
Abstract
Adult skeletal muscle possesses a resident stem cell population called satellite cells, which are responsible for tissue repair following damage. Satellite cell migration is crucial in promoting rapid tissue regeneration, but it is a poorly understood process. Furthermore, the mechanisms facilitating satellite cell movement have yet to be elucidated. This study investigates the process of satellite cell migration, revealing that they undergo two distinct phases of movement, first under the basal lamina and then rapidly increasing their velocity when on the myofiber surface. Most significantly, we show that satellite cells move using a highly dynamic blebbing or amoeboid-based mechanism and not via lamellipodia-mediated propulsion. We show that nitric oxide and noncanonical Wnt signaling pathways are necessary for regulating the formation of blebs and the migration of satellite cells. In summary, we propose that the formation of blebs and their necessity for satellite cell migration has significant implications in the future development of therapeutic regimes aimed at promoting skeletal muscle regeneration.
Introduction
A few studies have suggested that activated satellite cells can migrate along the entire length of myofibers in vivo and are also capable of moving between fibers. 7,8 However, experimental proof for this notion in vivo is still lacking. Indeed, there is much debate regarding the exact location of satellite cell migration with evidence for cell movement taking place both above and below the basal lamina. 9 Migration has been traditionally explained by the force generated by structures called lamellipodia that form at the leading edge of the cells and the retraction of the cell at the rear. The propagation of the lamellipodia has been shown to be mediated by the continuous growth of filamentous actin through the process of monomer polymerization at the front of the cell. 10 Most studies investigating muscle cell migration have been limited to using immortalized myoblast cell lines and support the notion that these cells move using lamellipodia. 11 –15 A recent elegant study by Siegel et al. 16 investigated satellite cell migration on isolated myofibers as opposed to cell lines and revealed a number of novel behavioral characteristics of satellite cells, including the ability of cells to move as cohorts. 16 The use of isolated myofibers permits the observation of primary satellite cells moving on their native substrate in a near-physiological context and provides an opportunity to investigate experimentally the mechanisms that regulate their migration through the use of pharmacological inhibitors, blocking antibodies or the addition of growth factors or chemokines. 13
At present, the signaling molecules that regulate satellite cell migration remain undefined. However clues to their identity come from the key players implicated in cell migration in other biological contexts. The first is nitric oxide (NO), a potent short-lived signaling molecule that regulates cell migration in numerous biological systems, including endothelial, epithelial, and trophoblast cells. 17 –19 NO is produced by a group of enzymes known as the nitric oxide synthases (NOS), which exist in three isoforms—neuronal NOS (nNOS), endothelial NOS (eNOS), and inducible NOS (iNOS). Recent work shows that NO plays an important role in the regulation of satellite cell function including their activation following injury. 20
The second is the Wnt signaling pathway. This family of highly conserved secreted proteins regulates almost every stage of animal development and tissue homeostasis. Previously, we have shown that members of the Wnt family have a ligand-specific effect on satellite cell proliferation based on the canonical signaling pathway involving β-catenin. 21 However, specific Wnt ligand and receptor combinations activate an alternative signaling cascade called the planar cell polarity (PCP) pathway, which is extensively deployed during cell movement both during normal development and disease progression. 22,23
The study of Siegel et al. 16 has demonstrated the power of coupling time-lapse microscopy with the single-fiber isolation technique. Previous work has suggested that satellite cells are a heterogeneous population, with approximately 10% of cells never having expressed the myogenic regulatory factor Myf5, and that the Myf5− subpopulation corresponds to the self-renewing skeletal muscle stem cells. 24 Pertinent to this study, Kuang et al. 24 have suggested that the initial division of satellite cells takes place within the original satellite cell niche beneath the basal lamina of the myofiber, and that it is the plane of division (perpendicular to, or parallel to the basal lamina) within this niche that determines the fate of daughter cells; perpendicular divisions result in asymmetric divisions within the satellite cell population, and that these divisions occur predominantly in the Myf5− population. 24
The study of Siegel et al. 16 has demonstrated the power of coupling time-lapse microscopy with the single-fiber isolation technique. In this study, we have aimed to establish the mechanism that permits satellite cell migration. Here we report a number of novel properties of motile satellite cells, including evidence for lamellipodia-based migration immediately after single-fiber isolation. However, our most striking observation is that satellite cells employ blebbing or amoeboid-based movement to move along the myofibers after emerging from the basal lamina. Blebbing-based migration is a mechanism permitting rapid cell movement mediated by the extension of plasma membrane protrusions into the extracellular matrix driven by local hydrodynamic changes in cytosolic pressure, 25 (also reviewed by Charras and Paluch.) 26 Blebbing occurs shortly after the cells emerge onto the surface of the myofibers, and our time-lapse microscopy studies capture satellite cells extending protrusions approximately 7% of cell length over periods of 15 sec before initiating their retraction. Furthermore, we have determined that satellite cells migrate at very high velocities. We also show that entire clusters of satellite cells can move along the myofiber and that this movement also requires membrane blebbing. The formation of blebs and the migration of satellite cells were found to be dependent on both nitric oxide and noncanonical Wnt signaling, suggesting possible new regulatory mechanisms for bleb formation and novel therapeutic targets for muscle regeneration. Finally, we show that intracellular inhibition of a key regulator of bleb formation causes satellite cells to form lamellipodia.
