
Abstract
An association study of heterogeneous nuclear ribonucleoprotein (hnRNP)-A1 was carried out in a population of 274 patients with frontotemporal lobar degeneration (FTLD) and 287 with Alzheimer disease (AD) as compared with 344 age- and gender-matched controls. In addition, we evaluated expression levels of hnRNP-A1 and its regulatory microRNA (miR)-590-3p in blood cells from patients and controls. A statistically significant increased frequency of the hnRNP-A1 rs7967622 C/C genotype was observed in FTLD, but not in AD, in patients as compared to controls (23.0 versus 15.4%; p = 0.022, odds ratio [OR] 1.64, confidence interval [CI] 1.09–2.46). Stratifying according to gender, a statistically significant increased frequency of the hnRNP-A1 rs7967622 C/C genotype was observed in male patients as compared to male controls (23.1 versus 11.3%; p = 0.015, OR 2.36, CI 1.22–4.58 but not in females. Considering the rs4016671 single-nucleotide polymorphism (SNP), all patients and controls were wild type. Significantly increased hnRNP-A1 relative expression levels in peripheral blood mononuclear cells (PBMCs) was observed in patients with AD, but not with FTLD, as compared to controls (2.724 ± 0.570 versus 1.076 ± 0.187, p = 0.021). Decreased relative expression levels of hsa-miR-590-3p was observed in patients with AD versus controls (0.685 ± 0.080 versus 0.931 ± 0.111, p = 0.079), and correlated negatively with hnRNP-A1 mRNA levels (r = −0.615, p = 0.0237). According to these findings, hnRNP-A1 and its transcription regulatory factor miR-590-3p are disregulated in patients with AD, and the hnRNP-A1 rs7967622 C/C genotype is likely a risk factor for FTLD in male populations.
Introduction
There is increasing evidence that miRNAs represent key regulators of several biological phenomena, including cell proliferation and differentiation, apoptosis, signal transduction, and organ development. 4 The hnRNPA1-encoding gene (HNRPA1) maps at 12q13.13. It contains a glycine-rich consensus domain that is characteristic of other heterogeneous nuclear ribonucleoproteins, including TAR DNA-binding protein 43 (TDP43), which is known to play a role in frontotemporal lobar degeneration (FTLD) and other neurodegenerative disorders of the central nervous system. 5 TDP43 acts as transcription repressor and activator of exon skipping. 6,7 It has been shown to be capable of binding directly to several proteins of the hnRNP family, including hnRNPA1, that seem to be necessary for the splicing inhibitory activity of TDP43. 8 Nevertheless, in TDP43-positive brains there are no changes in subcellular distribution, no labeling of pathologic inclusions, and no biochemical alterations of hnRNPA1 (and other members of the hnRNP family). 9
hnRNPA1 plays a role also in spinal muscular atrophy, an autosomal recessive neuromuscular disorder due to mutations in survival of motor neuron-1 (SMN1). In humans, a nearly identical copy gene is present (SMN2), but it cannot compensate for the loss of SMN1 because of a silent nucleotide difference in SMN2 exon 7. hnRNPA1 has been shown to function with specificity in repression of SMN2 exon 7 splicing. 10
Given the potential importance of hnRNPA1, we carried out a case–control association study in a population consisting of 274 patients with FTLD and 287 patients with AD as compared to 314 age-matched healthy subjects, by analyzing two single-nucleotide polymorphisms (SNPs) (rs7967622 and rs4016671) (Fig. 1). The former is a tagging SNP covering 100% gene variability, and the latter is located in the 3′-UTR region, in a predicted binding site of miR-590-3p (Fig. 2). Thus, we analyzed miR-590-3p expression levels in peripheral blood mononuclear cells (PBMCs) and correlated them with hnRNP-A1 mRNA levels.
