
Abstract
Aging is associated with a progressive decline of skeletal muscle function and insulin sensitivity. Sex differences in the insulin response to different physiological situations have been found, leading to the development of type 2 diabetes. The aim of this study was to investigate the changes in insulin sensitivity with age in male and female rats and to elucidate whether there are sex differences in the alteration profiles of systemic insulin sensitivity parameters, adiposity, skeletal muscle oxidative damage, and the insulin signaling pathway. The gastrocnemius and soleus muscles of male and female rats of 3, 9, and 18 months of age were used. The decrease of insulin sensitivity with age was higher in female than in male rats. However, the increase of both serum insulin levels and adiposity with age shows a different profile in both sexes and suggests an earlier onset of age-related impairment of insulin sensitivity in male than in female rats. Sex differences in insulin signaling key protein levels were found mainly in the most aged rats, suggesting that sex differences in these proteins would be manifested at more advanced ages than differences in the insulin-sensitivity serum profile. In addition, the gastrocnemius muscle showed more age-associated oxidative damage and insulin resistance impact than the soleus in both sexes. These results suggest the sex differences found in the impairment of insulin sensitivity of aged rats would not be attributable to differences between sexes in the time course of the levels of key proteins of the skeletal muscle insulin signaling pathway, at least in the first 18 months of life.
Introduction
Insulin binds to its receptor, which belongs to a family of tyrosine kinase receptors and provokes the phosphorylation of insulin receptor substrate 1 (IRS-1) and, subsequently, the protein kinase B (Akt), finally resulting in the activation of glucose transporter 4 (GLUT4) translocation and glucose uptake. 3 It has been shown that insulin action is greater in oxidative than in glycolytic skeletal muscles. 4,5
Aging is associated with a progressive decline of muscle mass and function due to a reduction in the size and number of muscle fibers. 6 This condition is accompanied by a decrease of insulin sensitivity that has been attributed to chronological age itself and/or to a variety of secondary factors associated with the aging process, such as an increase in body fat and/or in central adiposity, and a reduction in spontaneous physical activity. 7,8 The decline of insulin action occurs through an impairment of the insulin signaling pathway, 9 leading to the development of type 2 diabetes.
Although the underlying mechanism for age-related insulin resistance is not completely elucidated, increased oxidative stress and decreased antioxidant defenses are likely to be involved in the impairment of muscle insulin signaling. 4,10,11 Moreover, the increase of adiposity associated with aging is accompanied by changes in the levels of circulating adipokines, some of which are negative regulators of insulin signaling contributing to the development of insulin resistance. 12 Most studies in rats on aging-associated impairment of skeletal insulin sensitivity have shown a decline of the insulin signaling pathway beyond 24 months of age, 4,9,13 and some evidence points to an effect at earlier ages, at 12 14 and 20 15 months. However, the time point that may be considered the beginning of this impairment remains unclear and whether it is different between sexes. Thus, we selected male and female rats of both sexes and of three groups of age (3, 9, and 18 months) to investigate the time course of the alterations of insulin sensitivity found in aged rats (over 30 months of age).
Previous studies in our laboratory showed sex differences in mitochondrial oxidative function and oxidative stress in different tissues of rat, such as liver, 16 –18 brown adipose tissue, 19,20 brain, 21 pancreas (unpublished results), and skeletal muscle, 22 –24 in which female rats showed a higher mitochondrial oxidative capacity and a more effective antioxidant protection than males. Moreover, female rats have been reported to be protected from lipid-induced reductions in insulin action compared to male rats 25 as well as to show a better response of skeletal muscle insulin sensitivity to high-fat diet feeding than males. 24 In addition, sex-related differences in the pattern of muscle mass loss with age have been described. 26 However, as far as we know, there are no studies comparing the progression of skeletal muscle insulin sensitivity between sexes that would allow us to understand the molecular mechanisms underlying the different incidence of type 2 diabetes found in humans. 27
The aim of this study was to investigate whether the differences between sexes found in the systemic insulin sensitivity of aged rats could be due to differences in the time course of the impairment of insulin signaling pathway. We hypothesized that the changes in insulin signaling key intermediates that occur with age may show a different profile in both sexes and in different muscle types. To test this hypothesis, we investigated the progression with age of the levels of key proteins of the insulin signaling pathway in two skeletal muscles with different metabolic characteristics, one oxidative (soleus) and another glycolytic (gastrocnemius), as well as the possible sex differences in the change over time of systemic insulin-sensitivity parameters, adiposity, and skeletal muscle oxidative damage in male and female rats. The understanding of the molecular mechanisms underlying the sex differences in the progression of insulin sensitivity with age may be useful in designing strategies to prevent the development of type 2 diabetes.