Materials and Methods
Single myofiber culture
Myofibers were isolated from the extensor digitorum longus (EDL) muscle of 4-month-old female Swiss mice as described previously.
21
Briefly, an undamaged EDL muscle was dissected with both tendons intact, and the single fibers were liberated through digestion with 0.1% type I collagenase in Dulbecco modified Eagle medium (DMEM) at 37°C 5% CO2. Using tapered glass pipettes, single fibers were plated out in floating culture wells containing DMEM supplemented with 10% horse serum and 0.5% chick embryo extract. When required, small molecule inhibitors 5 mM N-nitro-
Time-lapse microscopy
Myofiber cultures and C2C12 myoblast cells were monitored using two separate phase-contrast microscope systems that had controlled-environment chambers maintained at 37°C, 5% CO2. Time-lapse video was taken at rate of 1 frame every 15 min over 72 hr for satellite cells using a 10× objective, a previously well-studied time course of satellite cell progression following skeletal muscle injury 21,27 or for 24 hr for C2C12 cells. In separate analysis, high-power time-lapse microscopy using a 40× objective was carried out, enabling the visualization of cell-surface blebbing on single satellite cells. For such analysis, cells were cultured for 48 hr then selected and subsequently tracked at a rate of 1 frame per 20 sec.
Immunocytochemistry
Immunocytochemistry of satellite cells on single myofibers was carried out as described previously. 21 Briefly, myofibers were fixed gently in 2% paraformaldehyde (PFA) in phosphate-buffered saline (PBS) for 10 min and washed three times in PBS. Myofibers were permeabilized in a solution of 20 mM HEPES, 300 mM sucrose, 50 mM NaCl, 3 mM MgCl2, and 0.5% Triton X-100 (pH 7) at 4°C for 15 min and incubated in blocking wash buffer (5% newborn calf serum in PBS containing 0.01% Triton X-100) for 30 min prior to antibody incubation. Antibodies were diluted and preblocked in wash buffer for 30 min prior to addition to the myofibers. Primary antibodies used were: Polyclonal rabbit anti-Laminin (Sigma L-9393 1:200), monoclonal mouse anti-Pax7 (Developmental Studies Hybridoma Bank [DSHB] 1:1), polyclonal rabbit-anti-myogenin (Santa Cruz biotechnology 1:200), polyclonal rabbit anti-iNOS (Santa Cruz Biotechnology 1:200), and polyclonal rabbit anti-eNOS (Santa Cruz Biotechnology 1:200). All primary antibodies were incubated overnight at 4°C. Primary antibodies were visualized using the following secondary antibodies: Alexa Fluor goat anti-mouse 488 (Molecular probes A11029) and Alexa Fluor goat anti-rabbit 633 (Molecular probes A21071). Secondary antibodies were used at 1:200 and incubated at room temperature for 45 min. All myofibers were mounted in fluorescent mounting medium (DAKO) containing 7.5 mg/mL 4′,6-diamidino-2-phenylindole (DAPI) for nuclear visualization.
Fluorescence and confocal microscopy
Mounted myofibers were analyzed using a Zeiss Axioscope fluorescent microscope, and images were captured using an Axiocam digital camera system and Axiovision image analysis software (version 4.7). For confocal analysis, myofibers were analyzed using a Leica DMIRE2 confocal microscope. Z-stack images were taken every 0.5 μM, and three-dimensional reconstructions were performed using Leica confocal analysis software.
