these and neighboring cells and tissues. 8 Among studies on the immune system in aging, of special interest are those using noninvasive sampling, which allows the subsequent follow up of individuals under study. Peritoneal leukocyte suspension represents an easy access to this kind of sample in mice. Moreover, peritoneal leukocytes of aged mice are impaired in several of their functions and show increases in a variety of oxidative–inflammatory stress parameters, which have also been related to the increased morbidity and mortality of aged subjects. 8,9
The nervous and immune systems coordinate their activities to preserve homeostasis and health. 10 Given these close interactions between both regulatory systems, factors influencing the nervous system, eventually lead to immune modulation. In this regard, our research group has shown that mice exhibiting excess reactivity to stress and chronic anxiety suffer accelerated immune senescence and die prematurely. 9,11 –13 Conversely, some authors have described improved NK activity, together with lower levels of anxiety, in male mice exposed to 6 weeks of EE. 14 Moreover, our group has confirmed that this improvement of emotional responses by short-term EE exposure (5–8 weeks) is more marked in animals at older ages. 15 In this context, we wondered whether the improvement of age-associated deterioration in the nervous system by EE could result in positive effects for immune system aging too.
Thus, this is the first study in which the effects of EE (8–16 weeks) on several immune function and leukocyte oxidative stress parameters, previously described as health and longevity markers in both mice and humans, 6 –9 are analyzed in subjects at different stages of the aging process, namely adult, old, and very old. In addition, the long-term exposure to the EE until the natural death of the animals was monitored and effects on survival were determined.
Materials and Methods
Animals
We used 56 female ICR/CD-1 mice (Mus musculus) purchased from Harlan Ibérica (Barcelona, Spain) of different ages at the time of the study, namely adult (44 ± 4 weeks, n = 28), old (69 ± 4 weeks, n = 14), and very old (92 ± 4 weeks, n = 14). The mice were specifically pathogen free as tested by Harlan and according to the Federation of European Laboratory Science Associations recommendations. Mice were housed at 7 per cage and maintained at a constant temperature (22 ± 2°C) in sterile conditions inside an aseptic air negative-pressure environmental cabinet (Flufrance, Cachan, France), on a 12/12 h reversed light/dark cycle (lights on at 20:00 h). Mice had access to tap water and standard Sander Mus pellets (A04 diet from Panlab L.S., Barcelona, Spain) ad libitum. Diet was in accordance with the recommendations of the American Institute of Nutrition for laboratory animals. Mice were treated according to the guidelines of the European Community Council Directives (86/6091 EEC) as well as to the Spanish laws regarding protection of animals (1201/2005 RD).
Environmental enrichment
At the beginning of the experiment, mice were randomly distributed in two groups: control (C) and EE. Half of the animals from each age group were assigned to each experimental condition. EE consisted of two different objects included in the cages, which were changed every 2 days at 08:00 h. The rodent toys used were: orange bucket, jolly ball, hoop, holed ball, yellow tunnel, rough red object (Lillico, Panlab), yellow billard ball, and silver ball (El Corte Ingles, Madrid, Spain). A red kennel, which was cleansed once a week, was maintained permanently inside the cages. Animals from the control groups were only handled at the time of object changing. The EE animals had been exposed to 8–16 weeks of EE when the immune studies were performed (sampling and experimental procedures went on for 8 weeks). In addition, EE was maintained until the natural death of the animals, and mortality was monitored. Individuals that died prematurely and/or because of technical reasons were not considered for the longevity study.
Collection of peritoneal leukocytes
Peritoneal suspensions were collected between 08:00 and 10:00 h, to minimize circadian changes in the immune system, without sacrificing the animals. Each mouse was held by the cervical skin, the abdomen was cleansed with 70% ethanol, and 3 mL of sterile Hanks solution, previously tempered at 37°C, was injected intraperitoneally. After abdominal massage, approximately 80% of the injected volume was recovered. The following studies were performed using unfractionated peritoneal leukocytes to better reproduce the in vivo immune response and oxidative status. Macrophages and lymphocytes from the peritoneal suspensions were identified by their morphology and quantified in Neubauer chambers using optical microscopy (40×). Additionally, aliquots of the cellular suspensions were used to confirm the leukocyte counts by flow cytometry, as described below. Cellular viability was checked by the Trypan Blue (Sigma, St. Louis, MO) exclusion test, and only suspensions with cell viability of 99 ± 1% were used for the experiments. The peritoneal suspensions were adjusted to a specific number of macrophages, lymphocytes, or total leukocytes, depending on the parameter analyzed as described in the corresponding section.
Membrane expression of leukocyte differentiation antigens (CD) and Toll-like receptors
Percentages of CD11b (leukocyte differentiation antigen typical of macrophages), CD11c (dendritic cells), CD4 (T helper lymphocytes), CD8 (T cytotoxic lymphocytes), and CD19 (B lymphocytes) cells present in the samples were analyzed, as well as their membrane expression of Toll-like receptor-2 (TLR-2) and TLR-4, by flow cytometry. Monoclonal antibodies were used for superficial staining at the following final concentrations: Fluorescein isothiocyanate (FITC)-conjugated anti-CD11b (CALTAG Laboratories, Burlingame, USA), 1/100 (vol/vol); FITC-conjugated anti-CD11c (BD Pharmingen, San Diego, USA), 1/100 (vol/vol); phycoerythrin (PE)-cyanine (Cy5)-conjugated anti-CD4 (BD Pharmingen), 1 μL/106 cells; FITC-conjugated anti-CD8 (BD Pharmingen), 1/100 (vol/vol); FITC-conjugated anti-CD19 (BD Pharmingen), 2/100 (vol/vol); PE-conjugated anti-TLR2 and anti-TLR4 (Bioscience, San Diego, CA), 1 μg/106 cells. Briefly, aliquots of 3 × 105 leukocytes/mL were centrifuged at 560 × g for 10 min, supernatants were discarded, and 30 μL of single antibodies (CD11b, CD11c, CD19) or 30 μL of antibody mixtures (TLR2/CD11b, TLR4/CD11b, TLR2/CD11c, TLR4/CD11c, CD4/CD8, TLR2/CD4/CD8, TLR4/CD4/CD8, TLR2/CD19, TLR4/CD19) were added to each tube. A total of 30 μL of phosphate-buffered saline (PBS) and bovine serum albumin (BSA) was added to blank tubes. Cells were incubated in the presence of the antibodies for 30 min at 4°C in the dark. Then, tubes were washed twice at 560 × g for 5 min in PBS-BSA to remove free antibodies. Cells were acquired on the flow cytometer (FACSCalibur Flow Cytometer, Becton Dickinson, Franklin Lakes, NJ) immediately after staining. Cells were gated according to their forward- and side-scattering, and only the high expression of the corresponding leukocyte differentiation antigen or TLR was taken as positive (FL-H >101 for CD11b, CD11c, CD8, CD19, TLR-2 and TLR-4; FL-H >102 for CD4). Results were analyzed with Cell Quest Pro software (BD Biosciences, San Jose, CA) and expressed as percentages of CD11b, CD11c, CD4, CD8, and CD19 cells with regard to the total number of cells present in the samples, and percentages of cells positive for TLR-2 or TLR-4 regarding the total number of cells in each leukocyte population.