Methods
Animals and diets
Animal experiments were performed in accordance with the general guidelines approved by our institutional ethics committee and European Union (EU) regulations (86/609/CEE and 2003/65/CE). Eight male and 8 female Wistar rats (Charles River, Barcelona, Spain) 3, 9, and 18 months of age, respectively, were housed 2 per cage with free access to a pelleted diet (A04, Panlab, Barcelona) and water. They were kept at 22°C under a 12-h light–dark cycle. Rats were sacrificed by decapitation after a 12-h period of fasting. Blood was collected, and serum was separated by centrifugation at 900 × g for 20 min at 4°C. Adipose tissue depots were extracted, and the adiposity index was calculated from the sum of inguinal, gonadal, mesenteric, and retroperitoneal depot weights relative to 100 g of body weight.
Soleus and gastrocnemius skeletal muscles were dissected rapidly, weighed, frozen in liquid nitrogen, and stored at −80°C until processed. Pieces of muscle were homogenized at 4°C in a proportion of 1 g of muscle in 10 mL of buffer (50 mM HEPES, 100 mM NaF, 10 mM EDTA, 1 mM Na3VO4, 1% Triton X-100, 2 mM phenylmethylsulfonyl fluoride [PMSF], 10 μg/mL aprotinin, 10 μg/mL leupeptin, pH 7.4).
Materials
The Accutrend® glucose cholesterol triglyceride (GCT) meter and glucose test strips were supplied by Roche Diagnostics (Basel, Switzerland). Enzyme immunoassay kits were used for measurement of rat serum insulin (Mercodia, Uppsala, Sweden), and total and high-molecular-weight adiponectin (Phoenix Pharmaceuticals Inc., Belmont, CA, and Biovendor, Heidelberg, Germany, respectively). The OxyblotTM Protein Oxidation Detection kit and rabbit polyclonal antibodies to rat UCP3 (cat. no. AB4036) and GLUT4 (cat. no. sc-7938) were purchased from Chemicon International (Temecula, CA). Insulin receptor-β (IR-β) antibody (cat. no. 611277) was from BD Biosciences (San Jose, CA). IRS-1 (cat. no. 2382); Akt (cat. no. 9272) and phosphorylated-Akt (p-Akt, cat. no. 9271) antibodies were obtained from Cell Signaling Technology (Danvers, MA). The chemiluminescence kit (ECL) for immunoblot development was purchased from Bio-Rad (Hercules, CA). Routine chemicals were supplied by Pronadisa (Madrid, Spain), Panreac (Barcelona, Spain), and Sigma-Aldrich (St. Louis, MO).
Oral glucose tolerance test
The test was performed on the week before the sacrifice. Rats were fasted for 12 h and then glucose (2 g/kg body weight) was given orally. Blood was collected from the tail vein just before glucose administration and after 15, 30, 60, 90, and 120 min, and glucose concentrations were measured using the Accutrend® system.
Serum glucose, insulin, and adiponectin levels
Serum glucose levels were measured by using the Accutrend® system, and serum insulin and adiponectin levels were measured by enzyme immunoassay kits. Homeostasis model assessment (HOMA-IR) was used to estimate insulin resistance (IR) 28 and was calculated as [fasting glucose (mM) × fasting insulin (μU/mL)]/22.5.
Measurements of skeletal muscle thiobarbituric acid-reactive substances (TBARS) and protein carbonyl groups
TBARS levels were measured in sample homogenates as previously described 29 and used as an index of lipid peroxidation. Protein carbonyl groups were determined as an index of protein oxidation by dot-blot detection using the OxyBlotTM Protein Oxidation Detection kit according to the manufacturer's protocol with several modifications. 21
Western blot analysis
Homogenized samples were centrifuged for 20 min at 13,000 × g at 4°C, and supernatants were collected, as previously reported. 30 Then 50 μg of soleus or gastrocnemius muscle protein were fractionated on 12% or 10% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) gels and electrotransferred onto a nitrocellulose filter. At least one sample from each experimental group was loaded in every gel, and gels were run simultaneously. Rabbit polyclonal antibodies to rat IR-β, IRS-1, Akt, p-Akt, UCP3, and GLUT4 were used as primary antibodies. Anti-rabbit immunoglobulin G (IgG)-alkaline phosphatase antibody was used as a secondary antibody. Development of immunoblots was performed using an enhanced chemiluminescence kit. Bands were visualized with the ChemiDoc XRS system (Bio-Rad, Hercules, CA) and analyzed with the image analysis program Quantity One© (Bio-Rad, Hercules, CA). Bands revealed an apparent molecular mass of 92 kDa (IR-β), 180 kD (IRS-1), 60 kD (Akt and p-Akt), 34 kD (UCP3), and 45 kD (GLUT4).