Scanning electron microscopy
Single myofibers were fixed following 24, 48, and 72 hr in standard culture conditions in 2% PFA for 10 min. Myofibers were dehydrated through 30%, 50%, 70%, 80%, 90%, and 100% ethanol solutions (15 min for each step) and transferred to a critical point drier (Balzers CPD 030, using liquid carbon dioxide). Dried myofibers were then carefully transferred to scanning electron microscopy (SEM) chucks using microforceps under a light microscope. Myofibers were then gold-coated using an Edwards S150B sputter-coater. Coated myofibers were then analyzed under a FEI 600F SEM using the accompanying analysis software for image collection.
C2C12 cell culture
C2C12 cells were cultured in DMEM containing GlutaMAX and pyruvate supplemented with 1% penicillin/streptomycin and 10% fetal calf serum. For migration analysis, cells were plated out in four-well plates, supplemented with the appropriate inhibitors (5 mM L-NAME and/or 25 μM SP600125) and grown until confluent. Plates were then taken for time-lapse microscopy.
Image and movie analysis
All image analysis was carried out using freeware package ImageJ (version 1.4.3). Single C2C12 and satellite cells were individually and manually tracked using the plugin MTrackJ. Bleb dynamics were quantified using ImageJ. Bleb number was quantified manually on SEM images of cells using Adobe Photoshop version CS2; only SEM images of whole cells were used for this quantification. Satellite cell differentiation profiles following differing culture conditions were manually assessed using quantification of live images through the Zeiss axioscope and axiovision digital camera system.
Results
Migration of satellite cells
Investigations carried out by ourselves and others have shown that satellite cells undergo a program of molecular activation a few hours after single-fiber isolation and emerge from their sublaminar position at about 24 hr. 16,21,27 We found that after this time point, cells are able to move rapidly at approximately 50 μm/hr (n = 33 cells tracked). This rate of migration is faster than most other cells examined to date (Fig. 1A). However, detailed examination of migration speeds following single-fiber isolation showed distinct phases of migration speed. We found that soon after fiber isolation (0–18 hours) the satellite cells moved, but at a considerably lower speed (27 μm/hr, n = 33 cells tracked) than at approximately 24 hr, where they rapidly accelerated. After approximately 36 hr, migration rates of satellite cells decreased and by 54 hr movement decreased further (Fig. 1B). After approximately 72 hr, cells had formed clusters of progeny on the surface of myofibers (Supplementary Fig. 1F, arrowheads). Examination of time-lapse movies revealed interesting migratory patterns, some previously noted by Siegel et al. 16 Although movement was decreased following 54 hr of culture, there were frequent instances when whole