Macrophage and lymphocyte chemotaxis
The chemotactic index (CI) was evaluated by the capacity of macrophages and lymphocytes to migrate through a filter, with a pore diameter of 3 μm (Millipore, Madrid, Spain), included in a Boyden chamber 16 and towards a chemoattractant agent (f-Met-Leu-Phe [fMLP], Escherichia coli, Sigma; 10−8 M), following a method previously described. 17 After a 3-h incubation of aliquots of 300 μL of the peritoneal suspension adjusted to 5 × 105 macrophages/mL or 5 × 105 lymphocytes/mL in Hanks medium, filters were fixed and stained, and the CI was calculated by counting, in an optical microscope (100×), the total number of macrophages or lymphocytes, respectively, in one-third of the lower face of the filters.
Macrophage phagocytosis
Phagocytosis of inert particles (latex beads, Sigma; diluted to 1% in PBS) was carried out following a method previously described. 17 Aliquots of 200 μL of the peritoneal suspension adjusted to 5 × 105 macrophages/mL Hanks medium were incubated in migratory inhibitory factor (MIF) plates (Kartell, Noviglio, Italy) for 30 min. The adhered monolayer was washed and resuspended in 200 μL of Hanks medium plus 20 μL of latex solution. After 30 min of incubation, plates were washed, fixed, and stained, and the phagocytosis was expressed as the number of latex beads ingested per 100 macrophages (phagocytic index [PI]), which was assessed by optical microscopy (100×). The number of ingesting macrophages per 100 macrophages phagocytic efficiency, (PE) was also determined.
Reactive oxygen species
Reactive oxygen species (ROS) levels were measured by fluorometric assay following a method previously described 17 with some modifications, using the probe de-acetylated 2′,7′-dichlorodihydrofluorescein (H2DCF-DA; Molecular Probes, Paisley, UK) which is oxidized in the cytoplasm by ROS into 2′,7′-dichlorofluorescein (DCF), a highly fluorescent compound. Aliquots of 200 μL of the peritoneal suspension adjusted to 106 leukocytes/mL in Hanks medium were incubated for 30 min, and 20 μL of H2DCF-DA (1 mM/well) were added to each well. After 15 min of incubation, 20 μL of phorbol myristate acetate (PMA; 5 ng/mL in well; Sigma) and 20 μL of Hanks medium were added to stimulated and nonstimulated samples, respectively. Samples were incubated for 15 min, and fluorescence was recorded with excitation and emission wavelengths of 485 nm and 535 nm. The results were expressed as percentage of stimulation with respect to the basal value without PMA.
Lymphoproliferation
A previously described method 18 was used with minor modifications. The peritoneal suspension was adjusted to 5 × 105 lymphocytes/mL in complete medium containing RPMI-1640 (PAA, Pasching, Austria), 10% fetal bovine serum (Gibco), and 1% gentamicin (PAA), and 200 μL of the suspension were cultured in 96-well plates with concanavalin A (ConA; 1 μg/mL per well; Sigma) or lipopolysaccharide (LPS; Escherichia coli, 055:B5, 1 μg/mL per well; Sigma) as stimulated samples or in complete medium alone as nonstimulated samples. After 48 h of incubation, 5 μL [3H]thymidine (MP Biomedicals) were added to each well (2.5 μCi/well), 100 μL of culture supernatants were collected for cytokine measurements, and medium was renewed. Cells were harvested 24 h thereafter. The results were expressed as counts per minute (cpm).
Cytotoxicity
Murine lymphoma YAC-1 cells were used as targets in the NK activity assay. 18 An enzymatic colorimetric test was used for cytolysis measurements of target cells (Cytotox 96 TM, Promega, Madison, WI) based on lactate dehydrogenase determination using tetrazolium salts. Aliquots of 100 μL of peritoneal leukocytes, previously adjusted to 106 leukocytes/mL in RPMI-1640 without Phenol Red (PAA) were cultured at an effector/target rate of 10/1 and incubated for 4 h. Thereafter, lactate dehydrogenase enzymatic activity was measured in 50 μL/well of the supernatants by addition of the enzyme substrate and absorbance recording spectrophotometrically at 490 nm. The results were expressed as percentage of lysis of target cells.
Cytokines in response to stimuli
ConA-stimulated interleukin-2 (IL-2) and LPS-stimulated tumor necrosis factor-α (TNF-α) were measured by enzyme-linked immunosorbent assay (ELISA; R&D Systems, Minneapolis, MI, and HyCult Biotechnology, Uden, The Netherlands, respectively) in supernatants of peritoneal leukocytes cultured as described above. The results were expressed as pg/mL. Sensitivity for IL-2 and TNF-α was <0.3 and <10 pg/mL, respectively. The range of detection was 15.6–1000 pg/mL for IL-2 and 8–2000 pg/mL for TNF-α.
Total and oxidized glutathione
Total glutathione (GSH), the main nonenzymatic reducing agent of the organism, as well as its oxidized form (GSSG), were assayed by the enzymatic recycling method previously described 19,20 by monitoring the change in absorbance at 412 nm. Briefly, aliquots of 1 mL of the peritoneal suspension (106 cells/mL Hanks medium) were centrifuged at 1200 × g for 10 min at 4°C. Pelleted cells were resuspended in previously degassed media, containing 5% of trichloroacetic acid in 0.01 N HCl for GSH measurements and 50 mM phosphated buffer (pH 7.0) plus ethylene-diaminetetraacetic acid (EDTA, 1 mM; Sigma) and N-ethylmaleimide (NEM, 12.5 mM; Sigma) to block reduced GSH in tubes for measuring GSSG. Samples were then sonicated and centrifuged at 3200 × g for 5 min at 4°C. Aliquots of supernatants of immune cells were measured using the following reagents in the reaction mixture: 5,5′-dithiobis(2-nitrobenzoic acid) (DTNB, 6 mM; Sigma), β-nicotinamide adenine dinucleotide phosphate, reduced form (β-NADPH, 0.3 mM; Sigma), and glutathione reductase (GR, 10 U/mL; Sigma). The reaction was monitored for 240 s or 140 s for GSH or GSSG determinations, respectively, and the results were expressed as nmol/106 cells.
