
Abstract
Polyphenolic antioxidants from dietary sources are frequently a topic of interest due to widespread scientific agreement that they may help lower the incidence of certain cancers, cardiovascular and neurodegenerative diseases, and DNA damage and even may have antiaging properties. On the other hand, questions still remain as to whether some antioxidants could be potentially harmful to health, because an increase in glycation-mediated protein damage (carbonyl stress) has been reported in some cases. Nevertheless, the quest for healthy aging has led to the extensive use of phytochemically derived antioxidants to disrupt age-associated deterioration in physiological function and to prevent many age-related diseases. Although a diet rich in the polyphenolic forms of antioxidants does seem to offer hope in delaying the onset of age-related disorders, it is still too early to define their exact clinical benefit for treating age-related disease. This review critically examines polyphenolic antioxidants, such as flavonoids, curcumene, and resveratrol in health, disease, and aging with the hope that a better understanding of the many mechanisms involved with these diverse compounds may lead to better health and novel treatment approaches for age-related diseases.
Introduction
Oxidative stress is the basis for the free radical theory of aging, first proposed by Denham Harman 3 and widely believed to be the root cause of age-related protein damage and tissue deterioration in the body. Antioxidants react with ROS and RNS and thus alleviate cellular damage by terminating the otherwise harmful free radical chain reactions. In that regard, several mechanisms that include exogenous nutrients and endogenous antioxidant systems have evolved to protect from inevitable intracellular or extracellular oxidative stress. 4 The most widely known antioxidant nutrients are derived from phytochemicals (plant-derived chemicals), which include polyphenols in the form of flavonoids. The flavonoids are broadly classified into anthocyanidins (e.g., cyaidin, delphinidin, malvidin), flavanols (e.g., catechin, epicatechin), flavonols (e.g., quercetin, fisetin), and flavones (e.g., luteolin) (Fig. 1). Some of the nonphenolic antioxidants include carotenoids (e.g., β-carotene, lycopene), vitamin C (ascorbic acid), and vitamin E (tocopherols). These antioxidants have excellent free-radical quenching properties (Fig. 2). 5

Structures of representative flavonoids and rutin, a naturally occurring glycoside of quercetin.
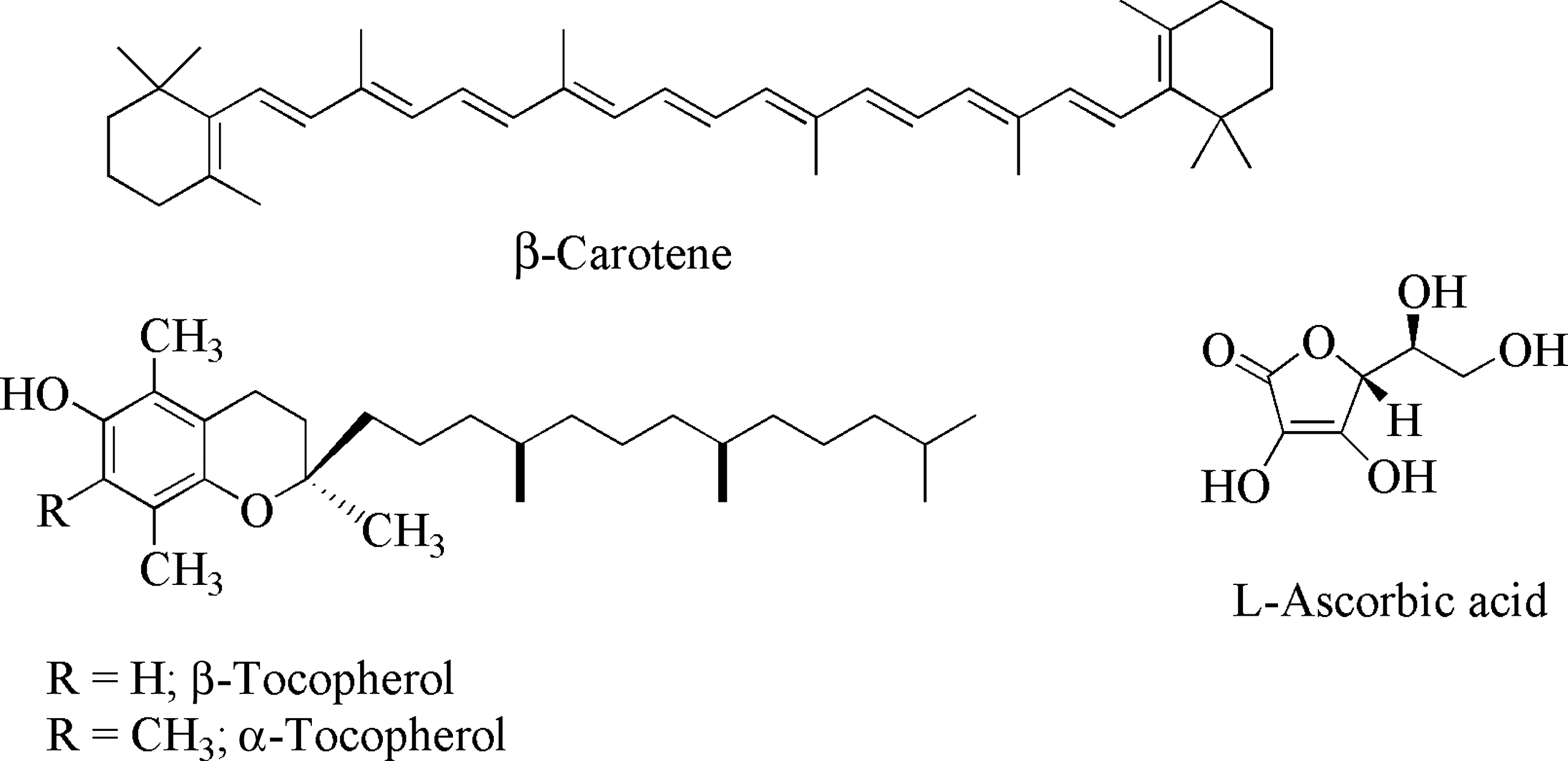
Structures of some representative nonpolyphenolic antioxidants: β-carotene, vitamin E (tocopherols), and vitamin C (
When the vast antioxidant literature is explored, mixed results are reported concerning the benefits of antioxidant supplementation for treating age-related diseases. A diet rich in antioxidants seems to offer hope in delaying the onset of many age-related disorders, such as atherosclerosis or cancer. However, large clinical trials that used only select antioxidants have not shown evidence for the benefit of antioxidants as therapeutic agents. 6 Conversely, clinical trials involving relatively smaller groups of patients have shown beneficial effects for select antioxidants, such as N-acetylcysteine, or when some antioxidants are used in combination with several antioxidant species. 6 Therefore, it is necessary to enroll ideal patients and use appropriate duration when undertaking this kind of clinical trial before we can make fair assessment for the therapeutic role of these antioxidants. The reported clinical data, however, are in agreement with the central premise that age-related progression of many diseases may be related to general cytokine stress (inflammation) and oxidative stress (the production of ROS and RNS), which can result in subsequent molecular damage and eventual cell death.
Antioxidant Nature of Flavonoids
Phenolic compounds are widely distributed in plant-based foods and have widely diverse structures, largely via hydroxylated aromatic rings. Flavonoids typically exist as their corresponding glycosides in plants, and, depending on structure and the oxidation state of the central pyran ring, are further subclassified. For example, rutin exists in plant sources as the quercetin-3-O-rhamnosyl glucoside. A variety of flavonoids, including flavonols (quercetin), flavanols (epicatechins and catechins), flavones (luteolin), flavanones, isoflavones (soy), and anthocyanidins (cyanidin), have been characterized in plant-based foods (Fig. 1).