Statistical analysis
All data are expressed as mean values ± standard error of the mean (SEM) of 8 animals per group. Statistical analyses were performed using a statistical software package (SPSS 17.0 for Windows, Inc., Chicago, IL). Statistical differences between experimental groups were analyzed by two-way analysis of variance (ANOVA) followed by least significance difference (LSD) post hoc test and the Student t-test. A p value of less than 0.05 was considered statistically significant.
Results
Body and skeletal muscle tissue weights and adiposity index
Body weight and adiposity index increased with age in both sexes (Table 1). Adiposity index was lower in 9-month-old female rats than in their male counterparts, but no differences between sexes at 3 and 18 months of age were found. Gastrocnemius muscle weight increased with age in both sexes and was higher in male rats than in females. Soleus muscle weight was not modified with age and was also higher in males. Soleus and gastrocnemius relative weights decreased with age in both sexes and, as a whole, were higher in female rats than in males.
Adiposity index is the sum of inguinal, gonadal, mesenteric, and retroperitoneal depot weights relative to 100 g of body weight. Values are expressed as the mean ± the standard error of the mean (SEM) of 8 animals per group. Analysis of variance (ANOVA) (p < 0.05): S indicates sex effect, A indicates aging effect, and S*A indicates sex and aging interactive effect. Post hoc analysis of age (least significant difference [LSD] test; p < 0.05): a indicates 9- and 18-month-old rats versus 3-month-old rats; b indicates 18-month-old rats versus 9 month-old rats. Student t-test (p < 0.05): c indicates female rats versus male rats.
Serum glucose, insulin and adiponectin levels
Serum glucose and insulin levels and HOMA-IR values were higher in male rats than in females (Table 2) and increased with age in both sexes. When serum glucose and insulin levels were expressed as a percentage with respect to the values shown by 3-month-old animals (Fig. 1), the higher increase of serum insulin was observed between 3 and 9 months of age