
Abstract
Danshen-Gegen (DG) decoction, an herbal formulation comprised of Radix Salvia Miltiorrhiza and Radix Puerariae Lobata, is prescribed for the treatment of coronary heart disease in Chinese medicine. Experimental and clinical studies have indicated that DG decoction can reduce the extent of atherosclerosis. In the present study, using an ex vivo rat model of myocardial ischemia/reperfusion (I/R) injury, we investigated the myocardial preconditioning effect of an aqueous DG extract prepared from an optimized weight-to-weight ratio of Danshen and Gegen. Long-term treatment with DG extract at increasing doses (including the equivalent of a human dose) protected against myocardial I/R injury in rats. The cardioprotection afforded by DG pretreatment was paralleled by enhancements in mitochondrial antioxidant status and membrane integrity, as well as a decrease in the sensitivity of mitochondria to Ca2+-stimulated permeability transition in vitro, particularly under I/R conditions. Long-term treatment with the DG extract enhanced the translocation of protein kinase C-epsilon (PKCε) from the cytosol to mitochondria in rat myocardium, and this translocation was inhibited by α-tocopherol co-treatment with DG extract in rats. Long-term DG treatment may precondition the myocardium via a redox-sensitive PKCε/mKATP pathway, with resultant inhibition of the mitochondrial permeability transition. The results suggest that clinical studies examining the effectiveness of DG extract given prophylactically in affording protection against myocardial I/R injury would be warranted.
Introduction
Danshen and Gegen, which are roots of Salvia miltiorrhiza and Puerariae lobata, respectively, are traditional Chinese medicinal herbs that have been widely used in China, Japan, Korea, and Taiwan for the treatment of cardiac and cerebral ischemic conditions. 5,6 The herbal formulation made of Danshen and Gegen (referred to as “DG decoction”) has long been prescribed for the treatment of coronary heart disease in the practice of Chinese medicine. 7 Recently, it has been shown that treatment with DG decoction modulates key early atherogenic events in human monocyte-derived macrophages and thereby inhibit the formation of foam cells. 8 DG decoction has also been found to be effective in improving vascular structure and function in patients suffering from cardiovascular diseases. 9 Given the traditional use for the treatment of coronary heart disease, DG decoction may also protect the myocardium against I/R injury. However, the cardioprotective action of DG decoction has not been fully investigated. In the present study, an aqueous extract was prepared from a mixture of Danshen and Gegen in an optimized weight-to-weight ratio in terms of biological activity, and its effectiveness in ameliorating I/R injury was investigated using an ex vivo rat heart model system. The biochemical mechanism underlying the cardioprotection was investigated by examining changes in mitochondrial antioxidant status and structural integrity, as well as the sensitivity of mitochondria to Ca2+-induced permeability transition in control and ischemic/reperfused rat heats. The involvement of a PKCε- and mKATP-mediated signaling pathway in the cardioprotection afforded by DG was also investigated using specific inhibitors of these biochemical entities.
Materials and Methods
Preparation of herbal extracts
Radix Salviae Miltiorrhiza (Danshen) and Radix Puerariae Lobatae (Gegen) were purchased from mainland China. They were authenticated by an in-house herbalist at the Institute of Chinese Medicine (ICM) at the Chinese University of Hong Kong, Hong Kong. Voucher specimens of Danshen (#2008-3088b) and Gegen (#2008-3167b) were deposited in the ICM. DG extracts with varying weight-to-weight ratios of Danshen to Gegen (0:10, 3:7. 4:6, 5:5, 6:4, 8:2, 9:1, 10:1) were prepared using a laboratory scale protocol. In brief, herbs were cut into small pieces, and a total amount of 1.65 kg of herbal mixture was soaked in 9 L of distilled water for 1 h, followed by extraction with boiling water under reflux conditions for 1 h. The extraction procedure was repeated once, and the pooled aqueous extract was centrifuged at 600 × g for 20 min at 20°C. The resultant supernatant was concentrated under reduced pressure at 60°C, and the concentrate was dried by lyophilization. The yields of herbal extracts prepared with different ratios of Danshen to Gegen varied from 31% to 47% (w/w, with respect to raw herbs), with the highest and lowest values obtained from Danshen and Gegen per se, respectively.
DG extract with the optimal ratio of Danshen to Gegen (7:3), as assessed by cardioprotection against I/R injury (Fig. 1), was prepared in large scale for experimental and clinical investigations. Herbs were soaked in water (1:10, w/v) for 75 min, followed by extraction in boiling water for 1 h. The extraction procedure was repeated twice with boiling water (1:8) for 1 h and 30 min, respectively. The pooled aqueous extracts were concentrated under reduced pressure at 60°C, and the concentrate was spray-dried to obtain the powdered form of DG extract at a yield of 10.1%.
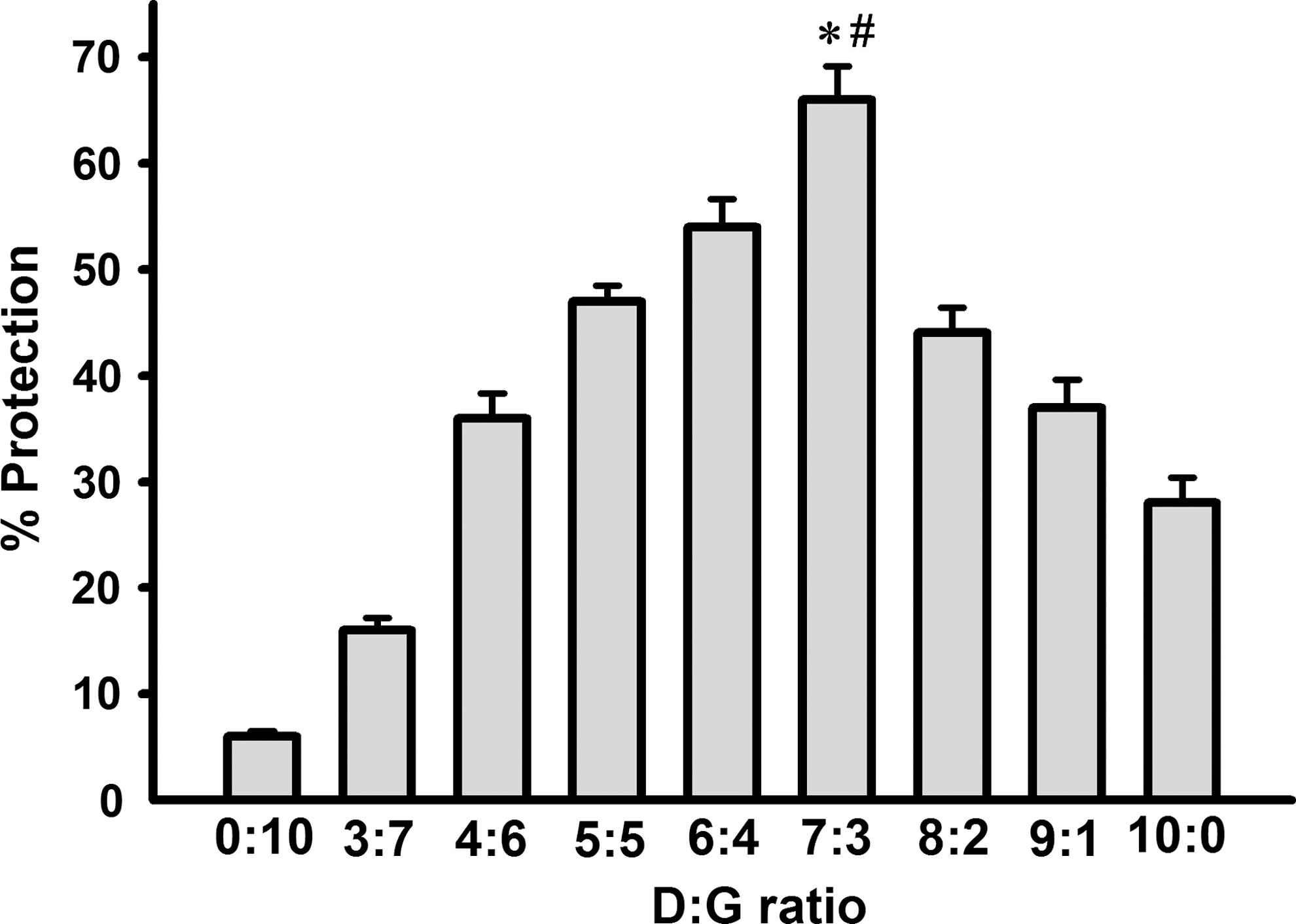
Effects of herbal extracts with varying ratios of Danshen (D) and Gegen (G) on myocardial ischemia/reperfusion injury in rats. Animals were administered the herbal extracts orally at a daily dose of 1 g/kg for 3 days. Control animals received the vehicle (i.e., water) only. Twenty-four hours after the last dosing, isolated-perfused hearts were subjected to 15 min of ischemia followed by 10 min of reperfusion. The extent of myocardial ischemia/reperfusion (I/R) injury was assessed by lactate dehydrogenase (LDH) leakage, as described in Materials and Methods, and data were expressed in percent protection with respect to the DG un-treated control. Values given are means ± standard error of the mean (SEM), with n = 5. (*) Significantly different from the D:G (6:4) group; (#) significantly different from the D:G (8:2) group.