clusters of cells could be observed migrating as a cohort along the surface of myofibers, and the clusters subsequently splitting into separate groups of cells (Supplementary Fig. 1I). We also found instances of satellite cells moving from one fiber to another (Supplementary Fig. 1L–N). At other times, individual satellite cells would migrate toward each other and then appear to make contact followed by them subsequently moving as a pair (Supplementary Fig.1O–Q).
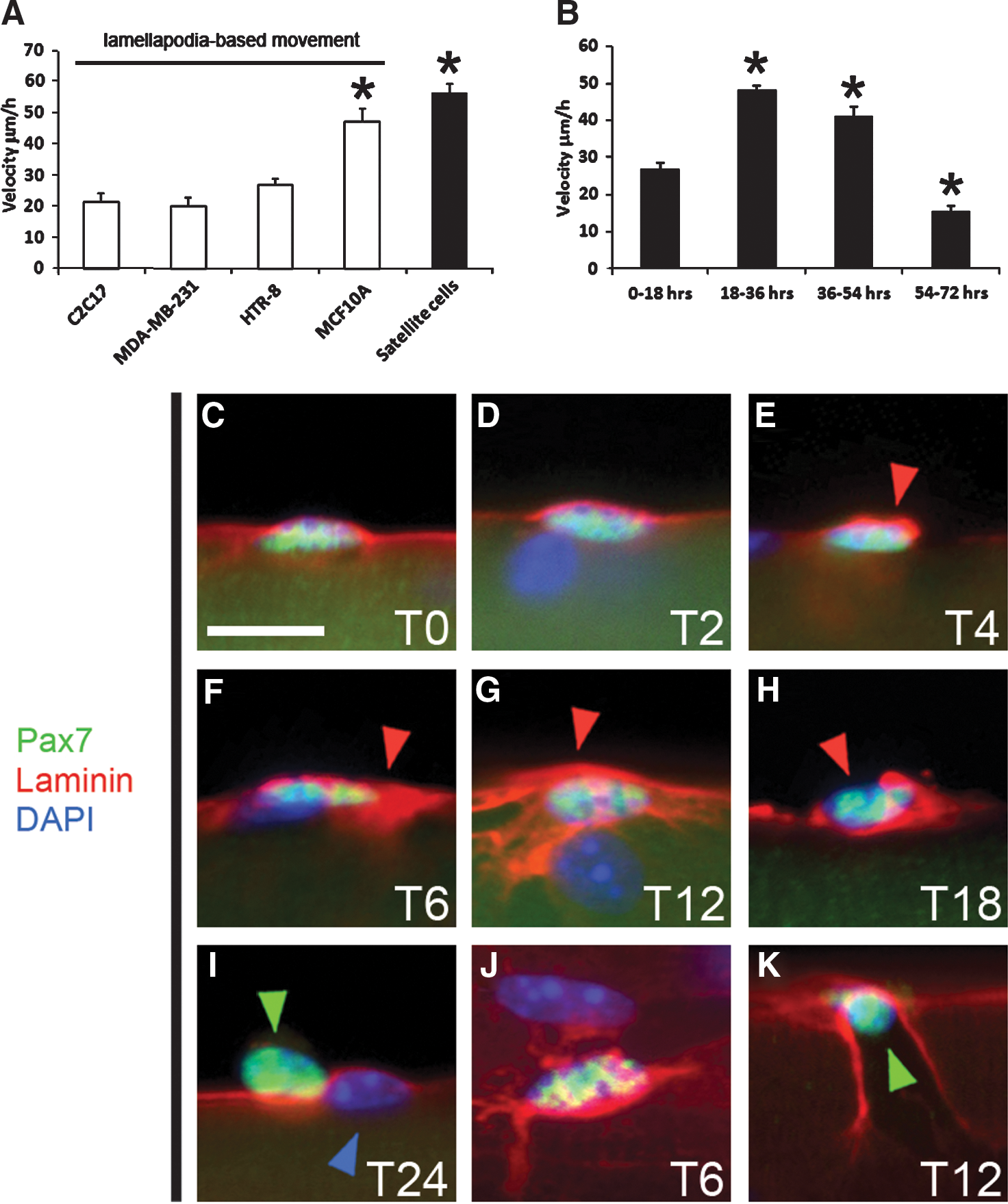
Rapid satellite cell migratory phase occurs following emergence from beneath the basal lamina. (
Laminin remodeling prior to satellite cell emergence
Previous work had reported that satellite cells emerged from their sublaminar position after 24 hr, 13 and yet we had detected migration prior to this event (Fig. 1B and Supplementary Movie 1A). Therefore, we carried out a detailed analysis of the emergence of satellite cells from under the basal lamina. Immediately after fiber isolation, we were able to detect the basal lamina over the Pax7-expressing cell (Fig. 1C). Importantly, there was a uniform layer of laminin over the satellite cell and the myofiber. Subsequently, we found that the distribution of laminin changed to become concentrated around and above the satellite cell—a phase lasting approximately 18 hr (Fig. 1D–G, red arrowheads, and Fig. 1J, dorsal view). From 18 hr onward, holes were noted in the concentrated basal lamina (Fig. 1H, red arrowhead). Satellite cells appeared to emerge from these holes in the basal lamina (Fig. 1K, green arrowhead). Emergence of the satellite cell often left a scarred basal lamina. By 24 hr, most of the satellite cells that had emerged had moved from their original resting place and were located over intact basal lamina (Fig. 1I). Time-lapse microscopy was used to examine satellite cell migration before emerging from the basal lamina. During this period, satellite cells displayed an elongated morphology and formed lamellipodia like projections ahead of the cell (Supplementary Movie 1A).
Migration characteristics of satellite cells
Satellite cells showed a remarkable ability to change direction and speed. We quantified movement by measuring the linear migration and found that straight-line movement was a rare event (see Fig. 4B). One key observation was that satellite cells stopped moving during division (Supplementary Fig. 1B). However, after division, daughter cells showed one of three types of behavior. One or both cells did not move, or both daughter cells moved together or, as most commonly found, the daughter cells moved independently of each other (as shown in Supplementary Fig. 1). Our time-lapse movies also revealed an interesting feature about cell division. We found that most cells divided at right angles to the muscle fiber so that for a brief period of time one cell appeared to be on top of the other (Supplementary Figure 1C,G).