in males (2.3 times), but between 9 and 18 months of age in females (2.2 times), with both sexes reaching a similar percentage increase at 18 months. Both sexes showed a significant increase of glycemia at 18 months of age. Serum adiponectin levels (Table 2) rose in males between 3 and 9 months but dropped in both sexes at 18 months of age.
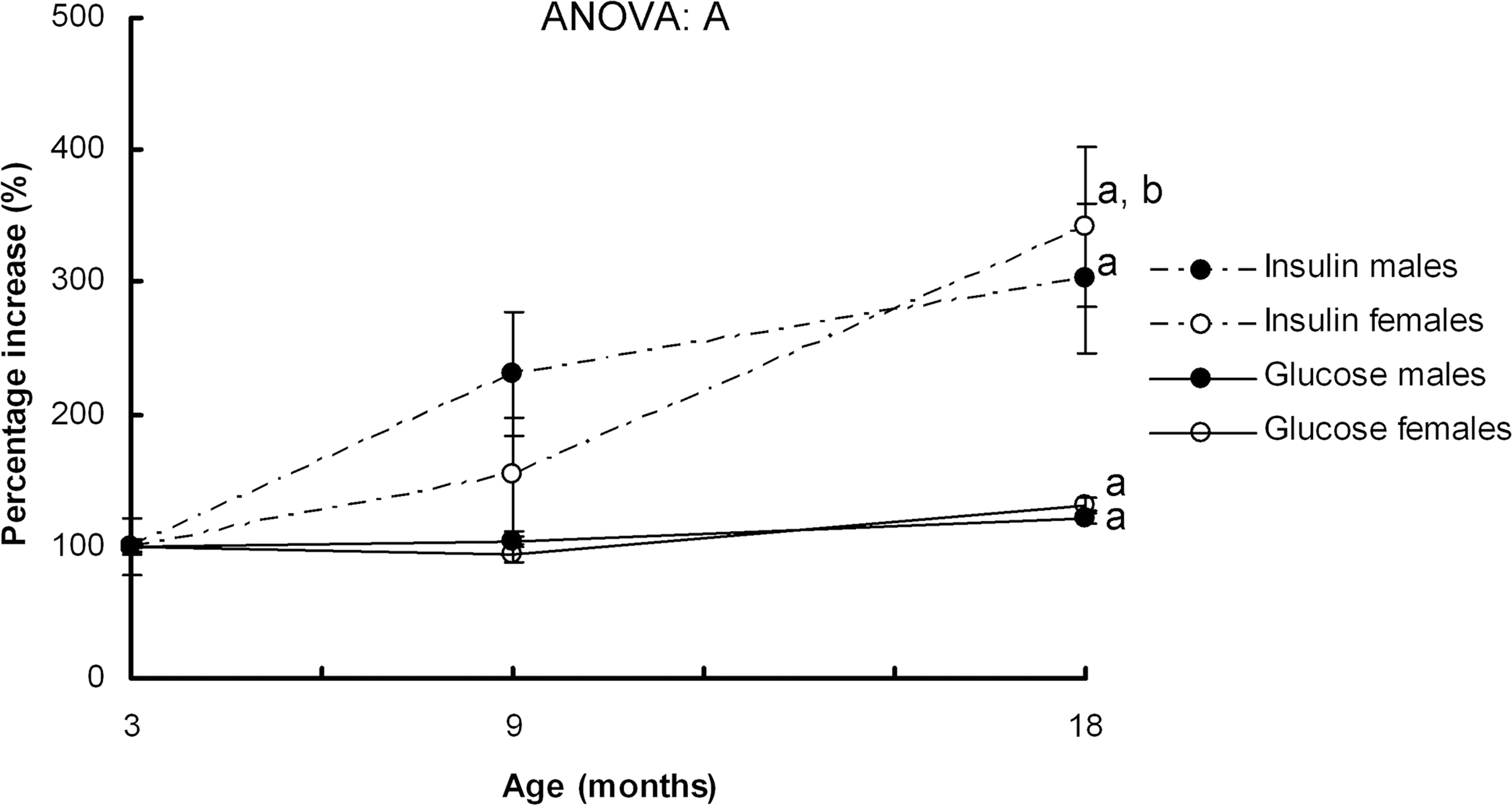
Serum insulin and glucose increase with age. Values are expressed as percentages and the levels of 3-month-old male rats were set as 100%. Data are the mean ± standard error of the mean (SEM) of 8 animals per group. Analysis of variance (ANOVA) (p < 0.05): A indicates aging effect. Post hoc analysis of age (least significance difference [LSD] test; p < 0.05).
Homeostasis model assesment of insulin resistance (HOMA-IR) was used to estimate insulin resistance and was calculated as [fasting glucose (mM) × fasting insulin (μU/mL)]/22.5. Values are expressed as the mean ± standard error of the mean (SEM) of 8 animals per group. Analysis of variance (ANOVA) (p < 0.05): S indicates sex effect and A indicates aging effect. Post hoc analysis of age (least significant difference [LSD] test; p < 0.05): a indicates 9- and 18-month-old rats versus 3-month-old rats, b indicates 18-month-old rats versus 9-month-old rats. Student t-test (p < 0.05): c indicates female rats versus male rats.
Oral glucose tolerance test
In both sexes, the time-course glucose levels increased at 18 months of age (Fig. 2), showing an impaired profile of glucose tolerance compared to rats of 3 and 9 months of age.
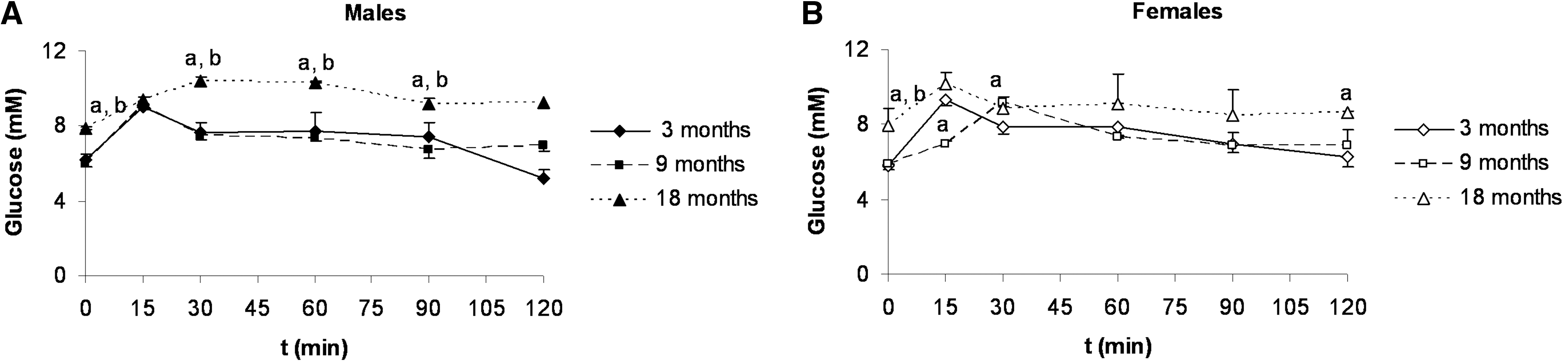
Oral glucose tolerance curves.
Skeletal muscle oxidative damage and UCP3 levels
In gastrocnemius muscle, 18-month-old male rats showed higher TBARS levels and protein carbonyl groups than their female counterparts (Table 3), but no differences between sexes at 3 and 9 months of age were found. Age increased gastrocnemius TBARS levels and protein carbonyl groups in both sexes. Gastrocnemius UCP3 protein levels were higher in female rats than in males, although the sex effect did not reach statistical significance (p = 0.051).