Chemical analysis of the DG extract
Identification and quantification of the major components in the DG extract were performed according to our previous study with minor modifications regarding the instrument and chromatographic conditions. 10 Briefly, a Waters high-performance liquid chromatography (HPLC) system (Waters, Milford, MA) equipped with a 2695 solvent delivery module and a 996 photodiode UV detector was used. The chromatographic separation of the analytes was achieved by an Agilent Eclipse XDB-C18 column (5250 × 4.6 mm i.d.; 5-μm particle size) connected to an Agilent C18 guard column. The mobile phase, consisting of 0.5% acetic acid in acetonitrile (solvent A) and 0.5% acetic acid in water (solvent B), was run with gradient elution at a flow rate of 1 mL/min. The linear gradient elution was carried out as follows: Solvent A was kept at 5% for the first 5 min, and then increased to 10%, 17%, 35%, and 90% in the next 13 min, 12 min, 10 min, and 30 min, respectively, then returned to 5% in 5 min and equilibrated for 15 min before the next injection. HPLC analysis indicated that the DG extract contained the following marker compounds (μg/100 mg; mean ± standard deviation [SD], n = 3): Danshensu (1,868.2 ± 33.7), salvianolic acid B (1,345.7 ± 18.5), protocatechuic aldehyde (78.3 ± 3.9), puerarin (1,760.1 ±23.4), daidzein 8-C-apiosyl-glucoside (404.1 ± 8.1), daidzin (159.4 ± 3.3), and daidzein (162.9 ± 1.4).
Animal care
Female Sprague-Dawley rats (8–10 weeks old, weighing 200–250 g) were maintained under a 12-h dark/light cycling at 22°C and allowed food and water ad libitum in the Animal & Plant Care Facility at the Hong Kong University of Science and Technology (HKUST, Hong Kong, China). All experimental protocols were approved by the University Committee on Research Practice at HKUST. Male or female rats could be used for the model of myocardial I/R injury, but female rats were chosen because of their availability.
Drug treatment
Animals were randomly divided into groups of 4–6 animals in each. In preliminary screening experiments, rats were treated intragastrically by gavage with DG extracts of varying composition at a dose of 1 g/kg for 3 days. In long-term DG treatment, rats were treated intragastrically with DG extract (7:3, dissolved in water) twice daily at doses of 0.09, 0.15, and 0.45 g/kg for 14 days. Control rats received water only by gavage. Twenty-four (for the screening experiments) or 12 (for the long-term treatment studies) hours after the last dosing with the DG extract, hearts were isolated from phenobarbital-anesthetized animals and subjected to I/R challenge, as described below. To determine the role of PKCε and mKATP in the effects of long-term DG treatment on I/R injury, isolated hearts were perfused with 2 μM PKCε translocation inhibitor (peptide EAVSLKPT; Calbiochem, Darmstadt, Germany) or 2 μM 5-hydroxydecanoate (5-HD; Sigma Chemical Co., St. Louis, MO) in Krebs–Henseleit buffer for 15 min prior to I/R challenge.
Isolated-perfused rat hearts
Hearts were quickly excised and immediately immersed in ice-cold and heparinized (50 U/mL) saline. The aorta was cannulated and transferred to a warm, moist chamber (double-walled jacket with temperature being maintained at 37°C by circulating warm water) of the Langendorff perfusion apparatus (Radnoti, Monrovia, CA). The hearts were perfused retrogradely at a constant pressure of 70 mmHg with Kreb–Henseleit solution of pH 7.4 gassed with 95% O2 and 5% CO2, as previously described. 11
Myocardial I/R injury
After an initial 30-min of perfusion for equilibration, the isolated heart was subjected to a 15-min (screening experiment) or 40-min (long-term study) period of "no-flow" global ischemia by stoppage of perfusate, followed by a 10-min or 20-min reperfusion, respectively. For convenience, brief durations of ischemia and reperfusion were adopted for screening experiments. Coronary effluent was collected in 1-min fractions at various time intervals during the course of equilibration and reperfusion. The fractions were immediately placed on ice until assayed for lactate dehydrogenase (LDH) activity. The extent of LDH leakage during the reperfusion period, an indirect index of myocardial injury, was estimated by computing the area under the curve (AUC) of the graph plotting the percent LDH activity (with respect to the mean preischemic value measured during the equilibration period) against the reperfusion time (1–20 min), as previously described, 11 and the value was expressed in arbitrary units. Non-I/R hearts were perfused for 90 min. After the non-I/R or I/R procedure, ventricular tissue samples were obtained and subjected to biochemical analysis.
Preparation of cytosolic and mitochondrial fractions
Cytosolic and mitochondrial fractions were prepared by differential centrifugation, as described by Starnes et al. 12 Myocardial ventricular tissue samples were rinsed with ice-cold isotonic buffer (210 mM mannitol, 70 mM sucrose, 5 mM HEPES, 1 mM EGTA [pH 7.4], 0.2 mg/mL soybean trypsin inhibitor, 0.2 mg/mL bacitracin, 0.16 mg/mL benzamidine). Tissue homogenates were prepared by homogenizing 0.6 g of minced tissue in 6 mL of ice-cold isotonic buffer using a Teflon-in-glass homogenizer (Glas-Col, Terre Haute, IN) at a speed of 1,600 rpm for 20 strokes on ice. The homogenates were centrifuged at 600 × g for 20 min at 4°C. After collecting the supernatants, the pellets were resuspended with the same volume of ice-cold homogenizing buffer (but without the protease inhibitors) and recentrifuged at 600 × g. The procedure was repeated twice. The pooled supernatants (a total of 4 volumes) were centrifuged at 9,200 × g for 30 min, and the mitochondrial pellets were collected. The supernatants were saved for the preparation of cytosolic fractions. The mitochondrial pellets were then washed with the same volume of ice-cold sucrose buffer containing 210 mM mannitol, 70 mM sucrose, and 5 mM HEPES-KOH (pH 7.4), and the mixtures were centrifuged at 9,200 × g for 30 min. The washing procedure was repeated. The mitochondrial pellets were resuspended in 1.0 mL of ice-cold sucrose buffer, and these constituted the mitochondrial fractions. Cytosolic fractions were prepared by centrifuging the above supernatant at 100,000 × g for 60 min at 4°C. The protein concentrations of the mitochondrial and cytosolic fractions were determined using a protein assay kit (Bio-Rad, Hercules, USA).