Xanthine oxidase
Xanthine oxidase (XO) activity was determined by fluorescence (Amplex Red Xanthine/Xanthine Oxidase Assay Kit, Molecular Probes) in 50 μL of the peritoneal suspension adjusted to 106 leukocytes/mL Hanks medium. In the assay, XO catalyzes the oxidation of purine bases (xanthine) to uric acid and superoxide. This superoxide spontaneously degrades in the reaction mixture to hydrogen peroxide (H2O2), which in the presence of horseradish peroxidase (HRP), reacts stoichiometrically with Amplex Red reagent to generate the red fluorescent oxidation product resorufin. Resorufin has fluorescence emission maxima of 585 nm and a high extinction coefficient (54,000 cm−1M−1). After 30 min of incubation, measurement of fluorescence was performed in a microplate reader using excitation at 530 nm and emission detection at 595 nm. Levels as low as 0.1 mU/mL of XO could be detected. The protein content of the same samples was evaluated following Bicinchoninic Acid (BCA) Protein Assay Kit protocol (Sigma). The results were expressed as international milliunits (mU) of enzymatic activity per mg of protein.
Antioxidant enzymes
The peritoneal suspension was adjusted to 106 leukocytes/mL in Hanks medium, and aliquots of 1 mL were used to perform the enzymatic assays. The cells were centrifuged at 1200 × g for 10 min at 4°C, and the pellets were resuspended in 50 mM phospate buffer (pH 7.0) previously degassed. Thereafter, the samples were sonicated and centrifuged at 3200 × g for 20 min at 4°C. Glutathione peroxidase (GPx) enzymatic activity, which plays a fundamental role in GSH cycle, was measured using the original technique described by Lawrence and Burk 21 with some modifications. 20 This method is based on the oxidation of GSH in the presence of cumene hydroperoxide (0.71 mM in phospate buffer; Sigma) by GPx. A reaction solution was used, containing GSH (4 mM; Sigma), GR (1 U/mL; Sigma), β-NADPH (0.2 mM; Sigma), EDTA (1 mM; Sigma), and sodium azide (4 mM; Sigma). The reaction was followed spectrophotometrically for 300 s by the decrease of the absorbance at 340 nm due to NADPH oxidation. The results were expressed as mU of enzymatic activity per 106 leukocytes. Catalase (CAT) activity was determined following the method described by Beers and Sizer, 22 with slight modifications previously introduced by us. 20 The enzymatic assay was followed spectrophotometrically for 80 s at 240 nm through the decomposition of H2O2 (14 mM in phospate buffer) into H2O + O2. The results were expressed as International Units (IU) of enzymatic activity per 106 leukocytes.
Statistical analysis
The results were expressed as mean ± standard error (SE). The normality of the samples and the homogeneity of variances were checked by the Kolmogorov–Smirnov and Levene analyses, respectively. Differences in immune parameters were studied through two-way analysis of variance (ANOVA) followed by one-way ANOVA. The Tukey test with a level of significance set at p < 0.05 was used for post hoc comparisons when variances were homogeneous, whereas its counterpart analysis Games–Howell set at the same significance level was used in case of unequal variances. Differences in life span were studied through the Kaplan–Meier test, with a minimum significance level (log rank, Mantel–Cox) also set at p < 0.05.
Results
Immunity
No remarkable changes were found with respect to the percentages of the different peritoneal leukocyte populations studied in enriched and nonenriched animals (Table 1). However, enriched subjects showed overall a decreased fraction of CD4 cells as compared to nonenriched controls (p < 0.05). Additionally, very old mice showed an overall increased percentage of CD11b cells with respect to old (control and enriched) subjects (p = 0.052). Interestingly, even if not statistically significant, old controls showed a decreasing trend in the percentage of CD11b cells as compared to the adults, which was increased in enriched animals. In contrast, very old controls showed an increasing trend in the relative amount of these cells when compared to the adults, which was decreased in their enriched age-matched counterparts (Table 1).
Results are expressed by mean ± standard error of 5–10 values corresponding to that number of subjects (5–10 adults; 5–7 old; 4–7 very old), each value being the mean of duplicate assays.
p < 0.05 with respect to the value in nonenriched animals.
p = 0.052 with respect to the value in old (control and enriched) animals.
C, Control; EE, environmentally enriched.
Regarding macrophage functions, the decrease in the chemotactic capacity of these cells (Fig. 1A) shown by very old animals (p < 0.001) was not observed in the very old enriched group, which showed similar values to the adults and higher than old (p < 0.05) and very old controls (p < 0.001). Enriched adults were found to present higher macrophage chemotactic capacity than age-matched controls (p < 0.001). Moreover, phagocytosis (Fig. 1B,C), a key function of macrophages, was reduced in both old (PI, p < 0.01; PE, p < 0.05) and very old (PI and PE, p < 0.001) controls as compared to adults, but not in the enriched groups. Thus, old (PI, p < 0.01) and very old (PI and PE, p < 0.001) enriched animals showed increased macrophage phagocytosis as compared to age-matched controls, and very old enriched subjects in fact higher than old controls (PI, p < 0.01). Overall, stimulation of ROS levels (Fig. 1D) was found to be higher in enriched than in control animals (p < 0.05).