Blebbing of Satellite cells during migration
To identify which mechanism regulated satellite cell migration, we generated electron micrographs at differing time points after fiber isolation. SEM images clearly showed the emergence of satellite cells from a hole in the basal lamina (Fig. 2A). Interestingly, at these time points, the surface of the cells appeared to be smooth. This morphology is greatly altered following emergence of the satellite cells from the basal lamina, when satellite cells began to display membrane blebs (Fig. 2B). The distribution of the blebs was polarized to one end of the cell. Blebs were still prominent following 48 hr and 72 hr of culture (Fig. 2C,D). Furthermore, blebbing was confined solely to the suprabasally positioned satellite cells and did not occur on the myofiber. We measured bleb size during the three phases of migration after emergence from under the basal lamina and found that bleb size remained fairly constant (0.69 μm at 24 hr, 0.67 μm at 48 hr, and 0.85 μm at 72 hr). These extensions are of significant length and represent protrusions of approximately 7% of overall satellite cell size.

Satellite cells display an amoeboid-like shape following emergence from the basal lamina.
Highly dynamic nature of satellite cell blebbing
To confirm that blebbing is the main mechanism by which satellite cells move, and that our SEM images were not the result of satellite cells undergoing apoptosis, we carried out two experiments. First, myofibers were cultured in the presence of a caspase inhibitor (Z-VAD-FMK), for 48 hr, a time point when satellite cell migration was at its most rapid, and myofibers were processed for SEM. SEM images revealed that satellite cells cultured in the presence of caspase inhibitor continued to bleb profusely (Fig. 2E). Time-lapse phase-contrast microscopy was used to visualize single satellite cells moving along the surface of myofibers, and to determine whether the blebbing on the satellite cells was a dynamic process. These movies, taken 30–48 hr following myofiber isolation over a period of 20–60 min, showed in all cases (n = 20 tracked cells) that cells formed blebs and did not produce obvious lamellipodia (see Supplementary Movie 1B). There was a significant difference in the morphology displayed by satellite cells that were positioned either on top of the fiber or under the basal lamina. Satellite cells positioned on the surface of myofibers were spherical in contrast to the elongated shape when localized under the basal lamina. One of the fascinating features of blebbing is the highly dynamic nature of plasma membrane protrusion and retraction. 25 We found that on the surface of satellite cells, blebs were rapidly extended (15 sec) and retracted at similarly high speeds (30–45 sec) with the whole process taking place in approximately 60 sec (Fig. 5B, below, and Supplementary Movie 1). Blebbing occurred over the entire surface of the satellite cells but was more pronounced at the leading region (Supplementary Movie 1B).
Regulation of satellite cell migration
Here we first aimed to establish a role for NO as a mediator of satellite cell migration. NO is produced by a family of enzymes called NOS. We found a dynamic expression of eNOS and iNOS in satellite cells. eNOS was not detected in quiescent satellite cells but was present following activation. Furthermore, its expression was found in both the cytoplasm and at the cell membrane. Moreover, it was differentially expressed at high and low levels in cells of the same cluster or progeny (compare red and blue arrowheads in Fig. 3B). Additionally its expression at the membrane was, at times, polarized to one end of the cell (Fig. 3B red arrowheads). In contrast, iNOS expression localized to the nuclei of the satellite cells and to a lesser degree that of the myonuclei (Fig. 3A).