In this review, we will focus on those polyphenolic compounds that are thermally stable, bioavailable, and are known to modulate health, disease, and aging. It is important to stress that many antioxidant compounds, in particular vitamins C, E, and some polyphenols, are known to possess antioxidant as well as prooxidant properties under certain conditions. The molecular mechanisms proposed for antioxidants are based largely on in vitro studies. In solution, many of these compounds are redox-reversible (Fig. 3), 7 which may explain their antioxidant as well as prooxidant properties, depending on the relative concentrations of the ROS/RNS. Nevertheless, some polyphenols possess this same lability in redox state in vivo, 7 which may explain why they possess some disease-fighting activity as well as protective properties, or conversely participate in glycation reactions. In that regard, the therapeutic use of antioxidants such as these can be considered “double-edged swords,” i.e., they have both reduction/oxidation potential and activity.
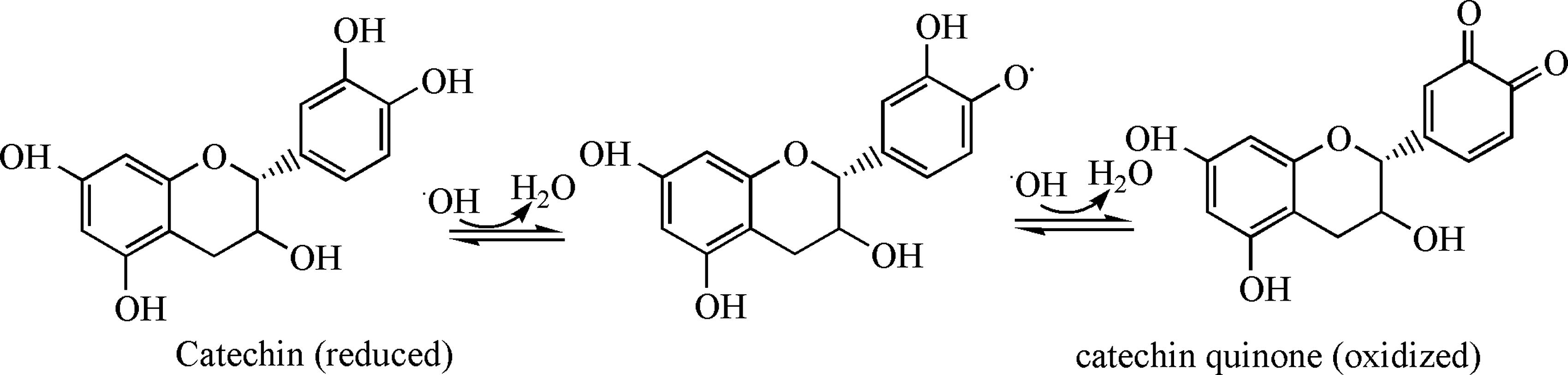
Redox forms of catechin in the presence of reactive oxygen species/reative nitrogen species (ROS/RNS).
However, it should be pointed out that not all phytochemicals or vitamins have the same mechanisms of action. For example, polyphenolic antioxidants and vitamins also can affect various molecular targets directly, such as activating receptors, which can be independent of their antioxidant activities. Furthermore, they can affect signal transduction pathways and have multiple downstream events, including, but not limited to, the modulation of cellular prooxidant/antioxidant balance. 8,9 Some polyphenolic compounds not only scavenge ROS but also have transition metal ion (e.g., Fe2+, Cu+) chelating properties, which substantially contribute to attenuation of oxidative stress. 10 Recently, it was suggested that vitamin C and E supplementation, i.e., nondietary antioxidant dosing, might preclude the health-promoting effects of exercise. 11 Data of this nature should be interpreted with caution for pragmatic reasons as well as those already elucidated. Moreover, while exercise can induce oxidative stress, it also ameliorates insulin resistance and boosts endogenous antioxidant defense capacity. Furthermore, there are many endogenous defenses to oxidative stress in vivo, and human plasma contains many proteins with intrinsic antioxidant properties, including proteins that are sulfur rich, such as glutathione peroxidase or transition metal-binding proteins and superoxide dismutase. 12,13
Although the human body has a complex endogenous antioxidant defense system that scavenges free radicals, cells may not function properly, die, or undergo apoptosis due to free radical-induced irreparable DNA damage, cellular membrane alterations, and damage. Many endogenous proteins and dietary nutrients can be cytoprotective, but it must be stressed that free radical damage is inevitable. Damage accumulates with advancing age, not only during many disease processes but in healthy individuals as well. Clearly, deficiencies in antioxidant vitamins, paucity of adequate antioxidant defense, and environmental stressors, such as ionizing radiation, all can lead to many disease states. 13
Polyphenols and the “French Paradox”
Polyphenols, such as those recently found in aged red wine, 14 have particular importance for diabetes, cancer, aging, and neurodegenerative diseases because of their effect on certain enzymes that posttranslationally modify the acetylation pattern of histone proteins. 15,16 France and Italy are two major wine-producing European countries said to demonstrate a paradoxical finding largely related to the diet in these countries. These same people are basically able to consume fat-rich and low-density lipid-saturated foods in greater quantities than most other groups, while at the same time remaining relatively unaffected in their cardiovascular health. 17 The French diet, in particular, is particularly lipid rich, and the French consume many fatty foods, yet cardiovascular pathologies are not prevalent. They frequently consume wine and have low morbidity and mortality through cardiovascular disease. This peculiarity has been commonly referred to as the “French Paradox.”