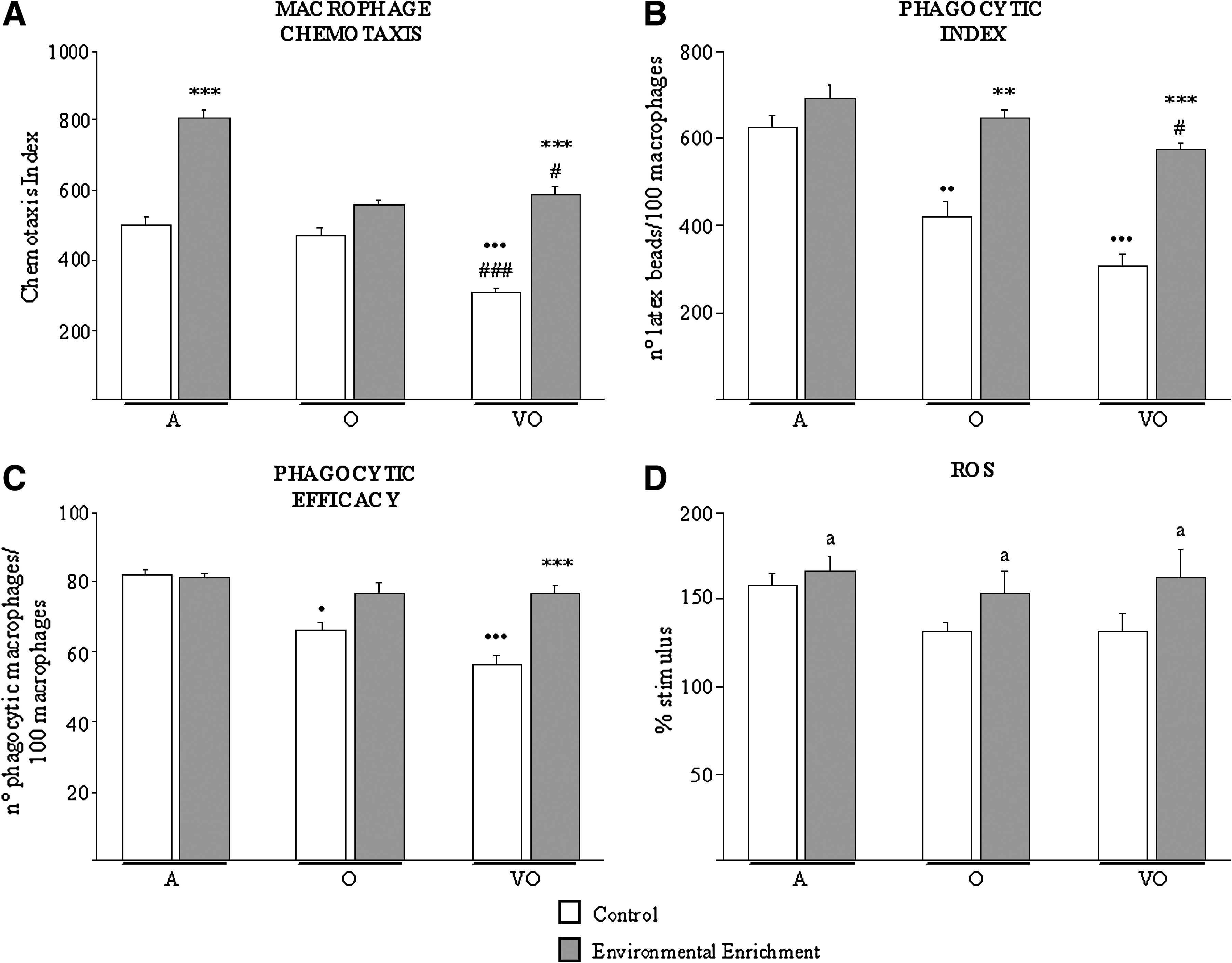
Macrophage functions and reactive oxygen species (ROS) levels. Macrophage chemotaxis index (CI, number of macrophages) (
One of the most important functions of lymphocytes crucial for their role in immune responses is their proliferative capacity (Fig. 2A). Basal lymphocyte proliferation appeared to be increased in enriched adults as compared to their age-matched counterparts (p < 0.05). Proliferation stimulated by the T cell mitogen ConA, which declined in old controls when compared to the adults (p < 0.001), was more preserved in old enriched animals (p < 0.05 vs. adult controls, p < 0.01 vs. old controls). In addition, very old enriched subjects showed higher function values than their age-matched controls (p < 0.001). Proliferation in response to LPS, mimicking bacterial infection, declined in lymphocytes from both older groups when compared to adults (p < 0.001). This loss of proliferative capacity was not observed in the very old enriched group, which showed higher function values than all three control groups (p < 0.001). Enriched adults were also found to have increased LPS-stimulated proliferation than control adults (p < 0.001). However, a slight functional decline was found in old enriched animals in comparison with their age-matched controls (p < 0.01).

Lymphocyte functions and cytotoxicity. Resting, concanavalin A (Con A)- and lipopolysaccharide (LPS)- stimulated lymphoproliferation (counts per minute) (
The lysis of YAC-1 tumor cells (FIg. 2B), which was lower in very old animals as compared to adults (p < 0.05), was preserved in the enriched group (p < 0.01 vs. very old controls). Moreover, the loss of chemotactic capacity of lymphocytes (Fig. 2C) shown by very old animals (p < 0.001 vs. adult controls) was not observed in the very old enriched group (p < 0.05 vs. very old controls). Additionally old enriched subjects showed higher functional values than their age-matched counterparts (p < 0.05 vs. old controls), although lower than adults (p < 0.001).
EE completely prevented the decline of ConA-stimulated IL-2 levels measured in culture supernatants of peritoneal leukocytes from very old individuals as compared to the adults (Fig. 3A, p < 0.001). Thus, levels of this cytokine in very old enriched animals were higher than in age-matched and younger old controls (p < 0.001). Regarding TNF-α levels in culture supernatants of peritoneal leukocytes in response to LPS (Fig. 3B), the enriched environment was efficient in preventing the decrease shown by old subjects when compared to the adults (p < 0.001). TNF-α levels were significantly higher in old enriched animals than in their age-matched counterparts (p < 0.01).

Cytokine levels in response to stimuli. Concanavalin A (Con A)-induced interleukin-2 (IL-2) (
TLR-2 and TLR-4 leukocyte membrane expression
Immune cells recognize pathogens via TLRs, the basic signaling receptors of innate immunity. TLR-4 is essential for effective development of the immune response against LPS from Gram-negative bacteria, and, together with TLR-2, represents the main receptor for the recognition of a great variety of components from bacteria cell walls. Moreover, TLRs have also been found to be expressed on cells from acquired immunity 23 and recognize diverse endogenous antigens, thus playing a role in age-related diseases such as Alzheimer's disease and atherosclerosis. 24 –26 Interestingly, most relevant effects of EE on TLR leukocyte membrane expression (Fig. 4) were shown by the prevention of age-related increase of TLR-2 on CD4 (p < 0.05 vs. adult and old controls) and CD8 (p < 0.05 vs. old controls) cells and of TLR-4 on CD4 cells (p < 0.01 vs. adult, p < 0.05 vs. old controls), from very old animals (Fig. 4[II]A,B). Thus, enriched very old subjects showed lower expression of TLRs on these cells than age-matched controls (p < 0.05 and p < 0.01 for TLR-2 on CD4 and CD8 cells, respectively; p < 0.001 for TLR-4 on CD4), similarly to younger ages. Other remarkable effects of EE were shown on CD11c cells (Fig. 4[I]B), reducing the expression of TLR-2 and TLR-4 in very old subjects as compared to age-matched controls (p < 0.01 and p < 0.05, respectively), whereas these showed higher values than younger old controls (p < 0.05 for TLR-2).