Values of protein carbonyl groups and UCP3 protein levels are expressed as %AU (arbitrary units), where levels of 3-month-old male rats were set as 100%. Data are the mean ± standard error of the mean (SEM) of 8 animals per group. Analysis of variance (ANOVA) (p < 0.05): S indicates sex effect, A indicates aging effect, and S*A indicates sex and aging interactive effect. Post hoc analysis of age Least significant difference [LSD] test, p < 0.05): a indicates 9- and 18-month-old rats versus 3-month-old rats, b indicates 18-month-old rats versus 9-month-old rats. Student t-test (p < 0.05): c indicates female rats versus male rats.
TBARS, thiobarbituric acid-reactive substances; N.S., not significant.
In soleus, male rats of 3 and 18 months of age showed higher protein carbonyl group levels than their female counterparts. In male rats, soleus muscle protein carbonyl levels increased at the age of 18 months, whereas in females they increased earlier, at the age of 9 months, and decreased at 18 months, reaching 3-month-old rat levels. No significant sex or age effects were found in soleus TBARS or UCP3 protein levels.
Skeletal muscle insulin signaling key protein levels
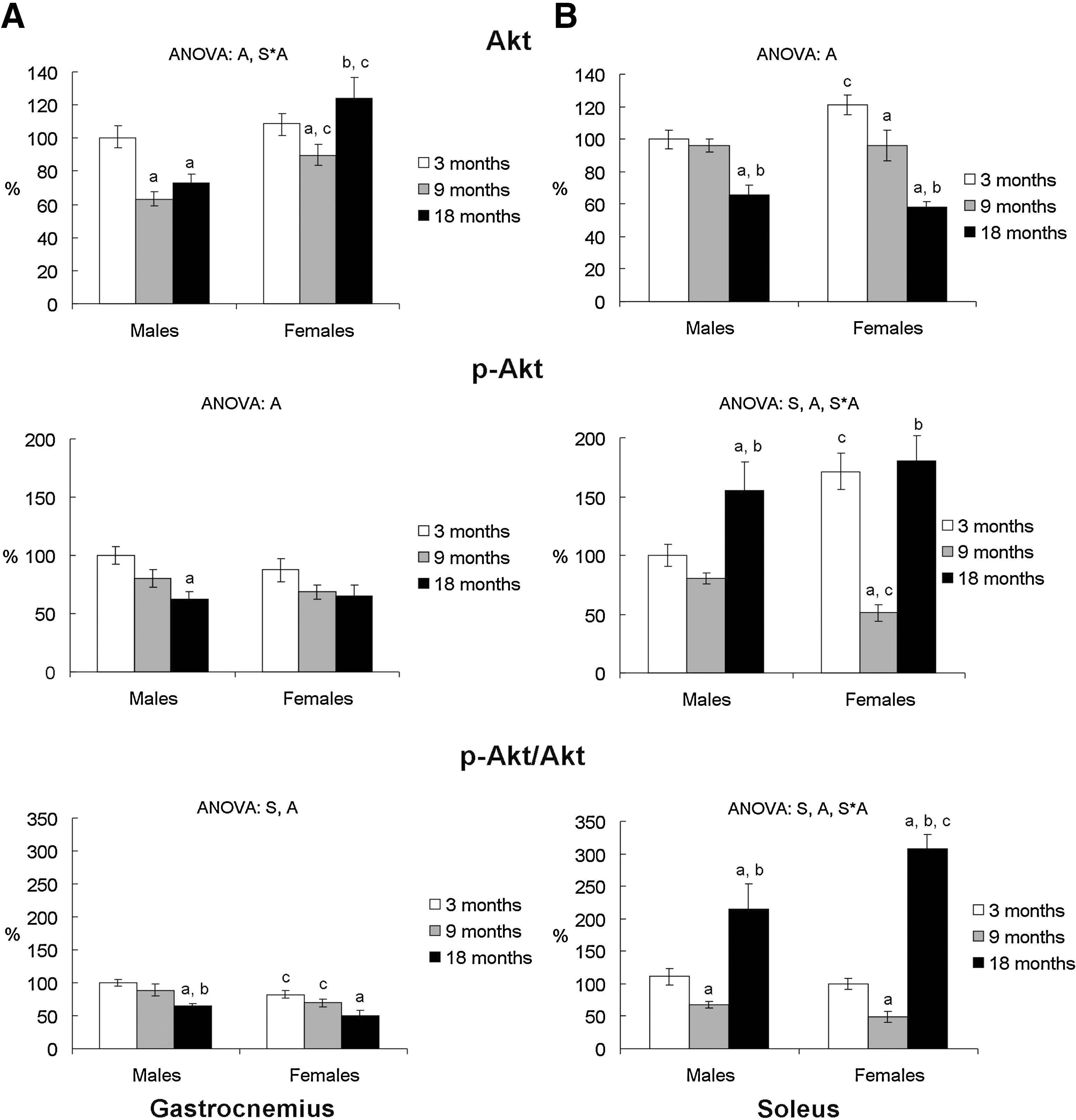
Gastrocnemius IR-β protein levels were higher in 3-month-old female rats compared to males and decreased with age, whereas male rats showed an age-related increase at 18 months (Fig. 3a). In soleus, age decreased IR-β protein levels in 18-month-old rats of both sexes (Fig. 3b). Gastrocnemius and soleus IRS-1 protein levels decreased with age in both sexes (Fig. 3). In male rats, gastrocnemius IRS-1 protein levels decreased at 9 months of age (Fig. 3a) and soleus IRS-1 at 18 months (Fig. 3b), whereas in females IRS-1 protein levels decreased at 9 months of age in both muscles. Gastrocnemius Akt protein levels were higher in 9- and 18-month-old female rats compared to their male counterparts (Fig. 4a), whereas the pAkt/Akt ratio was higher in males than in females at 3 and 9 months of age. Soleus Akt protein levels were higher in 3-month-old female rats than in their male counterparts (Fig. 4b). In both sexes, total Akt decreased at 18 months of age, whereas p-Akt and the p-Akt/Akt ratio decreased in 9-month-old rats and then increased in 18-month-old rats, with 18 month-old female rats showing a higher ratio value than their male counterparts. Three-month-old female rats showed lower gastrocnemius GLUT4 protein levels than males (Fig. 3a). GLUT4 levels decreased with aging in both sexes. No significant sex or age effects were found in soleus GLUT4 protein levels (Fig. 3b).