One explanation for the French Paradox can be found by considering the characteristic Mediterranean diet as a whole, which is generally rich in fruits, fresh high-fiber vegetables, and vitamins. 18,19 Of particular interest is the fact that red wine contains polyphenols (Fig. 1). The polyphenolic nature of any flavonoid compound is mainly responsible for its specific antioxidant properties, which is one of the main factors believed to be involved in protecting these people from heart attacks. 20 Flavonoids serve as electron donors to the appropriate free radical–derived oxidants based on their redox potentials. The reduction potentials of flavonoid radicals are lower than those of alkyl peroxyl radicals, and thus the flavonoids are effective chain-breaking antioxidants in oxidation processes mediated by peroxyl radicals. 21
Two main types of polyphenols exist in red wine, namely regular flavanols 22 and anthocyanidins 23 (Fig. 1). Anthocyanidines, the oxidized forms of flavanols have good metal chelation ability due through their vicinally located hydroxyl groups. Stilbene derivatives (e.g., trans-resveratrol, Fig. 4) also are found in red wine at up to 100 μM (Fig. 3). Resveratrol may have properties other than that as antioxidant. 24 Currently, it is the focus of intense research both for the cardiovascular system and the brain, but holds promise for aging as well. Although it is not the most abundant phenol in wine, trans-resveratrol has been recognized as the main agent responsible for the French Paradox. 25 However, polyphenols are not the only protective compounds in a vegetable-rich diet or in wine because other putative cardioprotective factors may involve folate as contributors to the French paradox, because folate affects homocysteine levels. 26,27
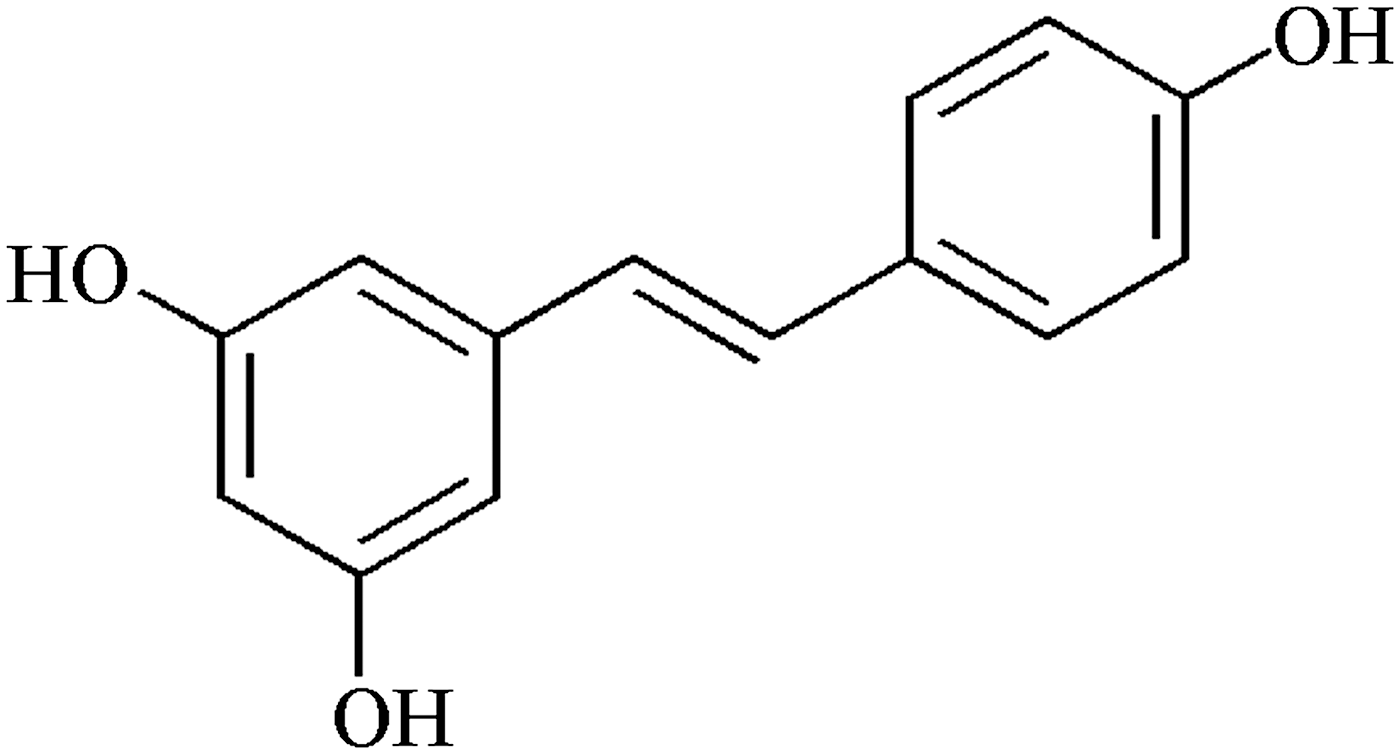
Structure of resveratrol.
Red wine chemicals have many interesting properties, including activity on endothelial cells and the vascular system. 28 For example, red wine has been shown to inhibit endothelin-1 (ET-1) synthesis, which is a highly potent peptide and vasoconstrictor. 29 High expression of ET-1 is known to contribute to the development of cerebrovascular disease and is associated with atherosclerosis. Vascular changes, including distribution changes in phosphotyrosine, 28 have been found in bovine aortic endothelial cells when ET-1 is overexpressed, which may indicate a role for red wine components in the multiple signal transduction pathways. The protective effects of flavonoids on the cardiovascular system and their anticancer, antiviral, and antiallergenic properties also are notable areas of interest. 30 –32
Of importance in the red wine debate is its ethanol content, which also can participate in the free radical reactions of biological systems by producing alkoxy and hydroxyl free radicals as the initial products. Therefore, ethanol may enhance cellular oxidative stress through the production of superoxide and hydrogen peroxide and may have deleterious effects under some conditions. Conversely, moderate ethanol consumption may be beneficial, because it has been shown to decrease inflammation and inflammatory markers in situ in adipocyte tissue explants and can attenuate whole-body inflammation and lower soluble tumor necrosis factor receptors in vivo. 33 Moreover, tumor necrosis factors (TNF) play a role in inflammation and cancer evasion, which may offer new uses for red wine components (polyphenols) as anticancer and antiinflammatory molecules. As suggested, other protective mechanism are observed with the French Paradox, and moderate red wine consumption, e.g., moderate alcohol consumption, can lower inflammatory mediators such as soluble TNF-l and -ll receptors and other cytokines. 34
Multiple antioxidant pathways have been observed for flavonoids. In particular, the flavonoids trap ROS and inhibit enzymes involved in the production of oxidative stress. Flavonoids also can regenerate other antioxidants, such as α, β, and γ-tocopherols (vitamin E). 35 The ROS-trapping mechanism and vitamin E regeneration can be explained through the oxidation of flavonoids to short-lived radicals, which degrade into relatively nontoxic products.
The French Paradox and Lipid Metabolism
Polyphenols, in particular, the catechins, are effective chain reaction–breaking antioxidants and scavengers of free radicals, thus inhibiting lipid peroxidation and oxidation of low-density lipoproteins (LDLs). LDLs are rich in polyunsaturatred fatty acids (PUFAs) and are the major cholesterol-carrying lipoproteins in plasma. Higher levels of LDL, PUFAs, and cholesterol can cause atherosclerotic lesions. The first step in the LDL oxidation process involves the oxidation of PUFAs, which are then degraded to aldehydes. Monocytes feed the process forward as they become resident macrophages and contribute to ROS in the microenvironment, where they attempt to remove the deposits through immune clearance that involves activation of several Phox enzymes and an oxidative burst. As the oxidized deposits are ingested, macrophages are subsequently transformed into foam cells, which along with oxidized LDL and apolipoprotein, are observed as components of fatty steaks in vessels. Oxidative stress by platelet aggregation and LDL oxidation can be inhibited by resveratrol in wine. 36 It has been found that the effect of resveratrol in ameliorating the oxidative damage induced by tert-butylhydroperoxide is more than that of vitamin C or vitamin E, the ubiquitous antioxidants. 37
Another possible explanation for the French Paradox and the effect of polyphenols may be explained by vitisin A and vitisin B, which showed a remarkable inhibitory activity against 3-hydroxy-3-methyl-glutaryl-coenzyme A (HMG-CoA) reductase. 38 HMG-CoA reductase is an NADPH-dependent enzyme and the rate-limiting step of the metabolic pathway that produces cholesterol and other isoprenoids, namely, the mevalonate pathway, and is a key target of the widely available statin cholesterol-lowering drugs. This may explain in part some aspect of the French Paradox as well.