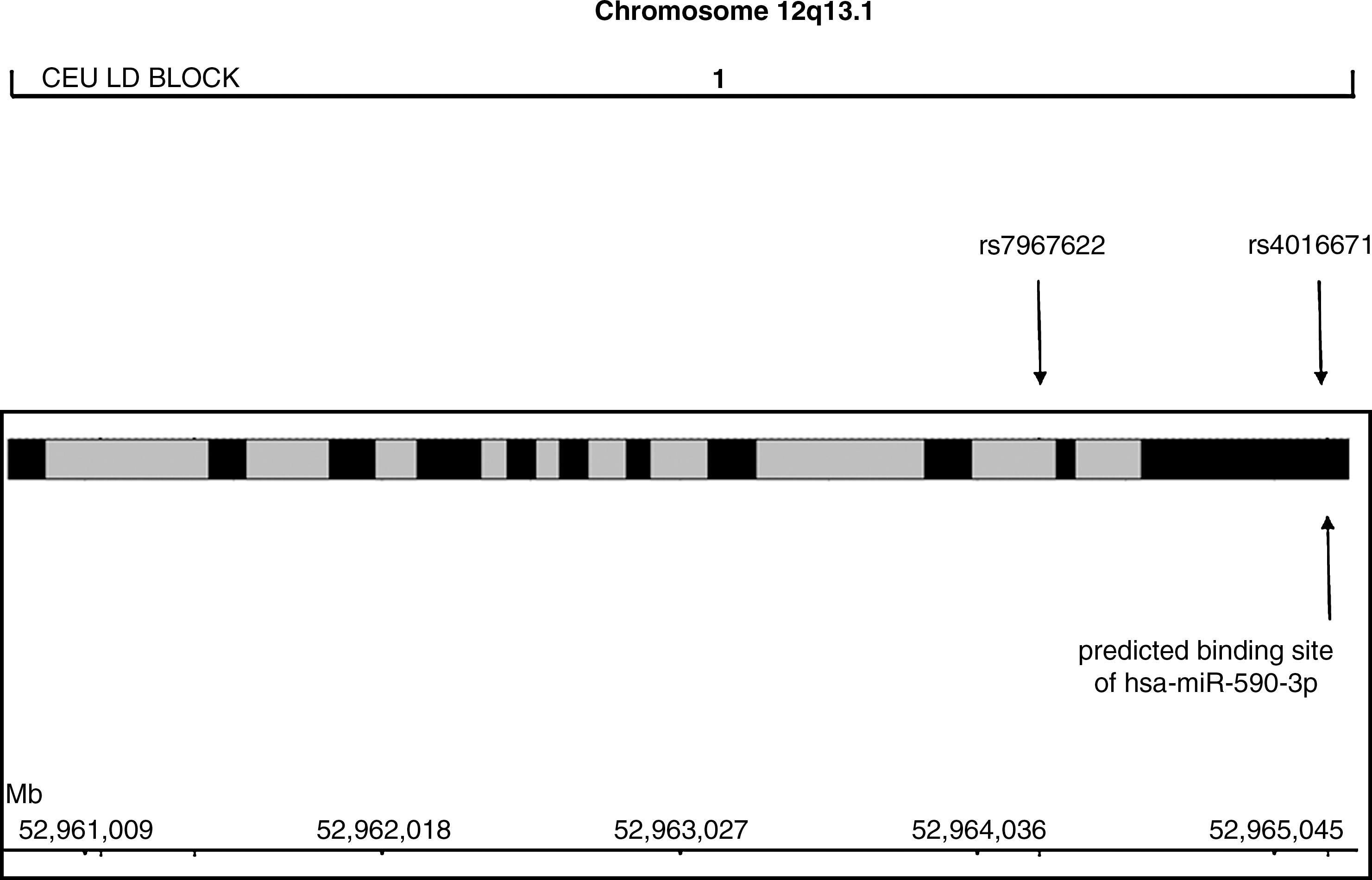
Localization of single-nucleotide polymorphisms (SNPs) selected according to Hapmap (
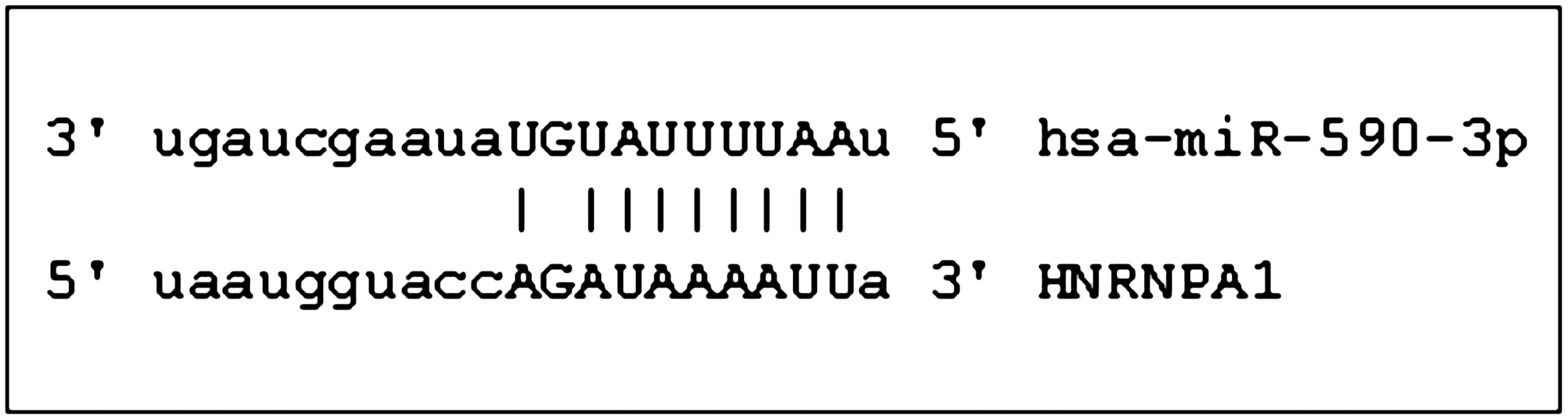
hsa-miR-590-3p and hnRNP-A1 sequence alignment.