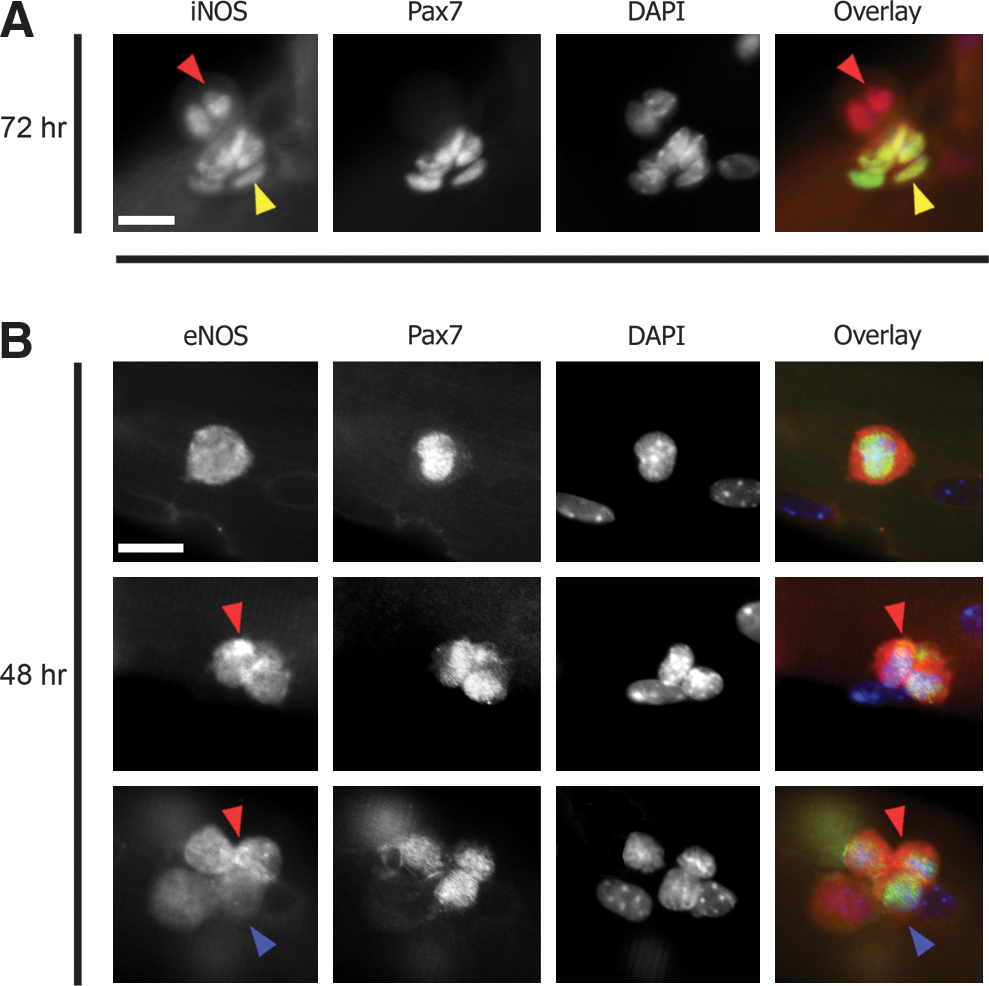
Inducible nitric oxide synthase (iNOS) and endothelial nitric oxide synthase (eNOS) are expressed during the satellite cell migratory phases.
The role of NO and Wnt signaling in satellite cell migration
We investigated whether the NO and Wnt signaling pathways were able to regulate satellite cell migration. We took advantage of inhibitors that prevent signaling mediated by these molecules. L-NAME acts a competitive inhibitor of all forms of NOS and prevents the synthesis of nitric oxide. JNK/SAPK, c-Jun N-terminal kinases/stress-activated protein kinase (c-jnk), has been shown to regulate cell migration 28 and is a component of the noncanonical Wnt signaling pathway and can be inhibited using SP600125. 29
First, we incubated isolated muscle fibers at differing phases of migration with either L-NAME or SP600125 and measured migration parameters of tracked satellite cells (n = 7 cells tracked for L-NAME and 11 cells tracked for SP600125, respectively). L-NAME attenuated the rapid cell movement displayed by untreated cells (Fig. 4A). In contrast to control cells, which displayed an increased change in migration speeds over time, L-NAME treatment rendered the cells slow moving throughout the course of observation. Treatment with SP600125 had quite a different outcome on satellite cell migration. We found that during the first 18 hr there was no difference in the migration rates compared to untreated cells (Fig. 4A). However after 18 hr, SP600125 brought approximately a 30% average reduction in migration speed in all cells, which carried on reducing over time (Fig. 4A).
Next we determined whether the NO or PCP signaling cascades regulated directionality of migration by calculating the straight-line migration characteristics of satellite cells treated with either L-NAME or SP600125. Our results showed that although both inhibitors decreased the velocity of migration, they did not alter the directionality (Fig. 4B).