Resveratrol in Caloric Restriction
The mechanism of action for resveratrol has not been elucidated fully. However, it has been reported to affect oxidative stress, longevity, chromatin organization, genetic expression, release of proinflammatory mediators, inhibition of inflammatory enzymes such as inducible nitric oxide synthase (iNOS) and cyclo-oxygenase-2 and inhibit nuclear factor κB (NF-κB) signaling. 39 One mechanism underlying some of these effects stems from reports that it slows aging in simple eukaryotes. Caloric restriction (CR) has similar actions, because it mitigates diseases of aging in mammals 40 and extends life span in a diverse variety of lower species, such as Drosophila melanogaster, Saccharomyces cerevisiae, and Caenorhabditis elegans. CR is a widely demonstrated mechanism for the extension of healthy life span, including in mice and primates. 41 Further evidence for a shared mechanism is demonstrated in resveratrol-fed elderly mice, which show a marked reduction in signs of aging, including reduced albuminuria, decreased inflammation and apoptosis in the vascular endothelium, increased aortic elasticity, greater motor coordination, reduced cataract formation, and preserved bone mineral density as compared to mice fed an ad libitum diet. 42
Clearly metabolism plays a role in resveratrol's action, which, like CR, may be inextricably linked with its role in aging. Indeed, recent data suggest this may be the case and argue for resveratrol's consideration in treating diseases that exhibit impaired metabolism, such as types 1, 2, and 3 diabetes. 43 Conserved genes that encode several classes of nicotinamide adenine dinucleotide (NAD)–dependent histone deacetylases (HDACs), also known as the sirtuins for silencing information regulatory proteins, mediate some of CR effects in lower species. One thing resveratrol reportedly does is cause substrate-specific activation of the sirtuins in yeast, Drosophila (Sir2), and humans (hSirt1). 44 The silent information regulator 2 (SIR2) gene is a life-span regulator, a property partly proposed to account for resveratrol's antiaging effects. Perhaps related to sirtuin activity is resveratrol's reputation as a putative CR mimetic, because limiting calories can result in the activation of SIR2, which has a role in regulating the physiological changes through CR. Thus, modulating the activity of SIR2 might provide potential CR mimetics and aid in fighting age-associated disease.
However, deacetylation activity is equivocal because Kaeberlein and colleagues have reported mixed findings in vitro; this group found that resveratrol enhanced binding and deacetylation of peptide substrates containing a nonphysiological fluorescent moiety. However, it had no effect on binding and deacetylation of acetylated peptides lacking the fluorophore and in three different yeast strain backgrounds, as measured by rDNA recombination, telomeric transcriptional silencing, and life span. 44 In light of these findings, the mechanism accounting for the putative longevity effects of resveratrol should be reexamined. Nevertheless, despite skepticism concerning its bioavailability, a growing body of in vivo evidence indicates that resveratrol has protective effects in rodent models of stress and disease. 45 This same observation occurs in response to CR for the brain and elsewhere. 46
Polyphenols in Neuroprotection
CR and antioxidants such as resveratrol protect cells from various cytotoxic insults and show useful effects in mouse models of aging as well as in models of Alzheimer disease (AD) and in limited clinical trials with human AD (vide supra). It has been demonstrated that polyphenols can affect spatial memory, which is enhanced by physical activity. The relationship between neurodegeneration and heart disease, the so-called heart/brain connection, has been explored for some time. 47 The French Paradox involves the cardiovasculature, which can be extrapolated to apply to the brain and cerebrovasculature as well. Main coronary occlusion depletes the blood supply to the myocardium and subsequently reduces cardiac function, which ultimately leads to heart failure. However, arterial occlusion can induce collateral artery development and reestablish and maintain blood flow to tissues such as the myocardium, which can protect against ischemic damage and cell death, as demonstrated in a rat myocardial infarction model. Resveratrol has been identified as a potential agent for the induction of new vessel growth. 48 The same authors found that resveratrol pretreatment increased antiapoptotic and proangiogenic factors, significantly upregulated protein expression for vascular endothelial growth factor (VEGF) and its tyrosine kinase receptor Flk-1/3, as well as increased both inducible and endothelial forms of NOS. Srivastava and colleagues noted increased capillary density and improved left ventricular function with resveratrol 3 weeks post myocardial infarct, and FOXO transcription factors and VEGF neutralizing antibody also enhanced the antiangiogenic effects of resveratrol. 49
Among age-associated pathologies, neuroprotection from green tea has been observed for Parkinson disease, AD, and ischemic damage. 50 One mechanism important for the protective effects for (−)-epicatechin gallate (ECG), and (−)-epigallocatechin 3-O-gallate (EGCG) in neurodegenerative diseases is their iron-chelating properties. 50 Dysregulated iron metabolism may be a central pathological feature of Parkinson disease. It is the galloyl-containing catechins, identified in a high-throughput screening, that are putative inhibitors of scrapie-associated prion protein formation. 51 Furthermore, many studies centered on consumption of polyphenols from green tea, such as epicatechin and EGCG, increased memory function and the expression of genes associated with learning, synaptic plasticity, and angiogenesis in the hippocampus. At the same time, green tea polyphenols lowered gene expression related to learning deficits and neurodegeneration.
Cytoprotective effects of catechin-rich flavanols and resveratrol include modulation of glutamatergic receptors, ion channels, and neuronal energy homeostasis. These compounds also appear to affect the cerebrovascular complications known to play a role in aging, vascular dementia, and AD, all of which have a particularly strong vascular component to the pathobiology of the disease 52 and appear to afford vascular protection 53 in a manner similar to that of ischemic preconditioning. The authors demonstrated that resveratrol pretreatment confers neuroprotection against lethal ischemic insults in the brain and other organ systems via SIRT1 activation. 54 The common factor in all these diseases is their age-related nature. In that regard, age-related cognitive decline was reversed in mice whose diets were supplemented with blueberries, spinach, or strawberries. 55 Taken together the data suggest an active lifestyle, combined with a polyphenol-rich diet, may prevent aging-related cognitive disorders 56 and neurodegenerative disease. 57
Resveratrol has been shown to reduce neuronal cell death induced by oxidized lipoproteins. 58 Oxidized lipoproteins were found to activate NF-κB binding activity and apoptosis in PC12 cells. 58 Conversely, resveratrol treatment lowered the expression of gene products that mediate inflammation (NF-κB), 59 lipid synthesis, and cell death in mice fed a high-calorie diet. 60 One question remains as to whether these compounds will prevent the damage of the sodium-potassium pump (Na+/K+) adenosine triphosphatase (ATPase) that arises out of heavy metal toxicity, specifically the damage derived from aluminum and mercury. 61,62 In that regard, it has been demonstrated in neuroblastoma SH-SY5Y cells that resveratrol may be neuroprotective by acting on the cell cycle, 63 and it is likely that resveratrol may have similar role for vulnerable neurons in AD as well.
Curcumin (Curcumene)
Curcumene (diferuoyl methane), the major component of Curcuma longa, is a free-radical sequestrating agent, and it has multiple mechanisms of action. It has been studied for its ability to modulate cancer, neurodegeneration, diabetes, and inflammation. The radical scavenging activity of curcumene is often compared to butylated hydroxyanisole (BHA), butylated hydroxytoluene (BHT), α-tocopherol, and trolox, which are prototypical reference antioxidants and radical scavengers. The antioxidant activity of curcumin (Fig. 5) has been determined through several antioxidant radical scavenging assays, namely, 2,2-diphenyl-1-picryhydrazyl (DPPH), 2,2′-azinobis [3-ethylbenzothiazoline-6-sulfonic acid (ABTS), and N,N-dimethyl-p-phenylenediamine (DMPD). 64 When curcumin was compared to other antioxidants in a lipid peroxidation assay of linoleic acid, curcumin inhibited the lipid peroxidation by 97.3% as compared to standard antioxidants: 95.4% for BHA, 99.7% for BHT, 84.6% for α-tocopherol, and 95.6% for trolox. 64 In the aforementioned study, curcumin was an effective superoxide anion and hydrogen peroxide scavenger. In addition, it had chelating properties to Fe(III) and Fe(II) ions. From the DPPH assay and superoxide trapping assay tests, all catechol-oximes show higher antioxidant properties comparable to the standards. The best radical scavengers were monohydroxylated oximes.