Materials and Methods
Patients and controls
A total of 274 Italian patients with FTLD (130 males and 144 females, mean age at onset ± standard error of the mean [SEM] 66.5 ± 0.59 years) and 287 with AD (105 males and 182 females, mean age at onset ± SEM 70.7 ± 0.61 years) were consecutively recruited at the Alzheimer Unit of Fondazione Cà Granda, IRCCS Ospedale Maggiore Policlinico (Milan), Ospedale L. Sacco (Milan), Ospedale S. Raffaele Turro (Milan), IRCCS Centro S. Giovanni di Dio-Fatebenefratelli (Brescia), University of Turin, and Casa di Cura Santa Maria di Castellanza (Varese). All patients underwent a standard battery of examinations, including medical history, physical and neurological examination, screening laboratory tests, neurocognitive evaluation, and imaging. Cognitive dysfunctions were assessed by the Clinical Dementia Rating (CDR), the Mini-Mental State Examination (MMSE), the Frontal Assessment Battery (FAB), the Wisconsin Card Sorting Test (WCST), and the Tower of London test.
FTLD was diagnosed according to current consensus criteria, 11 as well as their subsequent revision, 12 identifying three clinical syndromes: Behavioral variant frontotemporal dementia (bvFTD, n = 237, 113 males and 124 females, mean age at onset ± SEM 66.2 ± 0.65 years), progressive aphasia (PA, n = 22, 8 males and 14 females, mean age at onset ± SEM 67.2 ± 2.15 years), and semantic dementia (SD, n = 15, 9 males and 6 females, mean age at onset ± SEM 65.7 ± 2.59 years). Ten patients reported a documented family history for dementia, whereas 57 patients had a reported, but not documented, family history. For the AD group, the presence of significant vascular brain damage was excluded (Hachinski Ischemic Score <4). The diagnosis of probable AD was made according to National Institute of Neurological and Communicative Disorders and Stroke and the Alzheimer's Disease and Related Disorders Association (NINCDS-ADRDA) criteria. 13 The control group consisted of 344 nondemented volunteers matched for ethnic background and age (142 males and 202 females, mean age ± SEM 68.5 ± 0.55 years) without memory and psychobehavioral dysfunctions (MMSE ≥28). Characteristics of patients and controls are summarized in Table 1.
Age at disease onset.
p < 0.001, OR = 3.44, CI 95% = 2.40–.4.93.
CON, Control; AD, Alzheimer disease; FTLD, frontotemporal lobar degeneration; bvFTD, Behavioral variant frontotemporal dementia; PA, behavioral variant frontotemporal dementia; PA, progressive aphasia; SD, semantic dementia; M, male; F, female; SEM, standard error of the mean; OR, odds ratio; CI, confidence interval.
This study was approved by the Institutional Review board of the Department of Neurological Sciences, University of Milan, Fondazione Cà Granda, IRCCS Ospedale Maggiore Policlinico, Mangiagalli e Regina Elena. Informed consent to participate in this study was given by all subjects or their caregivers.