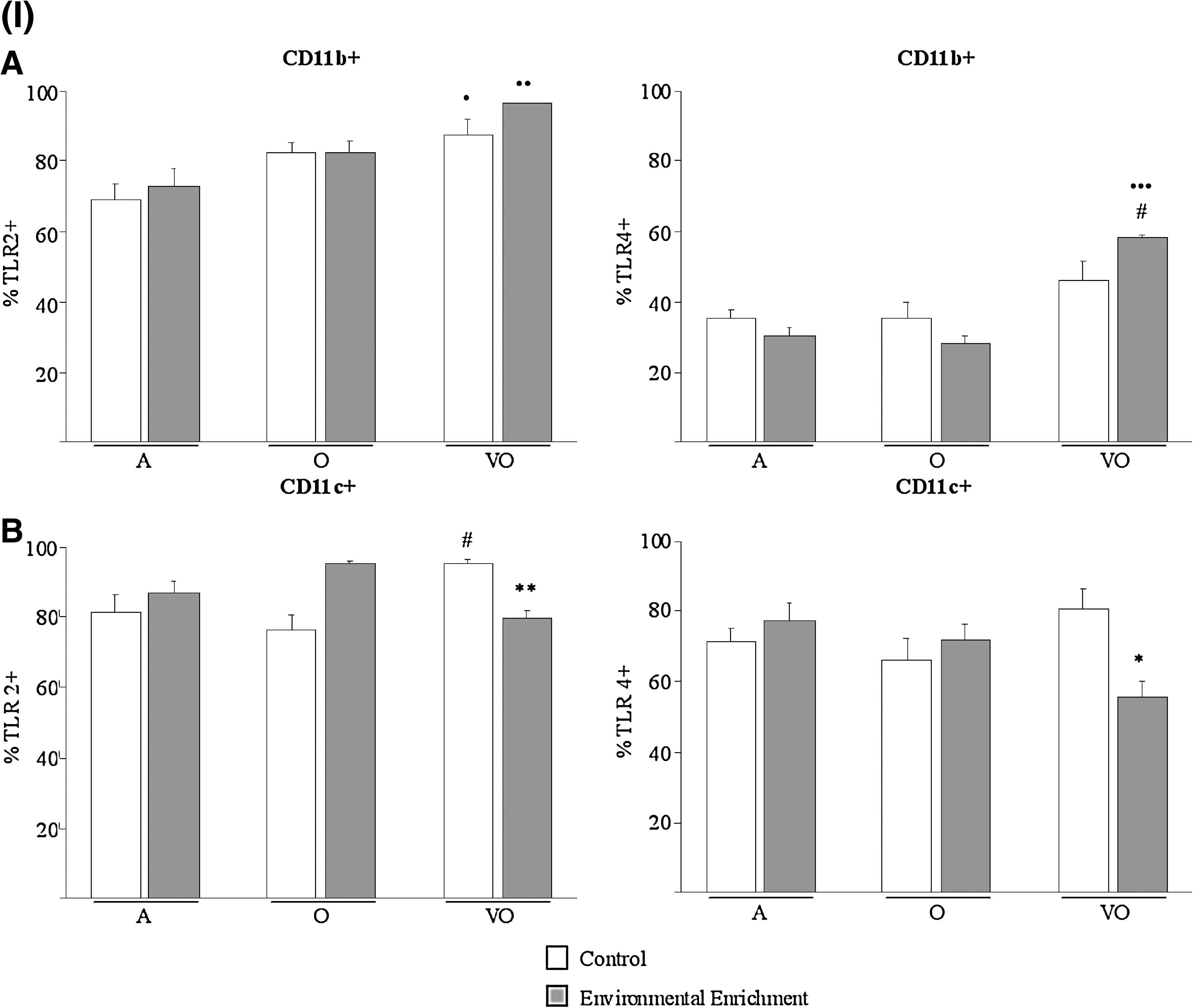
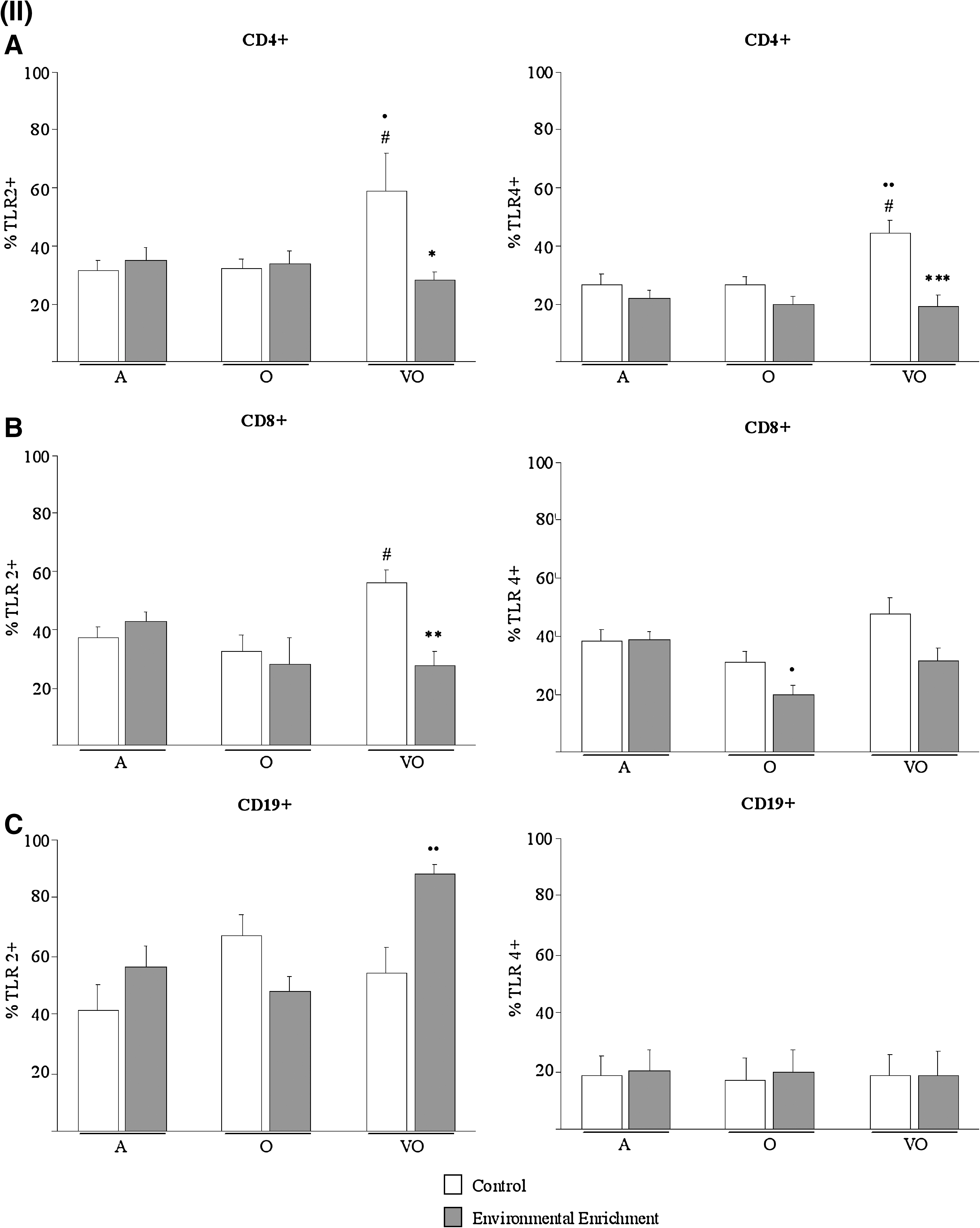
Leukocyte oxidative status
In regard to oxidant markers in leukocytes, EE completely prevented the increase in GSSG levels (Fig. 5A) and XO activity (Fig. 5B) shown by leukocytes from old animals compared to adults (p < 0.05 and p < 0.01, respectively). Thus, values of both oxidants were lower in enriched than in nonenriched old subjects (p < 0.05 and p < 0.001, respectively). In addition, enriched adults showed lower levels of GSSG than controls (Fig. 5A, p < 0.001).
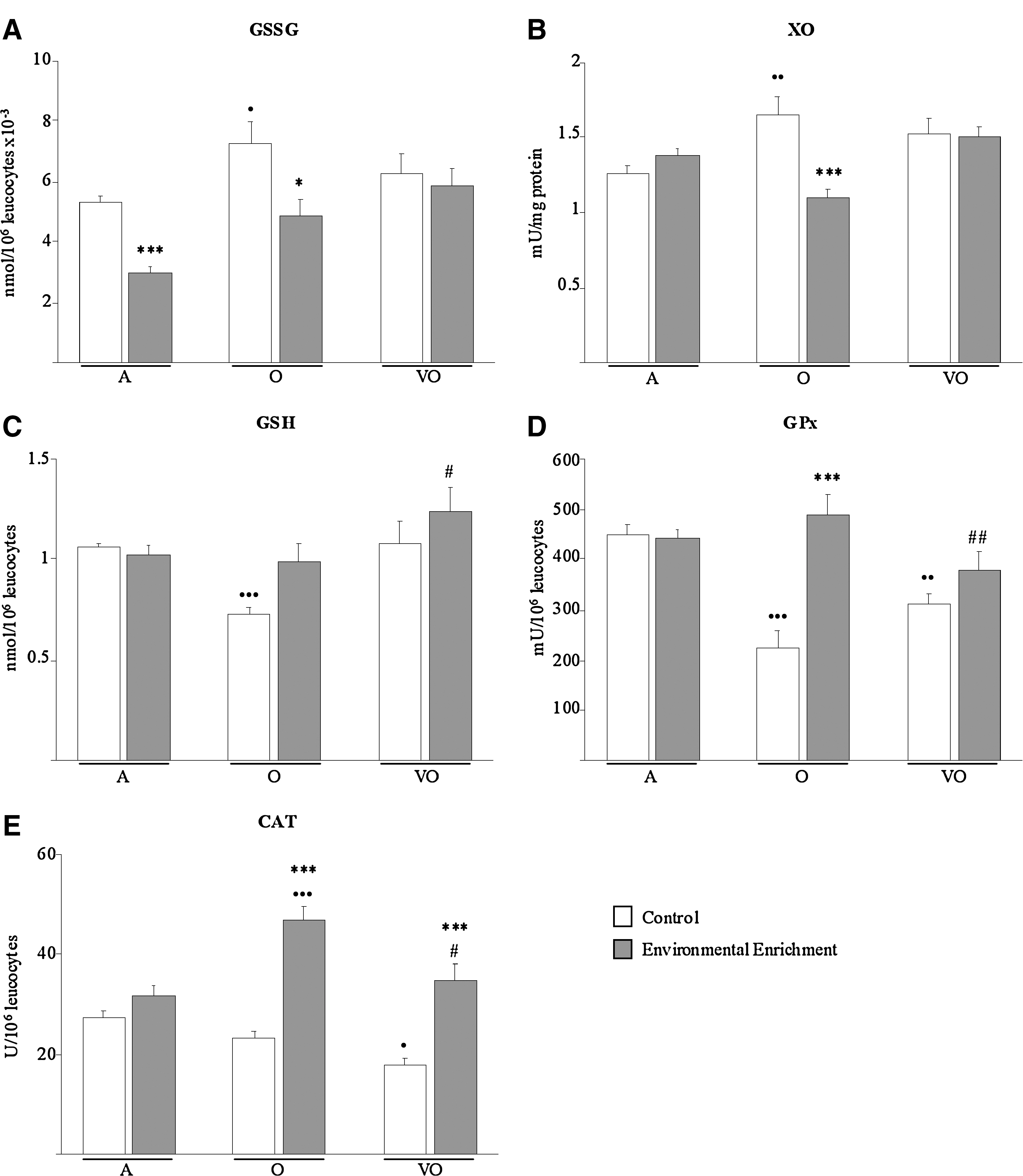
Oxidative status markers. Oxidants are oxidized glutathione levels (GSSG, nmol/106 leukocytes × 10−3) (
Conversely, the EE was efficient in preventing the decline in GSH content (Fig. 5C) and GPx activity (Fig. 5D) observed in leukocytes from both old (p < 0.001) and very old (p < 0.01 for GPx) animals when compared to adults. GPx activity was significantly higher in leukocytes from enriched than nonenriched old subjects (p < 0.001), and both parameters were raised in enriched very old animals over the values shown by old controls (p < 0.05 for GSH, p < 0.01 for GPx). Moreover, CAT activity (Fig. 5E), which protects against severe oxidative stress, was reduced in peritoneal leukocytes from very old mice (p < 0.05) in comparison to adults, but not in the enriched very old group, which showed similar values to the adults and higher than age-matched and younger old controls (p < 0.001 and p < 0.05, respectively). This positive effect of EE on the CAT activity of leukocytes was especially remarkable in the old group, which had higher values than age-matched and adult controls (p < 0.001).