Mitochondrial reactive oxygen species generation
The extent of mitochondrial reactive oxygen species (ROS) generation in vitro was measured as previously described. 13 An aliquot (50 μL) of mitochondrial fraction (50 μg protein/mL) and 60 μL of 2′,7′-dichlorofluorescin diacetate (DCFDA) (Fluka, Switzerland) solution (5 μM in the incubation buffer) was added to wells of a black microtiter plate. The mixture was incubated at 37°C for 10 min in the dark in a Victor 3TM Multi-Label Counter (Perkin-Elmer, Wellesly, MA). After the incubation, 50 μL of incubation buffer (0.1 mM EGTA, 5 mM KH2PO4, 3 mM MgCl2, 145 mM KCl, 30 mM HEPES [pH 7.4]) and 50 μL of substrate solution (20 mM pyruvate and 10 mM malate) were added. Fluorescence measurements (excitation 485 nm and emission 535 nm) of the reaction mixture were taken every 5 min for 30 min. Mitochondrial ROS generation was determined using the fluorescence intensity of the sample after subtracting the value of a blank sample containing incubation buffer, substrate solution, and DCFDA. The extent of ROS generation over the 30-min period of incubation was estimated by computing the AUC of the graph plotting fluorescent intensity against time (0–60 min) and expressed in arbitrary units.
Mitochondrial Ca2+ content
Mitochondrial Ca2+ content was measured using a Ca2+-sensitive fluorescence probe Fluo-5N AM ester (Molecular Probe, OR), as previously described. 14 The Ca2+ dissociation constant (K d) was determined using a Ca2+ calibration kit in a concentration range of 1–1,000 μM, with the K d value being estimated to be ≈98 μM, which is in good agreement with the data provided by the manufacturer. An aliquot (25 μL) of mitochondrial fraction (0.5 mg/mL final concentration) was mixed with 25 μL of incubation buffer (100 mM KCl, 30 mM MOPS [pH 7.2]) in 96-well black microtiter plates. The mixture was incubated at 25°C for 15 min and then added with 25 μL of digitonin (50 μg/mL) and 25 μL Fluo-5N AM ester (1 μM in 0.005% Pluronic F-127). The reaction mixture was incubated at 25°C for 30 min, and the fluorescence reading was measured at excitation and emission wavelengths of 488 nm at 532 nm, respectively, using Victor 3TM Multi-Label Counter. The mitochondrial Ca2+ content was estimated from a standard calibration curve and expressed in μmol/mg protein.
Mitochondrial cytochrome c release
Mitochondrial cytochrome c release was indirectly assessed by the measurement of cytosolic cytochrome c levels using western blot analysis, as previously described. 15 Total cytosolic fractions with equal amounts of protein (40 μg protein) were subjected to 15% sodium dodecyl sulfate–polyacrylamide gel elecrophoresis (SDS-PAGE), followed by immunoblotting using specific antibodies of cytochrome c (clone 7H8.2C12, BD PharMingen, San Diego, CA). The extent of mitochondrial contamination in the cytosolic fractions was determined using specific antibodies against complex IV. The protein-blot analysis was carried out using the ECLTM Western Blotting System (Cell Signaling Technology, Beverly, MA) according to the manufacturer's recommendations, and the protein bands were quantified by densitometry. The cytochrome c release was estimated from the amount (arbitrary units) of cytochrome c normalized with reference to actin (1:5000, Sigma Chemical Co, St Louis, MO) levels (arbitrary units) in the sample.
Mitochondrial permeability transition
Mitochondrial swelling was taken as an indirect measure of MPT in vitro, as previously described. 16 An aliquot (1.6 mL) of mitochondrial sample (0.5 mg protein/mL) was prepared by mixing the mitochondrial fraction with incubation buffer containing 125 mM sucrose, 65 mM KCl, 10 mM HEPES (pH 7.2), 5 mM succinate (freshly prepared), and 5 μM rotenone (freshly prepared). Aliquots (200 μL) of mitochondrial homogenate were mixed with 10 μL of cyclosporin A (CsA) (5 μM) in 0.5% (w/w) ethanol (final concentration in incubation buffer). The mixtures were incubated at 30°C for 5 min. An aliquot (10 μL) of calcium chloride (Ca2+) solution (1 μM final concentration) was then added, and the mixtures were incubated at 30°C for 5 min. Aliquots (180 μL) of the mixtures were added into 96-well microtiter plate, and the initial absorbance at 520 nm was monitored for 5 min at 30°C. The swelling reaction was then initiated by adding 20 μL of potassium phosphate (0.5 mM, pH 7.2), and the absorbance at 520 nm of the reaction mixtures was read every 2 min for 30 min at 30°C, using Victor 3TM Multi-Label Counter. The extent of mitochondrial swelling was estimated by computing the AUC of the declining graph plotting percent initial absorbance (100% as baseline) against time (min) to obtain AUC1. The extent of MPT (ΔAUC1) was estimated by subtracting the AUC1 with CsA from the AUC1 without CsA. The Ca2+-induced MPT was expressed as the ratio of ΔAUC1 induced by both Ca2+ and PO4 3− to that induced by PO4 3− only.
Western blot analysis for PKCε
Mitochondrial and cytosolic fractions were prepared from myocardial ventricular tissue samples by differential centrifugation as described above, except using a different homogenizing buffer,which contained 300 mM sucrose, 4 mM HEPES, 2 mM EGTA, 1 mM PMSF and 20 μM leupeptin (pH 7.4). After centrifugation, the resultant pellets were resuspended in ice-cold mitochondrial buffer (20 mM Tris-HCl, 1 mM EGTA, 1 mM phenylmethylsulfonyl fluoride [PMSF], 20 μM leupeptin, 0.5% Triton X-100, pH 7.4) and sonicated on ice for 3 min, and this constituted the mitochondrial fraction. The cytosolic and mitochondrial fractions were mixed with 6 volumes of SDS sample loading buffer (1 M Tris-HCl, pH 6.8, 10% SDS, 30% glycerol, 6 mM dithiothreitol [DTT], and 0.2 mM Bromophenol Blue). Mitochondrial and cytosolic proteins (50 μg) were resolved using 10% SDS-PAGE gels and transferred onto nitrocellulose membranes. An equal loading of proteins was confirmed by staining with Ponceau-S solution [0.5% (w/v) Ponceau S in 1% (v/v) acetic acid]. 17 After blocking with 5% skim milk, the membranes were incubated with anti-PKCε antibody (Upstate, Temecula, CA) at 4°C overnight. The membrane was subsequently washed and incubated with peroxidase-conjugated secondary antibody (1:2,000, Cell Signaling Technology, Beverly, MA) for 2 h at room temperature. Immunostained bands were analyzed quantitatively by densitometry using an ECLTM Western BlotSystem (Cell Signaling Technology, Beverly, MA). The intensity of the immunostained PKCε band was normalized to that of the sum of Ponceau S-stained bands in the sample. The ratio of the mitochondrial (m) PKCε level to the cytosolic (c) PKCε level indicated the extent of PKCε translocation.
Role of ROS in PKCε translocation
To investigate the role of ROS in DG-induced PKCε translocation, rats were treated intragastrically with α-tocopherol (dissolved in oil) at 400 μg/kg once daily 2 h prior to DG treatment (0.45 g/kg, twice daily for 14 days) in the morning. Mitochondrial and cytosolic fractions were prepared from heart tissues and measured for PKCε level as described above.
Biochemical analyses
LDH activity of coronary effluent was measured as described. 18 Aliquots (210 μL) of mitochondrial fractions were taken for measuring reduced glutathione (GSH) level by an enzymatic method described by Griffith. 19 Aliquots (250 μL) of mitochondrial fractions were taken for measuring malondialdehyde (MDA) level by using a HPLC method. 11
Statistical analysis
Data were analyzed by one-way analysis of variance (ANOVA). Post hoc multiple comparisons were done with the least significant difference test. p values < 0.05 were regarded as statistically significant.
Results
Long-term DG treatment protected against myocardial I/R injury in rats
Long-term treatment with the DG extract (0.09–0.45 g/kg, given orally twice daily for 14 days), did not affect the LDH leakage in isolated-perfused (i.e., non-I/R) rat hearts (Fig. 2). However, I/R challenge caused a 9.3-fold increase in the extent of LDH leakage when compared with the non-I/R hearts. DG pretreatment dose-dependently reduced the extent of I/R-induced LDH leakage, with the degree of protection ranging from 13% to 40% at the doses tested when compared with animals subjected to I/R in the absence of DG pretreatment (i.e., DG untreated I/R hearts).