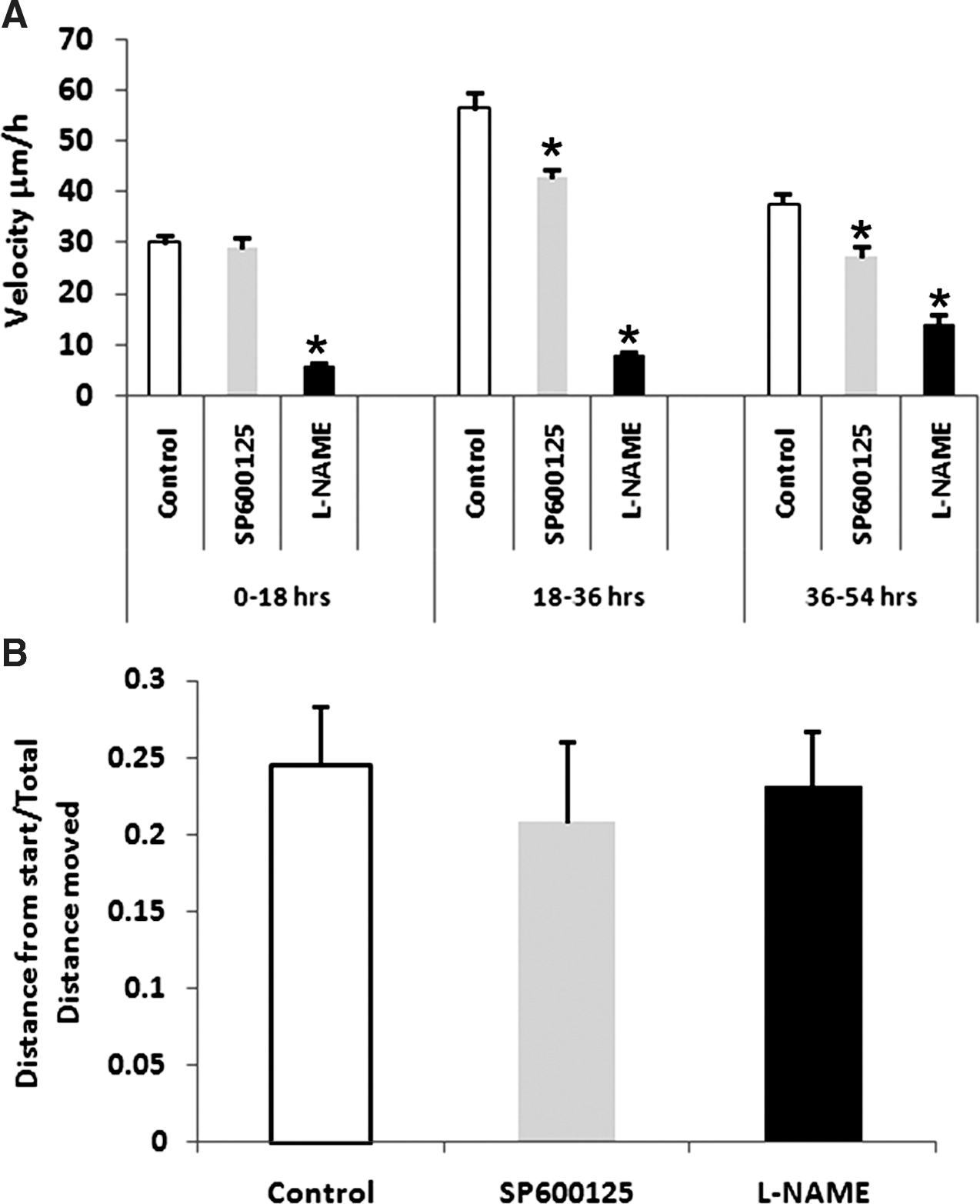
Nitric oxide (NO) and planar cell polarity (PCP) pathways play key roles in satellite cell migration.
The effect of inhibiting PCP and NO signaling on satelite bleb formation
We next examined whether the PCP and NO inhibitor compounds altered bleb morphology. Inhibition of the NO pathway with L-NAME led to very few blebs developing on satellite cells at all time points examined (Fig. 5C), in contrast to untreated cells at 24 hr that showed polarized blebbing events. At 48 hr when control cells were covered in blebs, the number on L-NAME–treated fibers was again considerably lower. Furthermore, the morphology of blebs in L-NAME–treated samples was altered compared to untreated fibers; L-NAME caused blebs to be much broader. A similar bleb profile was seen at 72 hr in the presence of L-NAME (Fig. 5C).