Longevity
As shown in Fig. 6, long-term exposure to EE from adulthood and until the natural death of the animals was an efficient strategy in improving the life span of the subjects (p < 0.01).
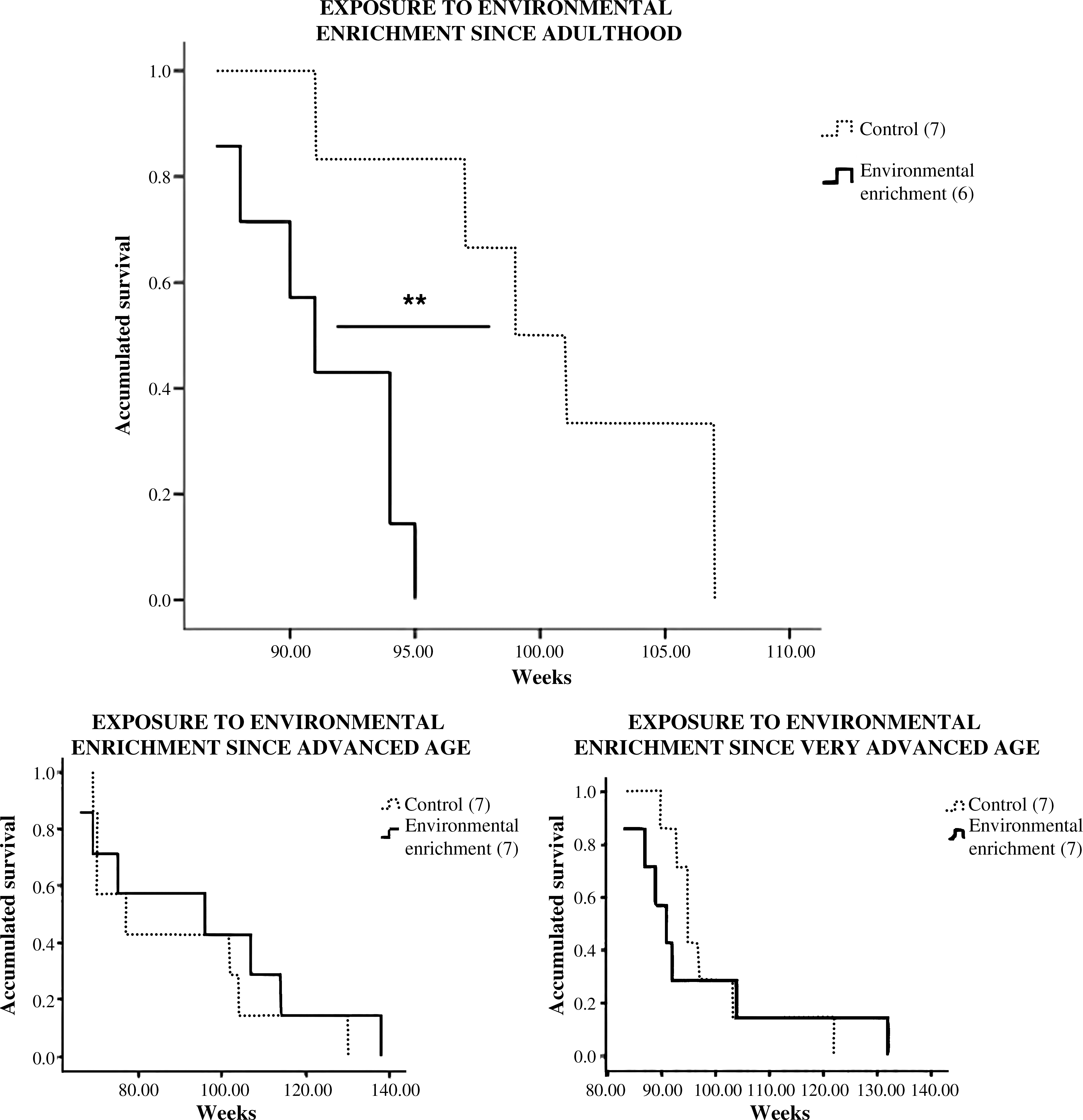
Mortality records from control ICR/CD1 female mice and from animals that had been exposed to environmental enrichment since adulthood, advanced, or very advanced age, and until their natural death. (**) p < 0.01 with respect to the value in the nonenriched control group of the same age.
Discussion
This is the first study describing the effects of EE on a variety of immune functions and leukocyte oxidative stress parameters, previously described as health and longevity biomarkers in mouse and humans, 6 –9 in subjects at different stages of the aging process, namely adult, old and very old. The results showed that 8–16 weeks of exposure to EE is successful for improving the function and decreasing the oxidative stress of immune cells. These positive effects of EE are especially remarkable in animals at older ages. Importantly, long-term exposure to EE from adult age and until natural death stands out as a useful strategy to improve longevity.
Age-related decline in phagocyte functions, such as chemotaxis to the infection focus or phagocytosis of foreign particles reported in the present and previous works, 9,12,27 is not shown by animals that had been exposed to EE, which at older ages preserves adult-like functional values. These positive effects of EE on innate immunity could result in lowered susceptibility and vulnerability to bacterial and viral infections during old age, which stand out as the most common causes of disease and death with aging. 8,28,29 Interestingly, regarding the CD11b cell fraction in peritoneal suspensions, the EE seems to exert a modulatory action that counteracts the effects of aging. However, this result should be confirmed in future studies using larger samples to improve the statistical significance of the present work. Moreover, the lower levels of TNF-α in response to bacterial LPS with aging shown in the present report and in previous data 30 are also improved by the EE, which thus could promote the inflammatory response against infection. Importantly, some authors have described that human subjects who at age 85 years produce low levels of LPS-induced TNF-α have a more than two-fold elevated overall mortality risk compared to their peers with higher production. 31
In addition, ROS levels represent the most important mechanism of microorganism destruction by immune cells, although ROS in excess have been described to be deleterious for leukocytes and neighboring cells. 32 Future studies are needed to check for the possible beneficial effects on leukocyte bactericidal activity of the increased ROS levels shown by enriched animals. These are unlikely to be harmful though, because their increase was moderate and in addition the results showed higher antioxidant capacity in leukocytes from enriched animals, which would make possible the fast neutralization of ROS.