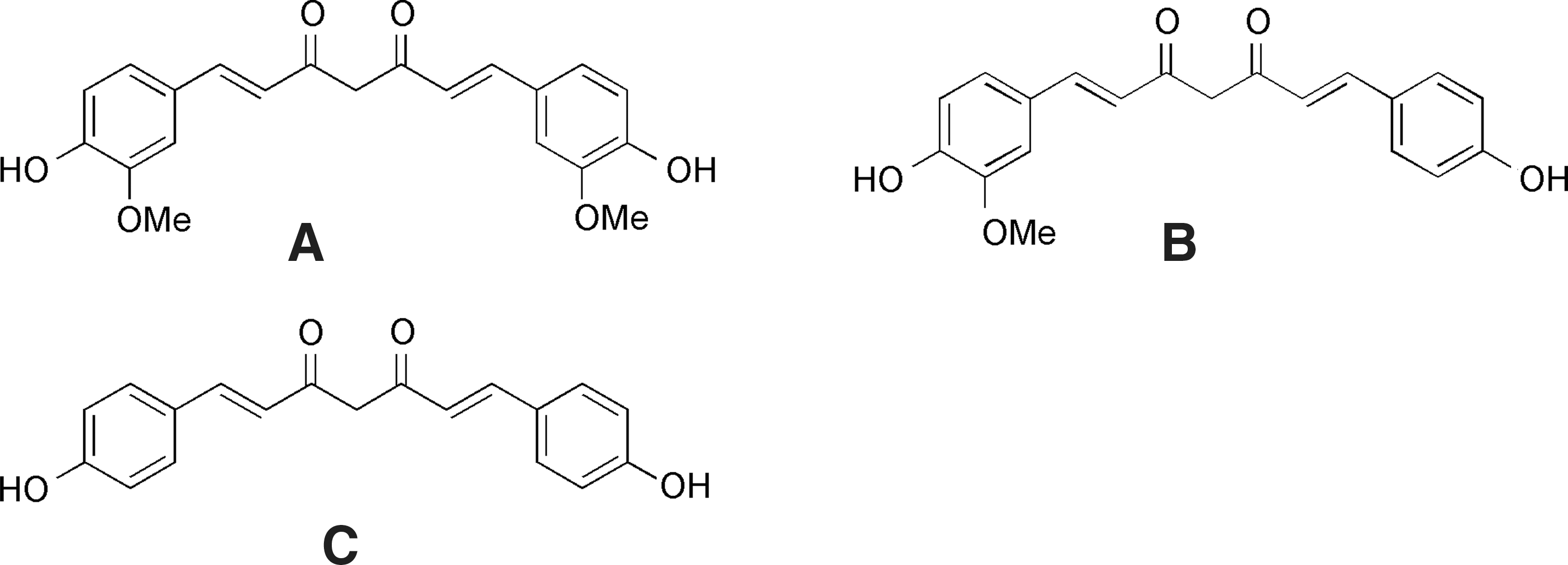
Structures of curcumenes. (
Curcumene and resveratrol exert their effects partly by inhibiting activation of NF-κB and downstream signaling. 65 Gonzales and colleagues measured cytokine gene expression for tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β, IL-6, and COX-2 with or without TNF-α stimulation in 3T3-L1–derived adipocytes and measured cytokine protein and prostaglandin E2 (PGE2) expression. The NF-κB signaling pathway was inhibited by both resveratrol and curcumin and resulted in a reduction of TNF-α, IL-1β, IL-6, and COX-2 gene expression and reduced secreted IL-6 and prostaglandin E2 (PGE2). Conversely, TNF-α treatment was found to increase IL-6, IL-1β, and COX-2 gene expression and activate NF-κB signaling in differentiated adipocytes through expected degradation of I-κB and NF-κB nuclear translocation. Furthermore, the mechanism of COX-1 inactivation by red wine meta-hydroquinones 66 adds a complexity to the question of what redox form and structure these compounds take when delivering their apparent activity. Taken together, these data suggest that curcumene, resveratrol, or other flavonols may reduce or inhibit chronic inflammatory stress and concomitant cytokine expression 67 from adipose tissue (most likely from visceral fat). Curcumene and resveratrol both may improve cardiovascular function and insulin sensitivity and, if so, this may offer new clues into mechanisms of the French Paradox.
Catechins
Catechin-derived flavonoids are abundant in human diets. Green tea is a growing dietary source of antioxidants, which contains numerous polyphenols, such as (−)-epicatechin (EC1), (+)-catechin, epigallocatechin (EGC), ECG, and EGCG (Figs. 1 and 6).
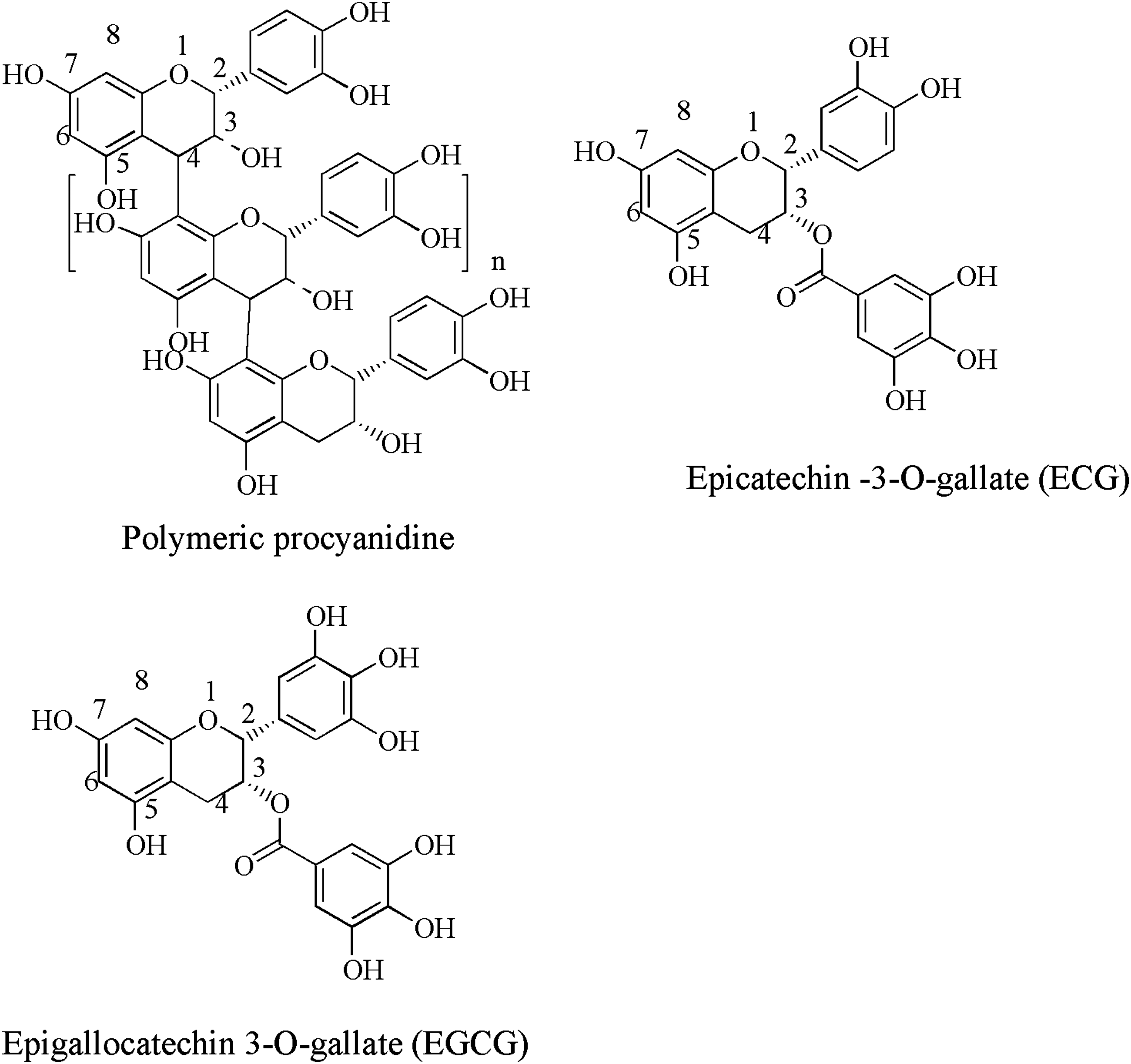
Structure of polymeric procyanidine, (−)-epicatechin gallate (ECG), and (−)-epigallocatechin 3-O-gallate (EGCG).