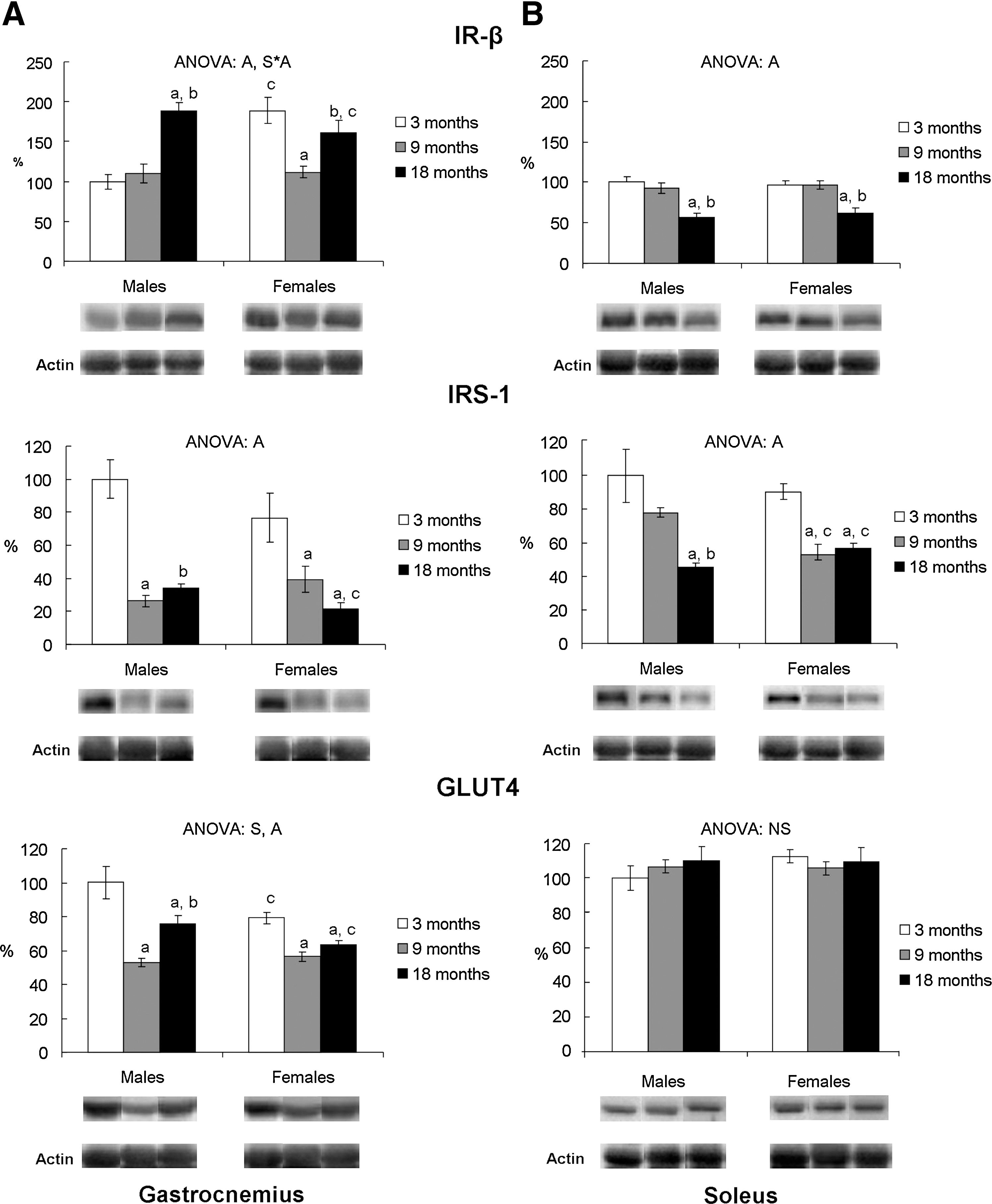
Gastrocnemius (
Gastrocnemius (
Discussion
Age implies a loss of insulin sensitivity that leads to an increase of serum insulin levels to maintain glucose homeostasis. We have found notable sex differences in the profile of the increase of serum insulin levels that occurs with age. Thus, at 9 months of age, male rats need to increase their insulin levels more than females to maintain glycemia, whereas in female rats this increase occurs at 18 months of age. These results suggest that the age-associated impairment of insulin sensitivity may develop earlier in male rats than in females. Despite 18-month-old female rats reaching a similar percentage of serum insulin increase than their male counterparts, male rats maintain a serum profile of lower insulin sensitivity than females in all the age groups, as has been previously reported in both rodents 24,31 and humans. 32
Although the underlying mechanism for age-related insulin resistance remains poorly defined, it has been proposed that increased oxidative stress and impaired antioxidant defenses are likely to be involved. 4,10,11 In our study, we found an increase of skeletal muscle oxidative damage with age, which takes place to a greater extent in male rats than in females and which is more enhanced in gastrocnemius muscle than in soleus, as levels of damage of both lipids and proteins suggested. In male rats, we observed a gradual increase of oxidative damage with age, whereas in females, the increase occurs between 3 and 9 months of age and tends to decrease at 18 months of age, suggesting that female rats may develop mechanisms to counteract the age-associated increase of oxidative damage.
Since a defensive role of UCP3 against skeletal muscle oxidative stress has been described, 33 sex differences in UCP3 level variations with age reinforce the idea that female rats have a higher protection against oxidative damage than males. Moreover, the sexual dimorphism found in the increase of skeletal muscle oxidative damage associated with age may also be attributed to differences in other components of the antioxidant defenses. In fact, a greater decrease of some mitochondrial antioxidant systems with age, such as glutathione and manganese superoxide dismutase (Mn-SOD) activity, has been recently reported in male mice compared to females. 34 Moreover, we have previously reported sex differences in skeletal muscle mitochondrial antioxidant enzyme capacity in rats of 6 22 and 18 months of age, 24 reinforcing the idea of a greater ability of female rats to maintain the efficiency of antioxidant enzymes compared to males.