DNA isolation
High-molecular-weight DNA was isolated from whole blood using a Flexigene Kit (Qiagen, Hildren, Gemany), as described by the manufacturer. The amount of DNA for each sample was determined by measuring the optical density at 260-nm wavelength using a spectrophotometer (Eppendorf AG, Germany). DNA samples were aliquoted and stored at −20°C until use.
Genotyping
One tagging SNP (rs7967622) and a SNP located in the 3′-UTR region (rs4016671) were analyzed by using TaqMan methodology. Each Taqman 5′-nuclease assay employed 25 ng of genomic DNA as template. Assay-on-demand products (ABI assay IDs: C___27275446_10 and C___44849276_10) were used for rs7967622 and rs4016671 genotyping, respectively.
Probes specific for these variants were labeled with 6-carboxyfluorescein (6-FAM) and VIC® as reporter dyes and MGB-NFQ (ABI) as quencher. Assays were performed in 25-μL reactions in 96-well plates using an ABI PRISM®7000 instrument (ABI). PCR mix consisted of 12.5 μL of universal master mix, 0.6 μL of probe, and water to reach the final volume. Thermocycling consisted of 10 min at 95°C for AmpliTaqGold activation, and 50 cycles at 92°C for 15 sec and 60°C for 1 min. To further confirm results obtained with allelic discrimination, 20 samples randomly selected were analyzed by direct sequencing (primers used available upon request). Apolipoprotein E (ApoE) genotyping was carried out by polymerase chain reaction–restriction fragment length polymorphism technique (PCR-RFLP), as previously described. 14
Progranulin and MAPT mutation scanning
As GRN mutations were demonstrated in sporadic cases, 15,16 the entire open reading frame, including the noncoding exon 0 and exon–intron boundaries of exons 1–12 of the PGRN gene, was sequenced in FTLD patients using specific primers, as previously described. 15 –17 Briefly, each PCR was carried out in a 50-μL total volume containing 25 ng of genomic DNA, 12.5 pmol of each specific primer, 0.6 μM of each dATP, dTTP, dCTP, and dGTP, and 1 U of Taq DNA polymerase (Applied Biosystems, ABI). Amplification conditions consisted of an initial denaturation step at 94°C for 5 min followed by 38 cycles of 94°C for 30 sec, Tm for 30 sec and 72°C for 45 sec, and a final extension step of 72°C for 10 min.
DNA fragments were purified using the ExoSAP-IT® Kit (USB, USA), according to instructions of the manufacturer, and then direct sequence was performed with an ABI PRISM 3130 gene analyzer (ABI). Sequencing PCR was carried out in a 20-μL total volume containing 1 μL of BigDye Terminator v3.1 (ABI) and 1.6 pmol of each primer and water to reach the final volume. Thermocycling consisted of 25 cycles of 96°C for 30 sec, 50°C for 20 sec, and 60°C for 3 min.
Because there is agreement that MAPT mutations are rare, lead to an early onset of the disease (often in the fifties), and no sporadic cases have been reported so far, 18 MAPT exons 1 and 9–13 were sequenced as previously described 19 in patients with an onset before 60 years of age or with a documented familial history (n = 65) to exclude the presence of causal mutations.