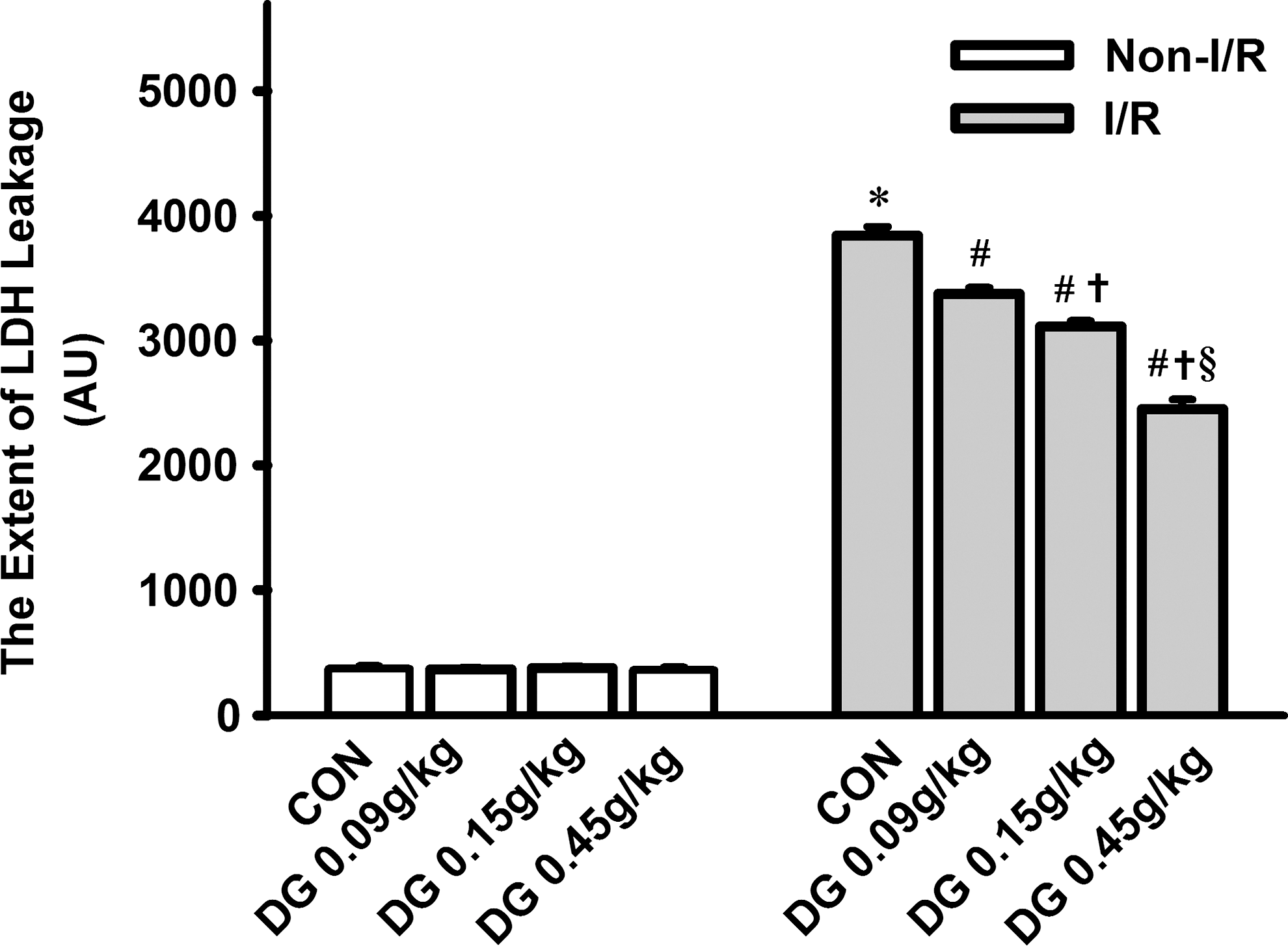
Effect of long-term Danshen-Gegen (DG) treatment on myocardial ischemia/reperfusion injury in rats. Animals were administered the DG extract orally twice daily for 14 days at the indicated doses. Control animals received the vehicle (i.e., water) only. Twelve hours after the last dosing, isolated-perfused hearts were subjected to 40 min of ischemia followed by 20 min of reperfusion. The extent of myocardial I/R injury was assessed by LDH leakage, as described in Materials and Methods, and data were expressed in arbitrary units (AU). Values given are means ± standard error of the mean (SEM), with n = 6. (*) Significantly different from the non-I/R control (CON) group; (#) significantly different from the I/R CON group; (†) significantly different from the respective DG 0.09 g/kg group; (§) significantly different from the respective DG 0.15 g/kg group.
I/R challenge also caused a significant decrease in mitochondrial GSH levels (15%) and an increase in ROS production (36%) in rat hearts, when compared with the non-I/R hearts (Fig. 3). Whereas long-term DG treatment dose-dependently increased mitochondrial GSH levels (11–29%) and decreased ROS production (4–17%) in non-I/R hearts, the cardioprotection afforded by DG pretreatment was associated with an increase in mitochondrial GSH levels (8–22%) and a decrease in ROS production (6–15%), when compared with DG untreated I/R hearts (Fig. 3).

Effects of long-term Danshen-Gegen (DG) treatment on mitochondrial antioxidant status in control and ischemic/reperfused (I/R) rat hearts. Animals were treated with the DG extract and isolated hearts were subjected to I/R challenge as described in Fig. 2. Mitochondrial antioxidant status was assessed by measurements of reduced glutathione (GSH) level and reactive oxygen species (ROS) production. Values given are means ± standard error of the mean (SEM), with n = 6. (*) Significantly different from the non-I/R control (CON) group; (#) significantly different from the I/R CON group; (†) significantly different from the respective DG 0.09 g/kg group; (§) significantly different from the respective DG 0.15 g/kg group.
I/R challenge significantly increased mitochondrial MDA production (32%), Ca2+ content (24%) and cytochrome c release (38%) in rat hearts when compared with the non-I/R hearts (Fig. 4). In non-I/R hearts, long-term DG treatment dose-dependently decreased mitochondrial MDA production (8–18%), Ca2+ content (7–24%), and cytochrome c release (6–20%) (Fig. 4); the cardioprotection against I/R injury afforded by DG pretreatment was also accompanied by decreases in the MDA production (5–14%), Ca2+ content (6–19%), and cytochrome c release (7–20%), when compared with the DG untreated I/R hearts.