Flavanols occur as monomeric, oligomeric, or polymeric forms known as proanthocyanidines. Procyanidine is a subunit of proanthocyanidines, which are present in cereals, vegetables and fruits, and wines. The monomeric unit of proanthocyanidines contains mainly epicatechins and/or catechins (Fig. 6). These units are linked mainly through C4–C8 or C4–C6 bonds (B-type). 68 However, other conjugates, such as cysteinyl-flavan-3-ol conjugates from grape procyanidins have been found that could have improved antioxidant properties. 69 Another antioxidant activity involves the dihydroxylation of rings within catechins and quercetin. 21 Catechins containing a gallate ester moiety at the 3-position (ECG and EGCG) have the highest activity as antioxidants and are the most effective inhibitors of lipid peroxidation. Phenolic hydroxyl groups explain the mechanism of action for catechin antioxidant activity on the B-ring of ungalloylated catechins (EC and EGC) and on the B- and D-rings of the galloylated forms (ECG and EGCG). 70 Epigallocatechin gallate was found to bind to CD4 and interferes with gp120 binding, 71 which may offer an explanation for EGCG's possible role in the treatment of human immunodeficiency virus/acquired immunodeficiency syndrome (HIV/AIDS).
Catechins, also known as tannins, are secondary plant metabolites that were originally recognized because they interacted strongly with collagen both in vivo and in situ. They are used in the process that bears its name (tanning), which converts animal skin to leather. Tannins were subsequently defined as high-molecular-weight polyphenol polymers that precipitate protein from solution. Of key importance is the bioavailability of these compounds in vivo, which depends on the degree of polymerization and will be discussed later. Oligomeric forms of these compounds were found to be more bioavailable and demonstrated high antioxidant activity when compared to their monomeric or polymeric forms with the order of radical scavenging effectiveness as follows: ECG > EGCG > EGC > EC > catechin. Depolymerizations of proanthocyanidines are generally done by thiolysis, which involves cleavage of interflavonoid linkages and nucleophilic attack by thiol groups at the C4 position. Depolymerization using other nucleophiles such as
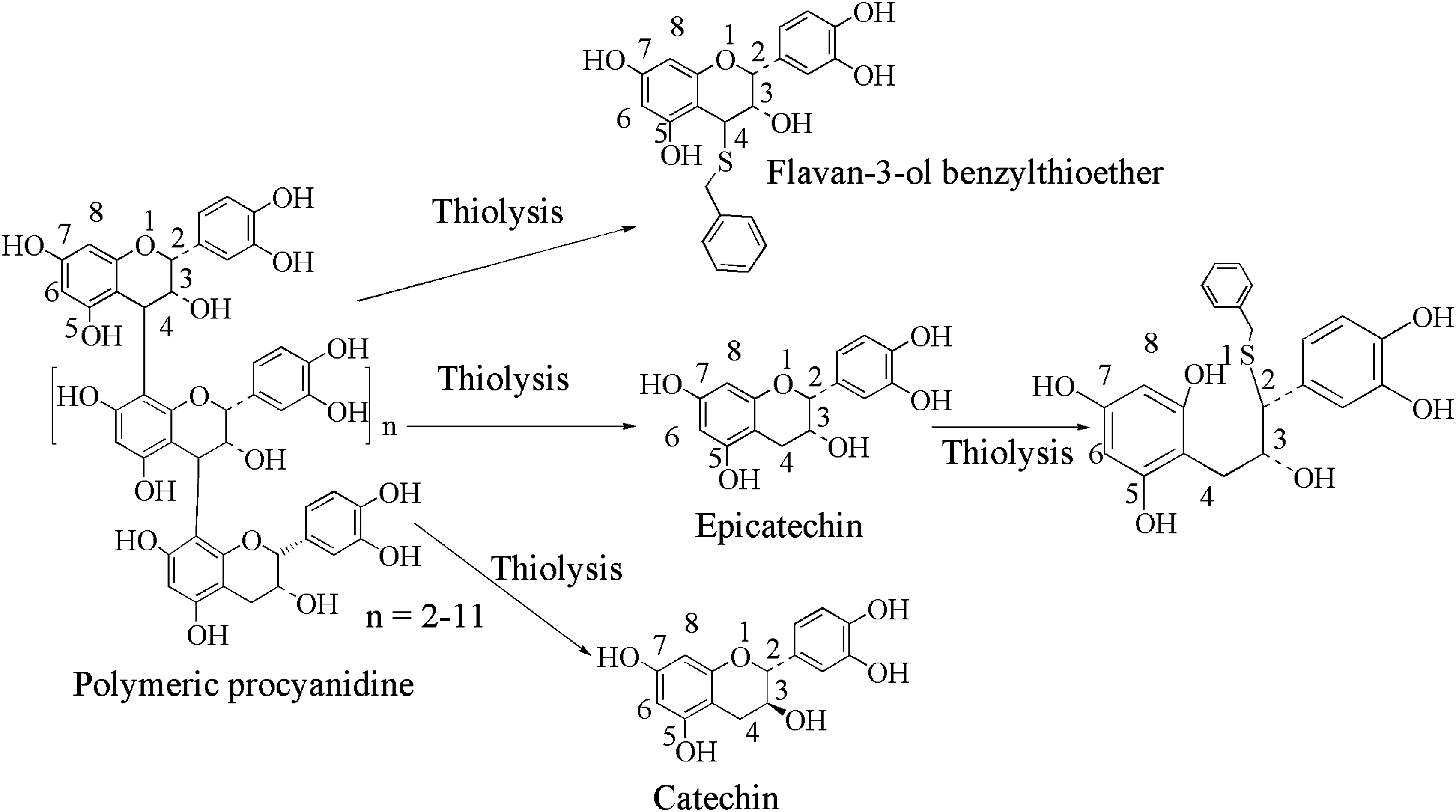
Thiolysis of procyanidins to give monomeric flavonoids and their derivatives.
These catechines are responsible for a variety of physiological activities, such as antioxidant, antimicrobial, 73 antiviral, 74 and antiangiogenic activities, 75 and selectively modify estrogen receptors (ERs), which is why they were explored in gynecological cancers, including metastatic breast cancer, with promising results. 76 The authors of the study explored the combination of resveratrol, quercetin, and catechin in cancer, which significantly reduced cell proliferation and blocked cell cycle progression in vitro and in nude mice with breast cancer xenografts. 76 Of interest in this study, peak inhibition was observed at 5 mg/kg body weight, which is a considerable amount of compound when proposed for use in humans. Several epidemiological studies as well as studies also have shown that green tea has chemopreventive properties against various other cancers, 30 such as those of the skin 77 , lung, 78 and prostate. 79
Other laboratory studies suggest that flavonoids are antimutagenic and anticarcinogenic. The antioxidant and anticancer properties of green tea were explored in a population-based case–control study, where an inverse association among tobacco smokers between lung cancer and the consumption of epicatechin, catechin, quercetin, and kaempferol was found; this study concluded that teas might protect smokers from lung cancer. 80 Furthermore, a randomized and controlled study demonstrated a positive effect that increased tea consumption (4 cups per day) had on oxidative DNA damage among smokers. 81 Although these large population studies are needed to establish any clinical relevance for polyphenols in preventing or treating disease, some authors do appreciate the value of mixtures of compounds in studies, which seemed to offer more benefit over using one single polyphenolic antioxidant.