Total mRNA extraction and hnRNP-A1 quantitative transcriptional analysis by real-time PCR
Total RNA from PBMCs was extracted with the single-step acid phenol method, using TRIzol (Invitrogen). RNA was then reverse-transcribed using the Ready-To-Go You-Prime First-Strand Beads kit (Amersham Biosciences). For a quantitative estimate of mRNA levels, an ABI 7000 Sequence Detector with dual-labeled TaqMan probes was used. A specific TaqMan probe was employed (Hs01656228_s1, Applied Biosystems), and the relative amount of hnRNP-A1 mRNA determined by comparison with the housekeeping 18SrRNA probes (Hs99999901_s1, Applied Biosystems). The final volume reaction was 20 mL, using TaqMan Universal Master Mix (ABI 4324018, Applied Biosystems). Cycle parameters used consist of 2 min at 50°C and 10 min at 95°C, followed by 40 cycles of 15 sec at 95°C for denaturation and 1 min at 60°C for annealing/extension. Relative mRNA levels were calculated as follows: 2 − [(DCt(sample) − DCt(calibrator)] = 2-DDCt, where DCt equals the cycle threshold (Ct) (molecule under analysis) − Ct (housekeeping gene). 20
hsa-miR-590-3p extraction and quantitative analysis by real-time PCR
Total RNA enriched for small RNA species was isolated using the mirVana miRNA Isolation Kit according to the instruction of the manufacturer (Applied Biosystems), and the RNA concentration was determined by using a NanoDrop ND-3300® Fluorospectrometer. The reverse transcription reaction was performed using the TaqMan@MicroRNA Reverse Transcription kit (Applied Biosystems). Briefly, the reaction consists of 10 ng of RNA, 0.15 μL of 100 mM dNTPs, 1 μL of MultiScribe Reverse Transcriptase (50 U/μL), 1.5 μL of 10X Reverse Transcription Buffer, 3 μL of 1X stem–loop reverse transcriptase (RT) primer, 0.19 μL of RNase inhibitor 20 U/μL, and 4.16 μL of nuclease-free water run in a total reaction volume of 15 μL and incubated at 16°C for 30 min, 42°C for 30 min, and 85°C for 5 min in a thermal cycler. The quantitative real-time PCR was performed on the ABI 7500 FAST system (Applied Biosystems). miR-590-3p was detected with a single TaqMan® miRNA assay (Applied Biosystems). Each reaction was performed in a total volume of 20 μL consisting of 1 μL of TaqMan microRNA assay 20X, 1.33 μL of products from the RT reaction (minimum 1:15 dilution), 10 μL of TaqMan 2X Universal PCR Master Mix, no amperase UNG, and 7.67 μL of water. The amplification protocol was performed according to the instruction of the manufacturer. Ct values were calculated using the SDS software v.2.3 using automatic baseline settings and a threshold of 0.2. The Ct value of the endogenous control gene encoding for the small nuclear RNA RNU48 was subtracted from the corresponding Ct value for the target gene, resulting in the ΔCt value, which was used for the subsequent relative quantification with the ΔΔct method. 20,21
Statistical analysis
Statistical analysis was performed using the Sigma Stat 3.1 software. Allelic and genotypic frequencies were obtained by direct counting. The chi-squared test was used to test for Hardy–Weinberg equilibrium (HWE,
Multivariate logistic regression analysis, adjusted for age and gender, was carried out with the SPSS software version 13.0 (SPSS for Windows, Rel. 13.0. Chicago: SPSS Inc.), and adjusted odds ratios (OR) with related 95% confidence interval (CI) calculated. Significance threshold was set at p < 0.05 The available bioinformatic database (
Results
Only patients with sporadic FTLD who did not carry any causal mutation in GRN or MAPT were included in this study (n = 274 and n = 287, respectively). As expected, the ApoE ε4 allele frequency was significantly higher in AD patients than in controls (43.6 versus 18.3%, p < 0.001, OR = 3.44, CI 95% = 2.40–4.93), whereas no differences were observed between patients with FTLD and controls (p > 0.05; Table 1). A statistically significant increased frequency of the hnRNP-A1 rs7967622 C/C genotype was observed in FTLD patients as compared with controls (23.0 versus 15.4%; p = 0.022, OR 1.64, CI 1.09–2.46, power = 0.630; Table 2). Stratifying according to gender, a statistically significant increased frequency of the hnRNP-A1 rs7967622 C/C genotype was observed in male patients as compared with male controls (23.1 versus 11.3%; p = 0.015, OR 2.36, CI 1.22–4.58, power = 0.686; Table 3), but not in female patients compared with female controls (22.9 versus 18.3%, p > 0.05; Table 3). Stratifying according to the ApoE status, no differences in rs7967622 distribution were observed (p > 0.05, data not shown). Applying the multivariate logistic regression analysis, adjusted for age, gender, and ApoE status, we obtained a borderline p value ( = 0.05) in the male population.
p = 0.022, OR 1.64, CI 1.09–2.46.
SNP, Single-nucleotide polymorphism; AD, Alzheimer disease; FTLD, frontotemporal lobar degeneration; bvFTD, Behavioral variant frontotemporal dementia; PA, behavioral variant frontotemporal dementia; PA, progressive aphasia; SD, semantic dementia.
p = 0.015, OR 2.36, CI 1.22–4.58.
SNP, Single-nucleotide polymorphism; FTLD, frontotemporal lobar degeneration.