The aforementioned higher age-associated increase of oxidative damage found in gastrocnemius muscle of both sexes compared to soleus points to a greater impact of age on glycolytic muscles compared to oxidative ones and agrees with previous studies. 35 However, neither gastrocnemius nor soleus oxidative damage was accompanied by the age-associated decline of muscle mass recently described in 30- and 36-month-old rats, 26,35 probably due to the high muscle plasticity to adapt to physiological changes and because muscle loss may appear at more advanced ages, when this plasticity would begin to fail.
Age-related insulin resistance may also be associated to the increase of adiposity that occurs with age, which is in turn linked to physical inactivity. 7,8 The increase of adiposity is accompanied by a decrease of serum adiponectin levels, which have been reported to be negative factors of insulin-stimulated glucose uptake. 12,36 In addition, the excess of adiposity is associated to the activation of inflammatory signaling pathways, such as IκB kinase complex (IKK-β) and c-jun N-terminal kinase (JNK), which inhibit the insulin signaling pathway in skeletal muscle and contribute to age-related insulin resistance. 37 –39 Interestingly, we found sex differences in the profile of age-increased adiposity, which could be involved in the sexual dimorphism found in age-associated impairment of insulin sensitivity.
The adiposity index increases with age following a similar profile to that of serum insulin levels, with male rats showing the greatest increase of adiposity at 9 months of age (72% increase in males and 50% increase in females with respect to their 3-month-old counterparts), whereas females exhibit it at 18 months (2% increase in males and 57% increase in females with respect to their 9-month-old counterparts). Because it has been shown in rats that age-associated impairment of insulin-stimulated glucose uptake occurs earlier in adipose tissue, at 8 months of age, than in skeletal muscle, at 24 months of age, 12 the greater increase of adiposity shown by male rats at 9 months of age would reinforce the idea of an earlier impairment of insulin sensitivity in male rats than in females.