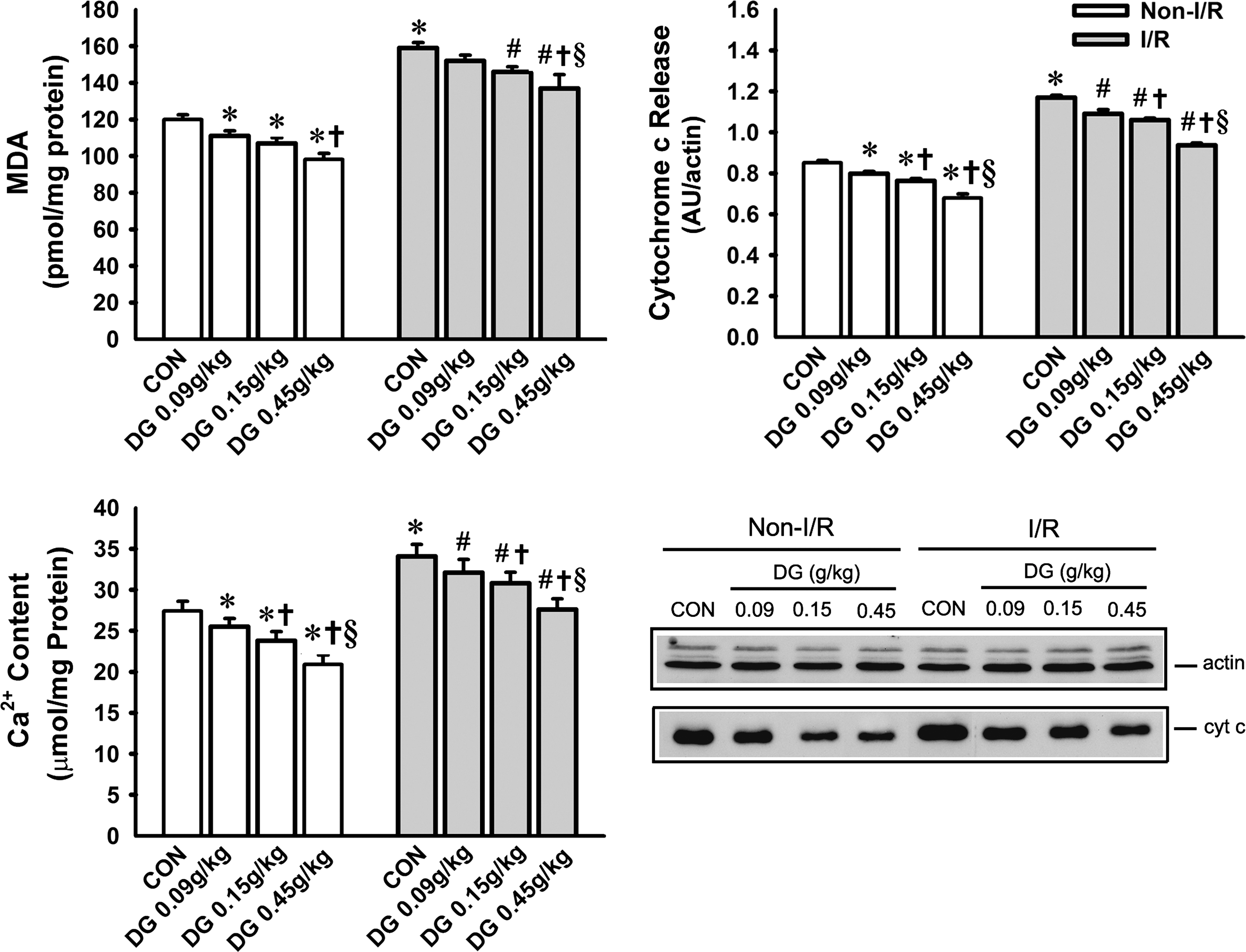
Effect of long-term Danshen-Gegen (DG) treatment on mitochondrial malondialdehyde (MDA) production, Ca2+ content, and cytochrome c release in control and ischemic/reperfused (I/R) rat hearts. Animals were treated with the DG extract and isolated hearts were subjected to I/R challenge as described in Fig. 2. Mitochondrial structural integrity was assessed by measurements of MDA production, Ca2+ content, and cytochrome c release, as described in Materials and Methods. The lower right panel shows the immunostained protein bands resulting from western blot analysis of cytosolic cytochrome c from representative samples. Values given are means ± standard error of the mean (SEM), with n = 6. (*) Significantly different from the non-I/R control (CON) group; (#) significantly different from the I/R CON group; (†) significantly different from the respective DG 0.09 g/kg group; (§) significantly different from the respective DG 0.15 g/kg group.
Long-term DG treatment decreased the sensitivity to Ca2+-induced MPT in control and ischemic/reperfused rat hearts
Ca2+-induced MPT was indirectly assessed by the in vitro measurement of mitochondrial swelling. I/R challenge increased the sensitivity of mitochondria to Ca2+-induced permeability transition (PT), as indicated by a significant increase in the extent of mitochondrial swelling (22%), when compared with the non-I/R hearts (Fig. 5). Whereas long-term DG treatment decreased the sensitivity of mitochondria to Ca2+-induced PT in non-I/R hearts (7–15%), the cardioprotection against I/R afforded by DG pretreatment was associated with protection against the alteration in mitochondrial sensitivity to Ca2+-induced PT (4–13%), when compared with DG untreated I/R hearts.

Effect of long-term Danshen-Gegen (DG) treatment on Ca2+-induced mitochondrial permeability transition in control and ischemic/reperfused (I/R) rat hearts. Animals were treated with the DG extract, and isolated hearts were subjected to I/R challenge as described in Fig. 2. Mitochondrial swelling was taken as an indirect measure of mitochondrial permeability transition (MPT). (
PKCε and mKATP inhibition suppressed DG-induced cardioprotection against I/R injury
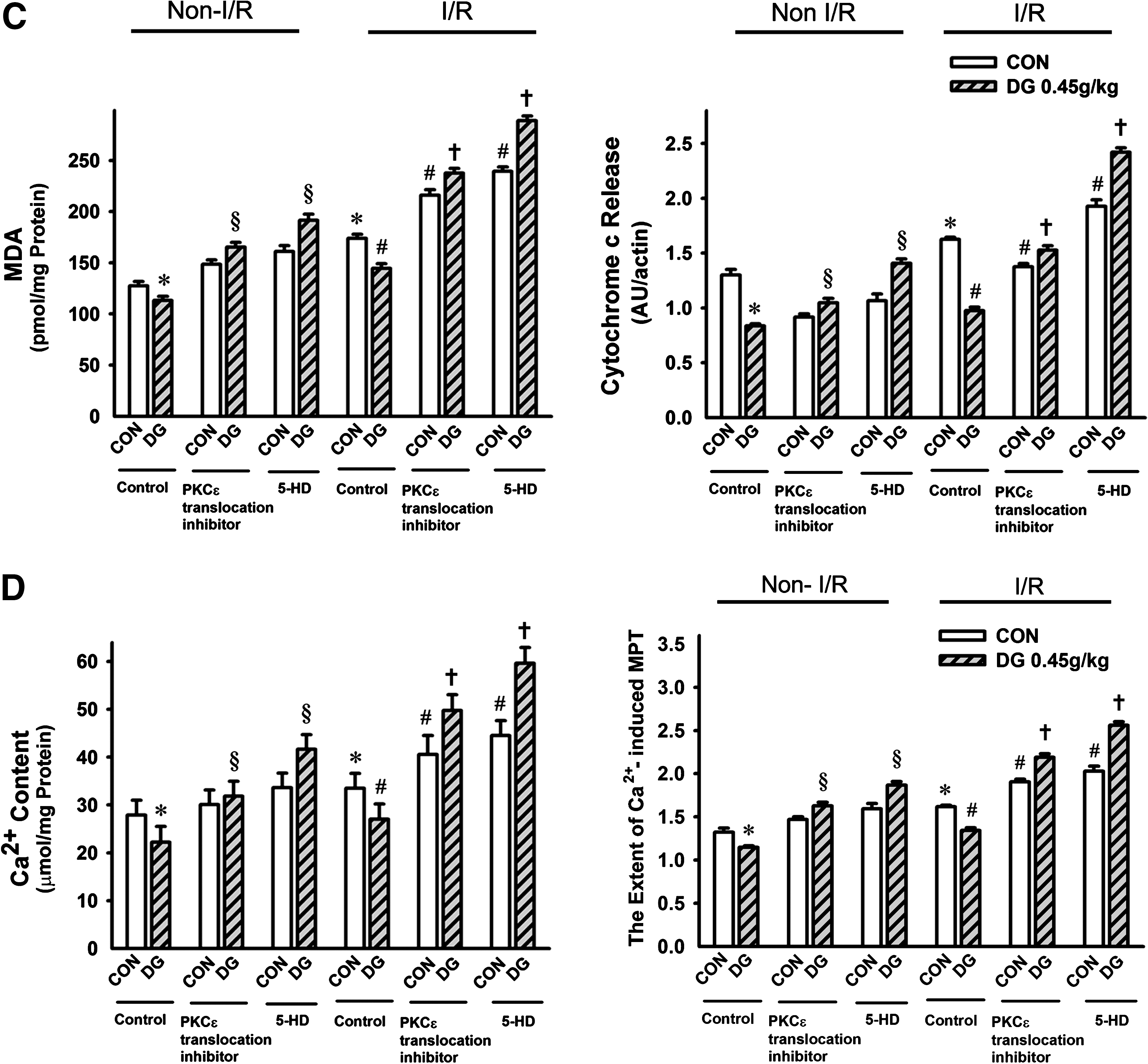
The involvement of a PKCε-mediated signaling pathway in the cardioprotective action of DG was investigated using agents known to act as inhibitors of PKCε translocation and of mKATP. The susceptibility of rat hearts to I/R injury, as assessed by LDH leakage, was increased by inhibition of PKCε or mKATP, but DG pretreatment (0.45 g/kg twice daily for 14 days) did not protect against myocardial I/R injury in the presence of PKCε or mKATP inhibition (Fig. 6a). The abolition of DG-induced cardioprotection was paralleled by the lack of improvement in the I/R-induced changes in mitochondrial GSH levels and ROS production (Fig. 6b), MDA production, Ca2+ content, cytochrome c release (Fig. 6c), or the sensitivity to Ca2+-induced PT (Fig. 6d).