Regarding lymphocyte functional abilities, age-related impairment of proliferative responses to the T cell stimulator ConA and decline of the T cell growth factor IL-2 shown in the present and previous work 11,33 are reversed in animals exposed to EE. Importantly, a preserved lymphoproliferative response to ConA constitutes a key immunological biomarker of the no immune risk phenotype in humans, leading to health and survival. 7 Preservation of proliferative abilities in healthy aging has also been related to a maintained CD4/CD8 ratio. 7 Nevertheless, CD4 cell fraction appeared to be decreased in peritoneal leukocytes of animals that had been exposed to EE. Other authors have described lowered percentages of CD4 and/or CD8 cells in splenocytes of enriched young male mice as compared to nonenriched controls. 34,35 The physiological significance of this result deserves future research. However, because the enriched animals of the present work were in good health and their immune cell responses were in general more adequate than in age-matched nonenriched controls, the decreased percentage of CD4 cells in enriched subjects does not seem to be detrimental but instead could show a mechanism aiming to achieve good homeostasis. Moreover, impairment of the lymphocyte migration to the infection focus in aged subjects, reported in the present and previous data, 8,36 is reversed in enriched animals. Taken together, the age-related decline in cellular immunity, which highly influences morbidity and mortality in the elderly, 6,7 is efficiently improved by the EE.
In addition, the decrease of antitumoral NK activity throughout the aging process shown in the present and previous reports 8,37 is reversed in enriched animals to the values shown by adult controls. Because NK cell activity is considered one of the most sensitive screening markers of neuroimmunoendocrine system imbalance, 38,39 the preservation of a better response in enriched animals could be reflecting recovery of the neuroimmunoendocrine communication, leading to preserved homeostasis and good health. In fact, improved NK cell activity together with lower anxiety levels have previously been described in male mice exposed to 6 weeks of EE. 14 Thus, the EE could decrease the age-related increase in viral infections and cancer. 40
Interestingly, TLRs and persistent activation of leukocytes such as T cells have been proposed to play a key role in a variety of age-related chronic inflammatory processes. 24 –26,41,42 However, EE completely reversed the increase in TLR-2 and TLR-4 on CD4 and CD8 cells shown by animals at late stages of the aging process, which could contribute to lower development of age-related diseases and decreased morbidity in the enriched animals. A similar result was observed regarding CD11c cells.
We propose that this improvement of the age-related decline in immune responses in enriched animals could be mediated by reduction of the oxidative stress affecting leukocytes. Indeed, the present work shows that relevant antioxidant defenses, such as GSH levels and GPx, and CAT which decline during the aging process according to the present and previous reports, 18,43 –45 are significantly promoted in both groups of enriched older animals to values similar or even above those shown by younger controls. Recovery of the intracellular GSH pool and promotion of its recycling in enriched animals could be the basis of the improvement in a variety of immune responses such as IL-2–dependent functions, including T cell proliferation and NK activity, which are especially sensitive to its depletion. 46 In addition, Fernández et al. have previously described that 8 weeks of EE increases brain GSH content in old rats, which the authors related to behavioral and cognitive improvements. 4
Because high catalase activity in immune cells has specifically been related to longevity in humans, 45 the result regarding this antioxidant enzyme, which is increased in older enriched subjects over the values of adult controls, is especially of interest. Moreover, increase in antioxidant defenses in leukocytes from enriched old subjects was coincident with decreased oxidant markers, in terms of lowered GSSG content and XO activity, which show age-related increases in the present and previous works. 18,47 Interestingly, GSSH also decreased in enriched adults when compared to age-matched controls, suggesting that animals at early stages of the aging process may benefit from several aspects of EE.
Thus, exposure to short-term (8–16 weeks) EE leads to a striking improvement of leukocyte functions, which is likely mediated through decreased oxidative stress affecting immune cells. These positive effects of the environment rich in intellectual and/or physical activities are especially relevant in animals at older ages, in which the age-related immune deterioration is more marked. Importantly, our group has confirmed in previous studies that the improvement of emotional responses after short-term (5–8 weeks) EE exposure reported by several authors 14 is also more marked in aged subjects. 15 Interaction with the EE provides the animals with a diversion from monotonous and thus stressful cage life, resulting in lower hypothalamic–pituitary–adrenal axis activity, in terms of adrenocorticotropic hormone and corticosterone concentrations, both in baseline conditions and after mild stress. 48 Research on stress in older adults has shown that chronic stress contributes to effects that mimic, exacerbate, and possibly accelerate the effects of aging on immunity. 49 Moreover stress-related emotional responses are exacerbated in aged subjects. 15 In this scenario, the improvement of the age-related immune changes exerted by EE reported in the present work could be based on the neuroendocrine improvements shown in previous works. 14,15,48 Correlation between both features should be further investigated in future studies.
In addition, our results indicate that when the EE starts at intermediate older ages, animals benefit overall, but they could take longer to adapt to the challenge of the EE. This is shown, for instance, regarding LPS-induced proliferation, which increases in enriched adult and very old animals compared to their respective age-matched controls, but decreases in enriched old subjects resorting to old controls. Moreover, the only strategy efficient in improving longevity was exposure to the EE from adulthood and prolonged until natural death. Importantly, longitudinal studies in human centenarians have shown lifelong preserved immunity as a marker of longevity, 7,45 and cross-sectional studies in naturally long-living mice have reproduced these results. 8,37 Furthermore, our research group has shown that mice exhibiting intrinsic excess reactivity to stress and chronic anxiety suffer accelerated immune senescence and die prematurely. 9,11 –13 Thus, lifelong, well-preserved immunity as well as emotional responses are likely to be the basis of this life-span increase by long-term EE exposure. Future longitudinal studies using a larger sample size that would allow adding a sampling point during the old and/or very old age in animals exposed to EE from adulthood are advisable to further confirm this hypothesis.
In conclusion, the present work confirms the importance of maintaining active mental and/or physical activity aiming to improve health and life quality in terms of immunity, and demonstrates that this active life must be initiated at early stages of the aging process and preserved until death to improve life span.
Footnotes
Acknowledgments
This work was supported by the Ministerio de Ciencia e Innovación (MCINN) (BFU2008-04336), Research Group of UCM (910379), and Red Temática de Investigación Cooperativa en Envejecimiento y Fragilidad (RETICEF) (RD06/0013/003).
Author Disclosure Statement
No competing financial interests exist.