Other health benefits attributed to green tea and its catechins include antiaging 82 and antiinflammatory activities 83 ; free radical chain breaking molecules 84 have the ability to lower cholesterol 85 and can prevent the development of atherosclerotic plaques. 86 It also has been shown to have antihypertensive activity, 87 antidiabetic properties in vivo and in models of insulin resistance, 88 as well as use as an approach to weight loss. 89 Green tea epigallocatechin 3-gallate accumulates in mitochondria and displays a selective antiapoptotic effect against inducers of mitochondrial oxidative stress in neurons. 90
In studies involving diabetes and related mitochondrial dysfunction, Rutter and co-workers showed that a commercial green tea extract suppressed the age-related increases in collagen crosslinking and fluorescent products in C57BL/6 mice. 91 However, when diabetic or euglycemic rats were treated with fresh-brewed Japanese green tea leaf extract, total crosslinking worsened and increased glycoxidation in tendon, aorta, and plasma was either worsened or had no significant improvement with green tea. The high content of tannins in the tea is possibly the best explanation for this finding. However, erythrocyte glutathione and plasma hydroperoxides were improved by green tea (p < 0.001), as was retinal superoxide production, lens crystallin fluorescence at 370/440 nm (p < 0.05), and the formation of acellular retinal capillaries or retinal pericyte ghosts. 92 In the mitochondria, renal NADH-linked adenosine diphosphate (ADP)-dependent and dinitrophenol-dependent respiration and complex III activity were improved by green tea, which also suppressed the methylglyoxal hydroimidazolone mitochondrial protein modification. Because green tea improved several diabetes-related cellular dysfunctions, but worsened matrix glycoxidation in selected tissues, it appeared as though the reaction shifted from oxidative stress to carbonyl stress, at least in the extracellular compartment. Supposedly, many of the tannins were removed from the commercial green tea extract, which could offer a partial explanation for these apparently dichotomous results.
Other trans-resveratrol oligostilbenoids and viniferins (Fig. 8), present in the human diet may be of importance as cancer chemopreventive agents, and epidemiological studies suggest that they could prevent malignant tumor development due to cytostatic and cytotoxic effects on tumor cells and cell lines. In that regard, trans-resveratrol analogs were tested for inhibitory effects on cell cycle progression proliferation and apoptosis of human colon tumor cells 93 and lymphoid and myeloid cell lines. 94
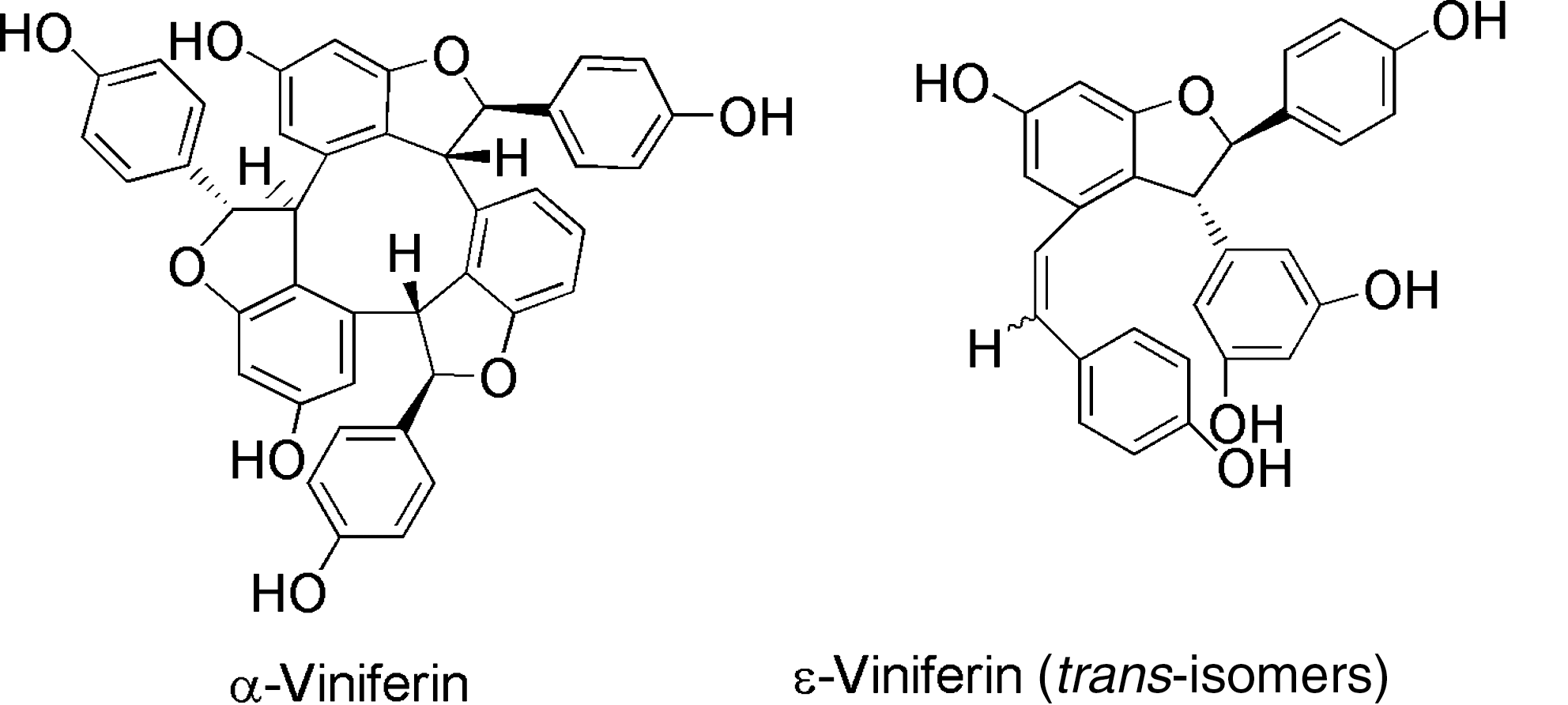
Structures of naturally occurring oligomers of resveratrol (trans-3,4,5-trihydoxystilbene), ε-viniferin, and α-viniferin.
The antitumor activities of resveratrol dimer, epsilon-viniferin, and acetylated forms (resveratrol triacetate, epsilon-viniferin penta-acetate) and of vineatrol (a grape extract) were compared on human colon adenocarcinoma and other cell lines with mixed results. Resveratrol triacetate and vineatrol were as efficient as trans-resveratrol at inducing the cell cycle accumulation of human colon cancer cells in early S phase, which associated with nuclear cyclin A redistribution and formation of cyclin A/cyclin-dependent kinase 2 complex. 93 The same authors demonstrated resveratrol and its triacetate form inhibited cell proliferation and caused cell cycle arrest, whereas ε-viniferin and ε-viniferin-pentaacetate only slightly reduced cell proliferation, favoring S phase accumulation. However, others failed to demonstrate apoptosis or cytotoxic activity of viniferin, trans-amurensin-B or trans-ε-viniferin on three cancer cell lines in vitro. 95,96 These trans-resveratrol analogs retained the cytostatic and cytotoxic activities of the parent molecule and were considered as chemo-sensitizers for 5-fluorouracil (5-FU)–mediated proliferation inhibition in human-derived colon cancer cells.