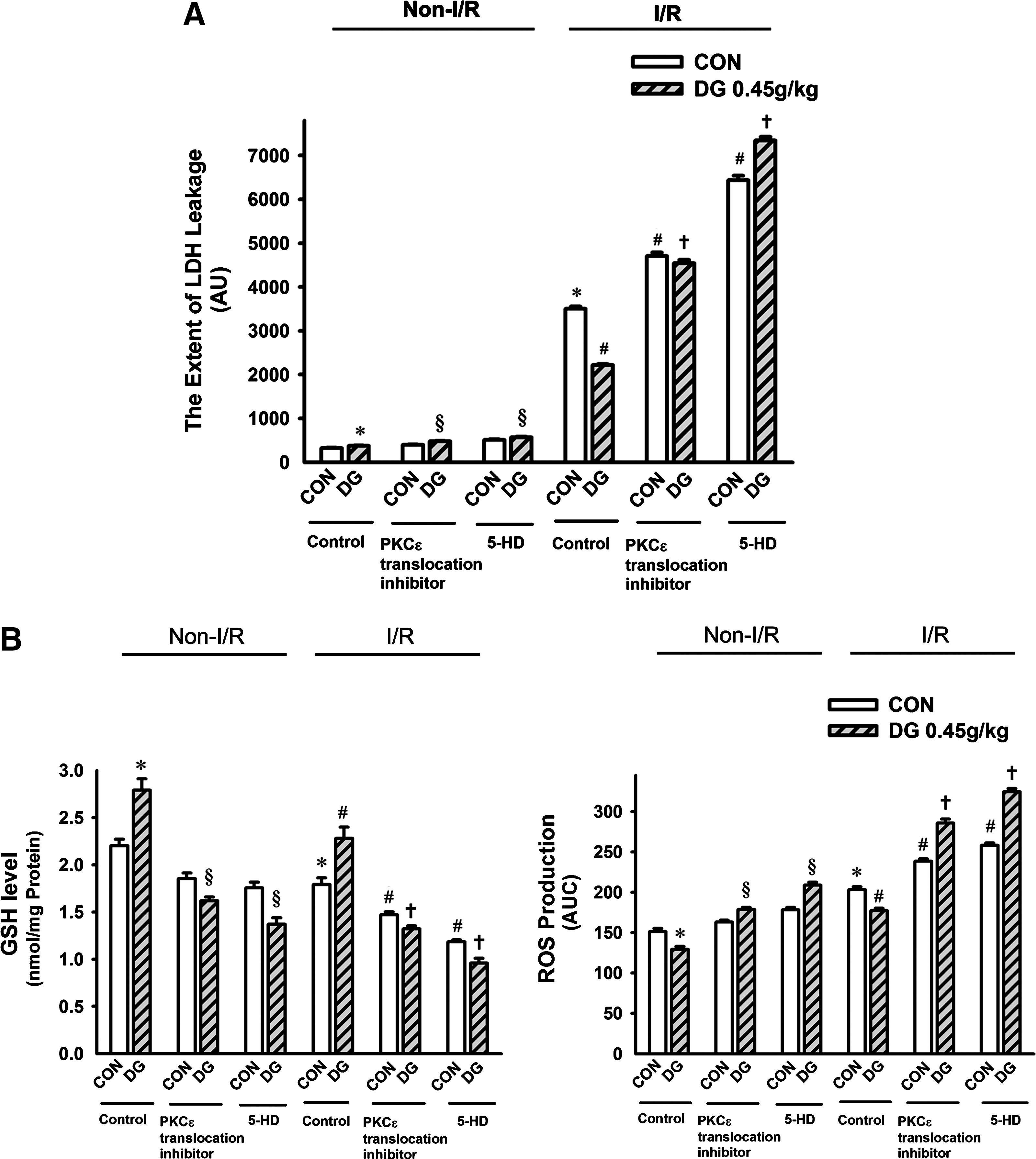
Effects of protein kinase C-epsilon (PKCε and mKATP inhibition on Danshen-Gegen (DG) afforded cardioprotection against ischemic/reperfusion (I/R) injury in rats. Animals were administered the DG extract orally twice daily at a dose of 0.45 g/kg for 14 days. Twelve hours after the last dosing, PKCε translocation inhibitor (Peptide EAVSLKPT) and mKATP inhibitor (5-hydroxy decanoate, 5-HD) was administered to isolated hearts by perfusion prior to the I/R challenge, as described in Materials and Methods. Isolated hearts were then subjected to 40 min of ischemia followed by 20 min of reperfusion, and the extent of I/R injury (
Long-term DG treatment activated PKCε translocation in rat hearts
The ratio of mitochondrial PKCε to cytosolic PKCε levels was taken as an indirect measure of PKCε translocation from the cytosol to mitochondria. DG pretreatment (0.45 g/kg twice daily for 14 days) enhanced PKCε translocation in rat hearts, as evidenced by a significant increase (73%) in the ratio of mPKCε to cPKCε, when compared with DG untreated hearts (Fig. 7).