Although tissues such as liver and white adipose tissue also contribute to the impairment of systemic insulin sensitivity, skeletal muscle is considered the most important. In our study, sex differences in the profile of insulin sensitivity with age do not exactly match the time course of the levels of key proteins of the skeletal muscle insulin signaling pathway, above all in the early ages. At 18 months of age, the higher gastrocnemius Akt protein levels and soleus Akt activation shown by female rats could be a response to attempt to maintain GLUT4 levels and may explain their higher capacity to counteract insulin resistance. This result is in concordance with other studies 26,40 in which Akt phosphorylation in 26- and 30-month-old female rats is associated with the maintenance of muscle mass with aging.
Thus, sexual dimorphism in insulin sensitivity could be related to changes in insulin signaling key protein levels, mainly in the most aged rats, suggesting that sex differences in these proteins would be manifested at more advanced ages than differences in the insulin-sensitivity serum profile probably, because both sexes may develop different strategies to attempt to counteract aging-associated insulin resistance. Nevertheless, an earlier impairment of white adipose tissue insulin sensitivity prior to that of skeletal muscle could also contribute to the alterations of the systemic insulin resistance markers. 9
Soleus and gastrocnemius muscles showed a different response to the effect of age on the insulin signaling pathway. Soleus muscle seemed to be better adapted than gastrocnemius to counteract age-associated changes in insulin sensitivity, as shown by levels of insulin signaling protein intermediates. Age brought about an impairment of gastrocnemius insulin signaling in both sexes, as indicated by the decrease of IRS-1, p-Akt/Akt ratio, and GLUT4 protein levels. However, IR-β levels did not decrease with age, supporting the idea that insulin signaling impairment with age could occur downstream of the insulin receptor, in agreement with a previous study. 13
The increase of IR-β levels observed only in the most aged rats could be understood as an attempt to counteract the impairment of insulin signaling by increasing their muscle insulin receptor levels. In soleus, although age decreased levels of IR-β, IRS-1, and total Akt proteins, insulin sensitivity could not have been compromised, as the maintenance of GLUT4 levels suggests, although the glucose uptake actually depends on GLUT4 translocation. Unfortunately, direct measurement of skeletal muscle glucose uptake has not been performed, which constitutes a limitation of our study. The enhanced Akt activation shown by the soleus muscle of the oldest group could be a response to counteract the decrease of the other insulin signaling intermediates. The different response that soleus and gastrocnemius muscles showed to the effect of age on insulin signaling was in agreement with a previous study reporting a greater age-associated insulin resistance impact in glycolytic than in oxidative muscle in rats of 24 months of age. 4
These data support the idea that insulin signaling seems to be more compromised in muscles composed primarily of glycolytic than oxidative fibers, 4,5 probably due to the preferential atrophy of fast-twitch fibers with age 4,6,9,41 and the greater decrease in expression of insulin signaling intermediates compared to slow-twitch fibers. 4 We found this different response to age of gastrocnemius and soleus muscles in up to 18-month-old rats, suggesting that differences found in the impact of age on the insulin signaling pathway may be evident prior to aging.
In summary, we have studied the changes in insulin sensitivity with age in male and female rats to discriminate whether the differences between sexes in the profile of age-associated variations of adiposity, skeletal muscle oxidative damage, and/or levels of insulin signaling pathway components may explain the sex differences in systemic insulin sensitivity previously found in aged rats. Overall, insulin sensitivity decreases with age in both sexes, and sex differences in both age-related increase of insulinemia and adiposity suggest an earlier impairment of insulin sensitivity in male rats than in females. However, sex differences in skeletal muscle oxidative damage and insulin signaling key protein levels were found only in the most aged rats. Thus, 18-month-old female rats showed a greater ability to respond to the impairment of the insulin signaling pathway than their male counterparts. In addition, gastrocnemius muscle showed more age-associated oxidative damage and insulin resistance impact than soleus. These results suggest that sex differences found in the age-associated impairment of insulin sensitivity would not be attributable to a different profile of age-associated alterations in the skeletal muscle insulin signaling pathway, probably because sex differences in the decline of insulin signaling proteins are manifested at more advanced ages.
Footnotes
Acknowledgments
This work was supported by Fondo de Investigaciones Sanitarias of the Spanish Government (PI042294, PI042377 and PI060293). Y. Gómez-Pérez was funded by grants from the Comunitat Autònoma de les Illes Balears.
Author Disclosure Statement
No competing financial interests exist.