Proliferation assays performed on myeloid and lymphoid cell lines show an antiproliferative and proapoptotic effect of resveratrol with two of its naturally occurring oligomers, ε-viniferin (a dimer) and miyabenol C (a trimer). 94 ε-Viniferin and, more importantly, miyabenol C are potent antitumor agents, when used in combination with resveratrol, on all cell types tested. They modify cell cycle distribution; i.e., cells treated with resveratrol accumulated in S phase, whereas those treated with ε-viniferin and miyabenol C accumulated in G2/M and G0/G1, respectively. 94 All of these compounds induced apoptosis in U266 cells via mechanisms entirely dependent on upstream caspase activation (8 and/or 2), depending on the compound. The activation was independent of Fas/Fas ligand interaction, without acting directly on the mitochondrial membrane.
In myocytes, trans-3,5,4-trihydroxystilbene has been shown to have various cardioprotective effects. 97 Vitisin A facilitated mitochondria swelling, depolarization, and cytochrome c release in a dose-dependent manner. There is an adage that says what is good for the heart is good for the brain, and polyphenols are not an exception. Keeping with the notion of cerebrovascular component to AD, the resveratrol dimer (+)-vitisinol E demonstrated inhibitory activity in vitro on BACE-1 (β-site APP-cleaving enzyme 1), in a dose-dependent manner. 98
Bioavailability and Potentially Harmful Effects of Polyphenolic Antioxidants
The polyphenolic food components must be bioavailable to exert any biological effects. The medicinal potential of polyphenols, such as curcumin, can be severely affected by limited systemic and target tissue bioavailability and rapid metabolism. Data regarding polyphenol absorption and metabolism on the absorption and tissue distribution of polyphenols are derived mainly from animal studies. 99–100 We can extrapolate animal findings to humans, but differences between the genomes and epigenetic phenomena potentially complicate these models. In that regard, rodents methylate phenols more extensively than humans. Nevertheless, it is apparent that most classes of polyphenols are sufficiently absorbed to exert biological effects. 101,102 Furthermore, repeat dosing, which may reflect long-term consumption of polyphenols, is shown to increase bioavailability. 103 Models offering different drug delivery systems, such as liposomes, phospholipid complexes, and nanoparticles, may address bioavailability. Some of the difluorocurcumin analogs that allow longer circulation times and preferential accumulation are now in clinical trials for prevention of tumor progression and/or treatments of malignancies. 104
However, many polyphenolic compounds such as the proanthocyanidins, tannins, and lignins exist in the polymeric or oligomeric forms or become polymerized into larger molecules or concatenations. A caloric restriction mimetic effect of resveratrol combined with tannins, such as those found in red wine, simply may be based on an ability to limit intestinal absorption of foods. A daily intake of tannin from foods, including fruits, beverages, and some grains, which contain condensed and hydrolyzable tannins, is estimated as roughly 1 gram. Tannins are known to bind to proteins in soluble or precipitable complexes, which can reduce nutrition in livestock feed. 105 A diet heavily enriched with tannins and tannic acid leads to impaired nutrition in domestic animals. 106 In that regard, a tannin-rich diet also is known to block absorption of several nutrients. We may apply this data to humans who consume high quantities of tannins, because there are several mechanisms that may explain impaired nutrition with plant polyphenol ingestion.
Salivary proline-rich proteins (PRPs), particularly the basic PRPs, form insoluble complexes with condensed tannins and are suggested to act as a type of bioflocculant by forming complexes with them. 107 The protein-binding action of these complexes would affect polyphenol reactivity as well as prevent interaction with other biological compounds or intestinal absorption 108 and influence its biological fate and the localization critical for function. 109 Tannin complexes can form in the gastrointestinal (GI) tract and are stable when prepared in vitro at GI pH as well as in the presence of proteases (pepsin, trypsin, chymotrypsin, carboxypeptidase A and B, and elastase) and the bile acids. 110
Another important mechanism involving protein and oxidized polyphenols includes the chemical or enzymatic processes, as well as the nonenzymatic browning reactions of plant polyphenols and their effects on amino acid and protein bioavailability. The interaction of quinones and amino acids has been described by Bittner 111 and is usually compartmentally separated in living systems. However, junctions where quinones react and influence the function of proteins can occur before or during food processing or in vivo with potentially damaging consequences.
Effects of Polyphenolic Antioxidants on Iron Bioavailability
Compounds that inhibit iron availability are linked to anticancer benefits and may be one explanation for the effect observed with consumptions of polyphenols. A study by Boato and colleagues confirmed this finding; they explored uptake of iron in combination with fortified cereals or fruit juices in a gastric digestion protocol. 112 Orange juice and white grape juice significantly increase iron bioavailability, whereas red grape juice, prune juice, and cranberry juice 113 have profound inhibitory effects on iron bioavailability. 112 It is well-known that ascorbic acid promotes iron bioavailability, whereas the polyphenolic compounds seem to bind and prevent absorption of soluble iron. The iron uptake inhibitory effects were attributed to high levels of polyphenolic compounds, which show their iron-complexing properties. 113 The extracellular thiol/disulfide redox state affects proliferation rate in a human colon carcinoma (Caco2) cell line, 114 and polyphenols are believed to contribute to thiol reduction directly and through chelation.
The iron-binding properties of quercetin, and other select polyphenolic compounds (chrysin, 3-hydroxyflavone, 3′,4′-dihydroxy flavone, rutin, and flavone) were investigated by Guo and colleagues. 113 These authors confirmed strong iron-binding properties for the compounds containing the "iron-binding motifs" in their structures. Surprisingly, quercetin, an oxygen-based ligand, binds Fe(II) even stronger than the well-known strong nitrogen-based Fe(II)-chelator ferrozine at pH 7.2. The strong iron-binding properties of polyphenols argue that they may be effective in modulating cellular iron homeostasis under physiological conditions. Dysregulation of iron homeostasis is thought to play a role in AD and other neurodegeneration. However, there is conjecture about the uptake and transport of these large aromatic molecules into brain tissues. 115 Nevertheless, quercetin can completely suppress Fenton chemistry both at micromolar levels and in the presence of major cellular iron chelators like adenosine triphosphate (ATP) or citrate. However, the radical scavenging activity of quercetin provided only partial protection against Fenton chemistry-mediated damage, whereas Fe(II) chelation by quercetin was found to completely inhibit Fenton chemistry, 113 indicating that the chelation may be key to its antioxidant activity. 111 Regardless, these results demonstrate that quercetin and other phenolic compounds can effectively modulate iron biochemistry under physiologically relevant conditions, providing possible insight into the mechanisms of action for bioactive polyphenols.
In summary, significant work has been done in regard to the use of polyphenolic compounds to treat age-related diseases, although some controversy exists as to some of their beneficial and harmful effects. Specifically, we need to explore the exact mechanism through which these compounds exert their protective or deleterious effects to maximize their efficacy and safety when treating disease. One aspect of these antioxidant compounds, which has not been well studied, is that these products involve reactive quinines or quinoid compounds as well as dimers and complex oligomers. More work is needed in this area to explore the effect of these reactive intermediates on proteins, lipids, and their implication in health and disease. More important roles for these compounds may involve changing paradigms, because they hold promise for preventing many diseases, including some deleterious effects from aging.