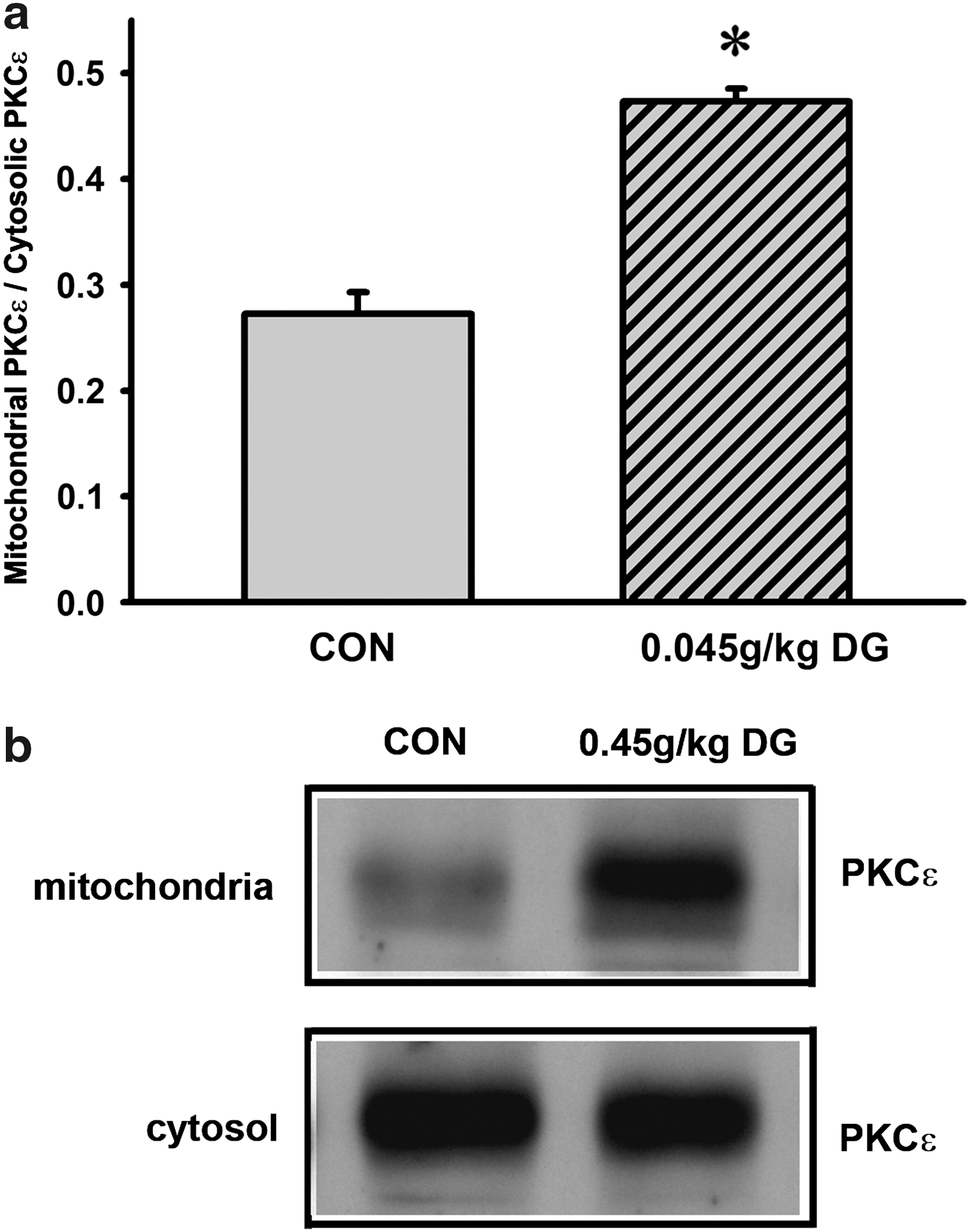
Effect of long-term Danshen-Gegen DG treatment on protein kinase C-epsilon (PKCε) translocation in rat hearts. (
Co-treatment with α-tocopherol (400 μg/kg) and the DG extract did not produce any detectable change in PKCε translocation in the hearts of control rats, the combined treatment completely suppressed PKCε translocation in the long-term DG-treated rats (Fig. 8).
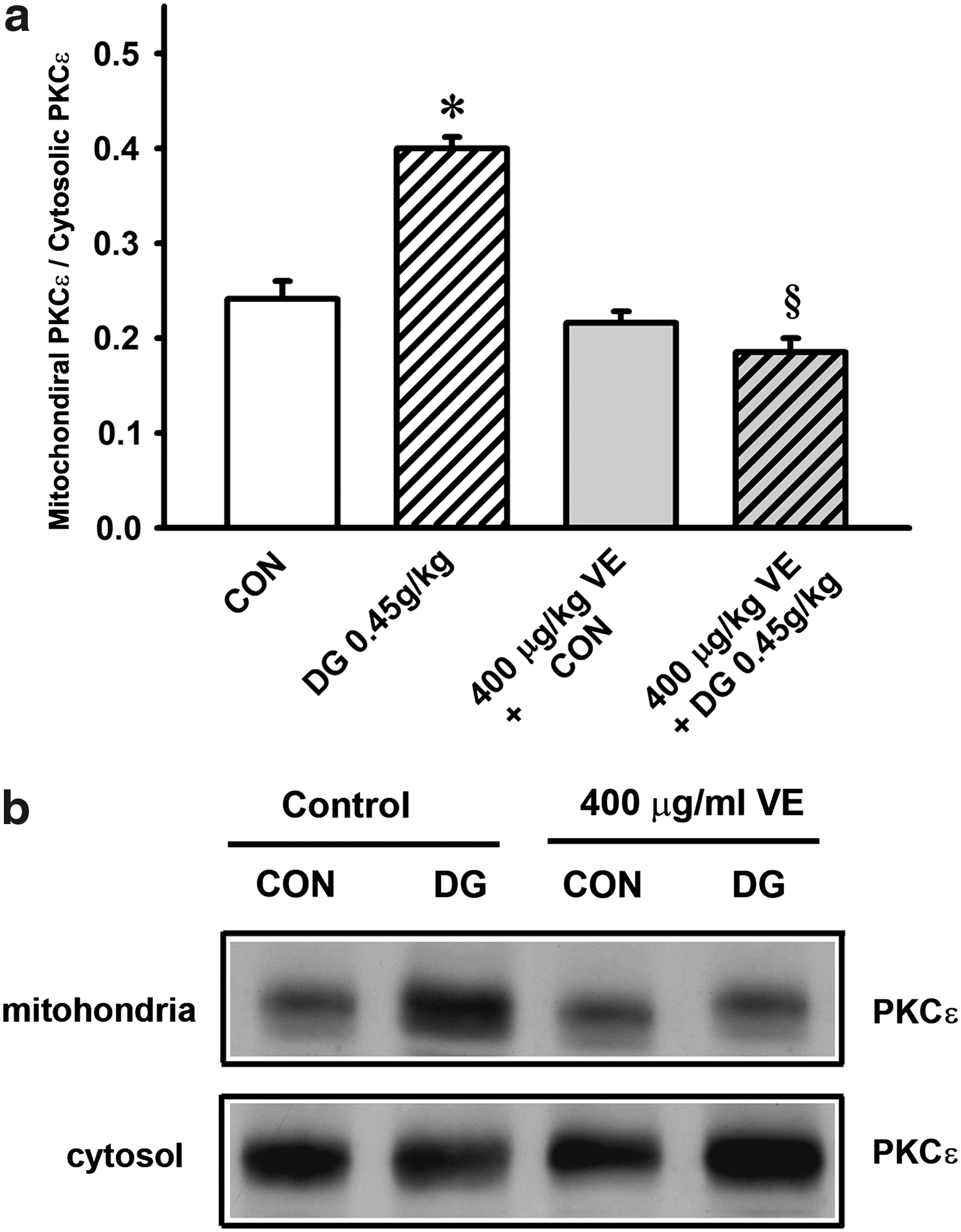
Effect of α-tocopherol co-treatment on Danshen-Gegen (DG)-induced protein kinase C-epsilon (PKCε translocation in rat hearts. (
Discussion
I/R challenge caused tissue damage in isolated perfused rat hearts as assessed by LDH leakage. It has been shown that the I/R-induced increase in the extent of LDH leakage positively correlates with histopathological changes in ventricular tissue in isolated-perfused rat hearts. 20 Therefore, the measurement of LDH leakage can be used as a reliable quantitative measure of myocardial injury caused by I/R challenge. Under the present experimental conditions, the myocardial injury caused by the ischemic and/or reperfusion insult was assessed by LDH leakage during the 20-min period of postischemic reperfusion. It is well established that ROS-mediated processes are involved in the pathogenesis of myocardial I/R injury, 21 and mitochondria are an important source of ROS production during postischemic reperfusion. 22 Consistent with this, mitochondria isolated from ischemic/reperfused rat hearts were found to have decreased GSH levels and increased ROS and MDA production, indicating an increase in oxidative stress.
Mitochondria, which are essential for maintaining cellular structural and functional integrity, serve as coordinators of cell survival and death. 23 In this regard, the opening of MPT pores is critically involved in cellular dysfunction and cell death. 24 In the present study, myocardial injury induced by I/R was associated with an increased sensitivity of mitochondria to Ca2+-induced PT. The opening of MPT pores, either under in vivo or in vitro conditions, is triggered by stimuli such as oxidants, high mitochondrial Ca2+ content, and/or depletion of adenine nucleotides. 25 The disruption of mitochondrial membrane structural integrity associated with I/R, as indicated by an increase in cytochrome c release, is paralleled by the increase in mitochondrial sensitivity to Ca2+-induced PT. Long-term treatment with DG at increasing doses (including an equivalent of a human dose adopted for clinical investigation) protected against myocardial I/R injury. This cardioprotection was paralleled by a partial protection against I/R-induced impairment in mitochondrial antioxidant and membrane structural status, GSH levels, and sensitivity to Ca2+-induced PT.