No differences in hnRNP-A1 rs7967622 allele and gene frequencies were observed in AD patients as compared with controls (p > 0.05), even adjusting for gender, age, and ApoE status. Considering the rs4016671 SNP, all patients and controls were wild-type (A/A). A statistically significant increased of hnRNP-A1 relative expression levels in PBMC was observed in AD patients as compared with controls (2.724 ± 0.570 versus 1.076 ± 0.187, p = 0.021; Fig. 3), but not in FTLD patients (1.495 ± 0.426 versus controls, p ≥ 0.05; Figure 3).
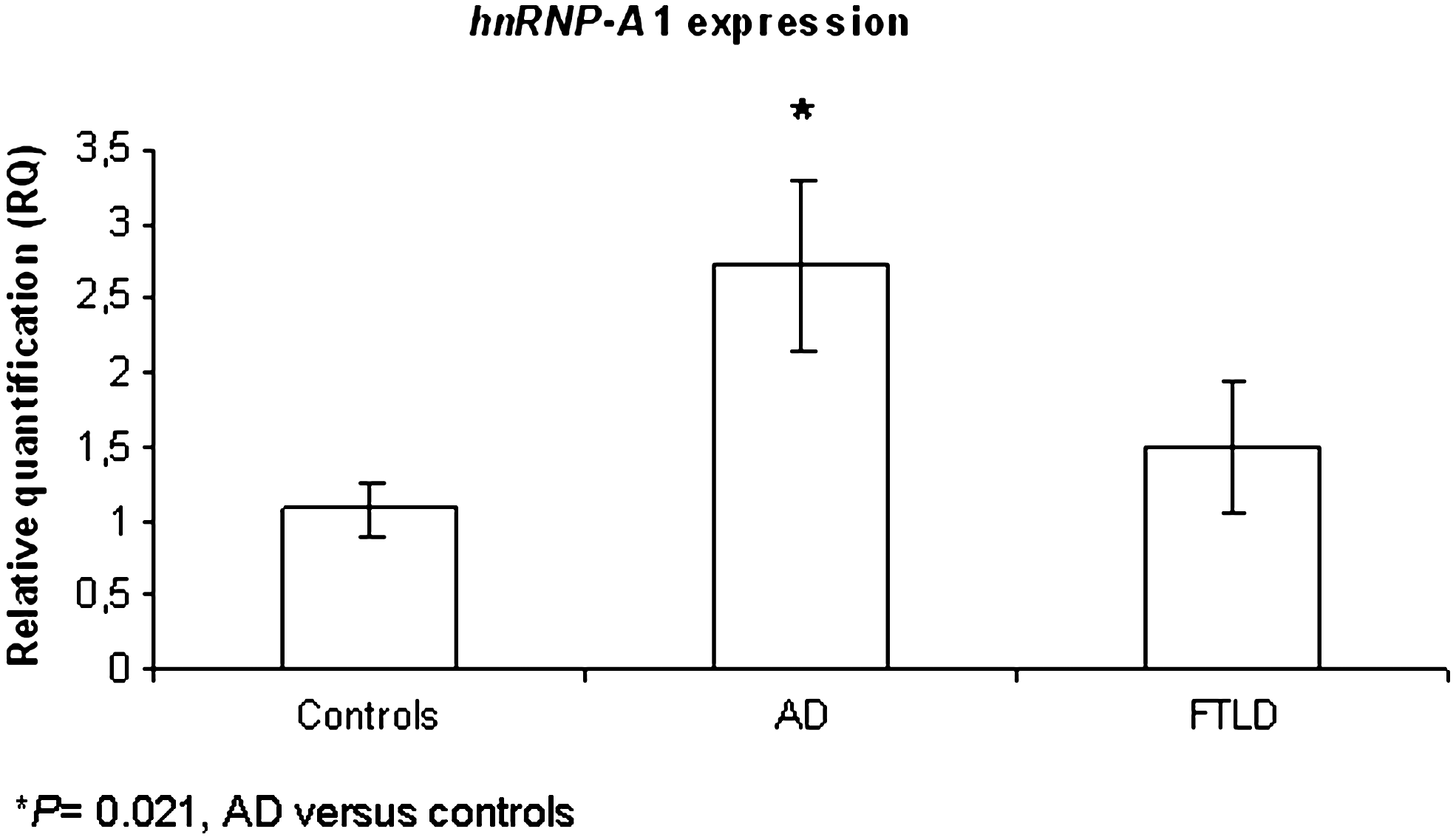
hnRNP-A1 expression levels in peripheral blood mononuclear cells (PBMCs) from controls and Alzheimer disease (AD) and frontotemporal lobar degeneration (FTLD) patients, expressed as fold increase ± standard error of the mean (SEM).