GSH plays an important role in numerous cellular functions, including the regulation of Ca2+ homeostasis and the detoxification of ROS. 26,27 The decrease in mitochondrial sensitivity to Ca2+-stimulated PT by DG, as observed in the present study, may be related to the enhancement of mitochondrial glutathione redox status, which is characterized by the increase in the level of GSH. In this regard, one of the two voltage-sensitive sites of the MPT pore was found to be gated by a critical dithiol that is sensitive to glutathione redox status, and a decrease in GSH levels as a result of oxidation could open this pore. 28,29 Furthermore, the modification of a specific thiol group on the adenine nucleotide translocase molecule, either by oxidative stress or thiol-selective reagents, also has been shown to decrease adenine nucleotide binding and activate the MPT pore. 30 Increased oxidative stress enhances cyclophilin binding and thereby increases the sensitivity of MPT. 30 The observation that a smaller degree of decrease in sensitivity to Ca2+-induced MPT than that of myocardial I/R injury afforded by long-term DG treatment supports the notion that the decrease in the sensitivity of mitochondria to PT is an effect secondary to the enhancement of mitochondrial glutathione redox status. The direct effect on increasing mitochondrial GSH level and/or the indirect effect on removing mitochondrial-derived ROS produced by DG pretreatment might well increase the threshold for MPT pore opening in the presence of Ca2+. 25
I/R challenge caused an increase in mitochondrial Ca2+ content in rat hearts. It has been shown that cytosolic Ca2+ content increases during myocardial I/R, 31 leading to the accumulation of Ca2+ in mitochondria via uptake by the inner-membrane Ca2+ uniporter. 32,33 The resulting mitochondrial Ca2+ overload not only generates energy-consuming futile cycles that divert the use of the inner membrane proton gradient to cation transport rather than ATP production, 33 but also predisposes the mitochondria to undergo a permeability transition. 34 MPT further jeopardizes the cellular energy status, and the consequent loss of ion homeostasis can lead to necrotic cell death. The release of cytochrome c from the mitochondrial inner membrane, an event that is believed to be secondary to the onset of MPT, 35 is a key step leading to apoptosis. 36
Although the relative contribution of necrotic and apoptotic cell death to I/R-induced tissue injury was not determined in the present study, our finding that mitochondria isolated from DG-pretreated hearts show a decreased sensitivity of to Ca2+-stimulated PT suggests that the increase in mitochondrial resistance to this transition may play an important role in protecting against myocardial I/R injury. It has been reported that myocardial preconditioning by tanshinone II A or puerarin from Danshen and Gegen, respectively, confers cardioprotection through the inhibition of MPT or opening of mKATP channels. 37,38 The observation that long-term DG treatment decreased mitochondrial Ca2+ levels in both non-I/R and I/R hearts may be related to the enhancement of mitochondrial GSH content by DG. 26
The cardioprotection against I/R injury afforded by DG pretreatment was abrogated by PKCε or mKATP inhibition, suggesting the involvement of PKCε activation and mKATP channel opening in the process of myocardial preconditioning by DG. PKCε is a member of a novel group of the PKC family of serine and threonine kinases that are involved in a wide range of physiological processes, including mitogenesis, cell survival under stressful conditions, metastasis, and transcriptional regulation. 39 In this connection, activation of PKCε prior to ischemia has been demonstrated to afford cardioprotection by mimicking ischemic preconditioning. 40
The role of mKATP in ischemic preconditioning is not well understood. 41 The prolongation of onset time for MPT by ischemic preconditioning was found to be associated with the translocation of PKCε from the cytoplasm to mitochondria in isolated adult rat cardiomyocytes. 42 Recently, it has been shown that phosphorylation by PKCε1 can open mKATP channels, which in turn increases intramitochondrial ROS production and the subsequent activation of PKCε2. 43 PKCε2 can inhibit MPT directly by phosphorylation of its component proteins, thereby reducing the extent of necrotic and/or apoptotic cell death in the myocardium during reperfusion. 43 Consistent with these observations, our studies showed that long-term DG treatment increased the level of phosphorylated PKCε (Ser729) in rat myocardium (data not shown), and the associated cardioprotection was abolished by the inhibition of PKCε translocation or of mKATP channels. In addition, our findings showed that long-term DG treatment enhanced PKCε translocation and this DG-induced PKCε translocation was abrogated by α-tocopherol co-treatment. In this connection, the translocation of PKCε has been shown to be triggered by ROS-mediated processes. 42 Conceivably, ROS generated from the biotransformation of DG-derived ingredients in cardiomyocytes, as observed in cultured cardiomyocytes (data not shown), may play a role in the cardioprotection afforded by DG pretreatment. In addition to the activation of PKCε translocation, the enhancement in mitochondrial GSH levels in DG-pretreated hearts may be related to a redox-sensitive and nuclear factor-erythroid derived 2 (NF-E2)–related factor 2-mediated antioxidant response. 44 Both cellular responses elicited by DG pretreatment precondition the myocardium and render it more resistant to I/R injury.
Long-term DG treatment at low doses, as was the case in the present study, likely preconditions the myocardium pharmacologically rather than exerting a direct action in attenuating the I/R injury. This postulation is supported by the observation that the oral administration of the DG extract at the same low doses did not produce any cardioprotective effect 24 h after the dosing in rats (data not shown). Consistent with this, long-term oral treatment with salvianolic acid B (an active ingredient present in the DG extract, 400 mg/kg × 4 weeks) protected against myocardial infarction in rats, partly by augmenting the expression of vascular endothelium growth factor and promoting angiogensis. 45 However, acute treatment with puerarin (0.24 mmol/L in perfusate for 5 min) or daidzein (10 mg/kg, intraperitoneally [i.p.]) prior to ischemia also conferred cardioprotection against I/R injury in rats both in vitro and in vivo by opening the calcium-activated potassium channel and activating PKC or inhibiting nuclear factor-κB (NF-κB) activation, respectively. 46,47
Interestingly, intravenous administration of a mixture of puerarin and danshensu prior to an ischemic insult was also found to protect against myocardial I/R injury in rats through antioxidant action. 48 In contrast to the myocardial protective effects of long-term treatment with the DG extract at relatively low doses, which are associated with preconditioning, acute treatment with high doses may exert direct cardioprotective effects, as was the case for puerarin and daidzein. A recent study from our laboratory has indicated that the oral administration of the DG extract at a dose of 4 g/kg immediately after the intraperitoneal injection of isoproterenol protected against isoproterenol-induced myocardial injury in rats (unpublished data). Conceivably, synergistic interactions among multiple active ingredients present in the DG extract may produce cardioprotection against oxidative challenge through an indirect myocardial preconditioning action or a direct cardiac action on specific target(s). Taken together, active ingredients from the DG extract may act directly (via opening calcium-activated potassium channel and inhibiting NF-κB activation) and/or indirectly (via enhancing mitochondrial glutathione antioxidant status, activating PKCε, opening mKATP and inhibiting MPT) in protecting the myocardium against I/R injury.
In conclusion, long-term DG treatment caused dose-dependent protection against myocardial I/R injury in rats. The cardioprotection afforded by DG pretreatment was associated with enhancements in mitochondrial antioxidant status and membrane structural integrity as well as a decrease in the sensitivity of mitochondria to Ca2+-stimulated PT in vitro, particularly under I/R conditions. Long-term DG treatment may precondition the myocardium via a redox-sensitive PKCε/mKATP pathway, with resultant inhibition of MPT. Our results suggest that clinical studies examining the effectiveness of DG extract given prophylactically in affording protection against myocardial I/R injury would be warranted.
Footnotes
Acknowledgments
The work described in this paper (or equipment/facility) was fully/substantially/partially supported by a grant from the University Grants Committee of the Hong Kong Special Administrative Region, China (Project No. AoE/B-10/01).