Decreased relative expression levels of hsa-miR-590-3p was observed in patients with AD as compared to controls, although the significant threshold was not reached (0.685 ± 0.080 versus 0.931 ± 0.111, p = 0.079; Fig. 4). A negative correlation between hnRNP-A1 mRNA and hsa-miR-590-3p expression levels was found in all subjects (r = −0.316, p = 0.05, data not shown). Considering each group separately, a statistically significant correlation was observed in patients with AD (r = −0.615, p = 0.0237; data not shown), and a similar trend was observed in both patients with FTLD and controls (r = −0.373 and −0.261, respectively, p > 0.05; data not shown). An effect of rs4016671 on miR-590-3p binding would be conceivable, but we could not test this hypothesis because none of individuals studied carried the polymorphic allele.

hsa-miR-590-3p expression levels in peripheral blood mononuclear cells (PBMCs) from controls and Alzheimer disease (AD) and frontotemporal lobar degeneration (FTLD) patients, expressed as fold increase ± standard error of the mean (SEM).
Discussion
On the basis of the data obtained in our population, the hnRNP-A1 rs7967622 C/C genotype is likely a risk factor for FTLD in patients without causal GRN and MAPT mutations, whereas it has no effect on the susceptibility to AD. However, expression levels of hnRNP-A1 were increased, and levels of miR-590-3p transcript decreased, in patients with AD. These results suggest a disregulation of hnRNP-A1, possibly mediated by miR-590-3p, as demonstrated by a marked negative correlation between the two transcripts. On the basis of these data, we can hypothesize an increased production of the hnRNP-A1 transcription factor, which could play a role in the pathogenesis of AD, because it is involved in the maturation of the APP mRNA. 1 Such deregulation has not been observed in FTLD, although the population studied is quite small and larger studies are needed to draw definitive conclusions.
Genetic variability in hnRNP-A1 influences the risk for developing FTLD, although to a quite small extent (OR = 1.64). The rs7967622 tagging SNP is located in an intronic region, and the mechanism by which it increases the risk to develop FTLD is not known. A possible effect on alternative splicing has been excluded because the SNP is not part of any splicing consensus sequence (
When applying the correction for age, we obtained a borderline p value, possibly due to the fact that cases were younger than controls. On the other hand, this would imply a decreased risk of “false” controls, as the development of FTLD in elderly people is unlike. Considering each clinical syndrome separately, we found similar data in all three of them, although the small size of the population of PA and SD patients did not allow us to get statistically significant conclusions. Stratifying for gender, the effect was even stronger in males, whereas it got lost in females, suggesting a different genetic background between sexes, possibly interacting with multiple environmental factors, as well as ethnicity. In this regard, individuals included in this study were recruited in six sites located in northern Italy. They were matched well ethnically because they were all Caucasians and living in this region for at least two generations. We cannot, however, exclude relatedness with certainty because no haplotype sharing data are available.
One of the aims of this work was to test whether the presence of the polymorphic allele of the rs4016671 SNP, which is located in the hnRNP-A1 3′-UTR, may influence the strength of miR-590-3p to its binding site. Unexpectedly, despite a predicted allelic frequency of about 10% in Caucasians (
In conclusion, according to these findings, hnRNP-A1 and its transcription regulatory factor miR-590-3p are deregulated in PBMCs from patients with AD, possibly playing a role in the pathogenesis of the disease through the modulation of APP. The rs7967622 C/C genotype is likely a risk factor for FTLD in male populations, whereas it has no effect on the susceptibility to AD. These preliminary data need to be replicated in an independent population, and possibly in brain tissues from patients. In addition, the experimental proof that miR-590-3p is indeed regulating hnRNP-A1 mRNA and protein levels would further support these preliminary data.
Footnotes
Acknowledgments
This work was supported by grants from Monzino and Cariplo (2009-2633) Foundations, IRCCS Ospedale Maggiore Milano, Programma Strategico RF 2007, conv. PS39, Italian Ministry of Health, Progetto Regione Lombardia (Delibera N°VIII/008724); FIRB 2006 “Gen-Etica”, Associazione per la Ricerca sulle Demenze (ARD), and Ing. Cesare Cusan.