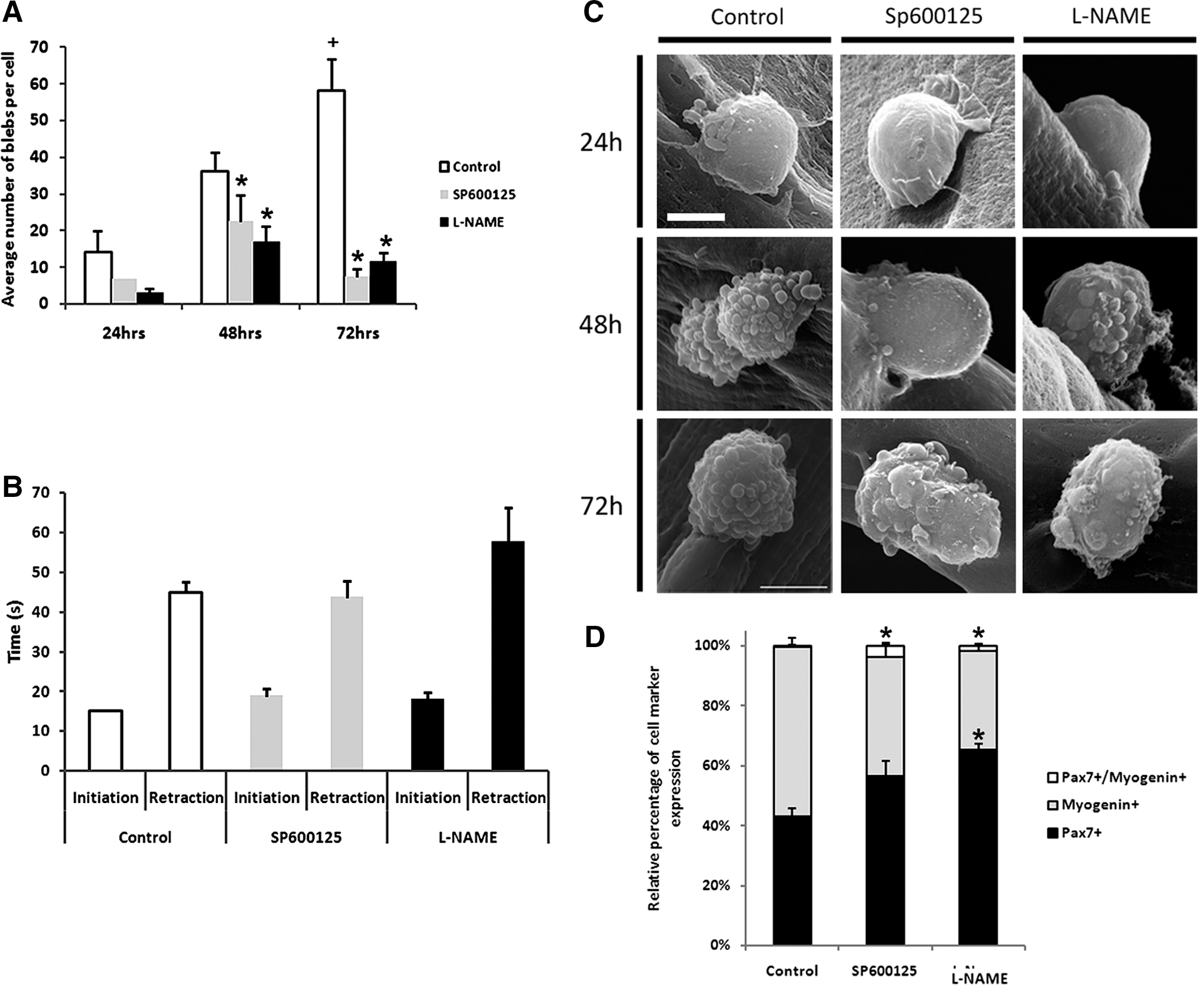
Reduced satellite cell migration velocity induced by SP600125 or N-nitro-
Treatment with SP600125 also brought about a dramatic alteration in bleb number and morphology. At all time points, there were fewer blebs in SP600125-treated samples. This was particularly evident at 72 hr, when the bleb number was severely decreased, and those blebs that were present were large and flat compared to those on untreated cells (Fig. 5C).
We quantified the number of blebs that formed at differing time points in the presence of the two inhibitors on satellite cells in SEM images. In untreated cells, the number of blebs had increased approximately four-fold by the 72-hr experimental period (Fig. 5A, n = 3, 21, and 14 cells analyzed at 24, 48, and 72 hr, respectively). In contrast, treatment with either L-NAME or SP600125 resulted in fewer blebs at any comparable time point. Furthermore, there was a small increase at 48 hr compared to 24 hr, but that increase was only transient and at 72 hr the bleb number was similar to that at 24 hr (Fig. 5A, n = 3, 10, and 15 cells for SP600125- and 4, 10, and 33 cells for L-NAME–treated cells at 24, 48, and 72 hr, respectively).
To confirm these effects of L-NAME and SP600125 on bleb formation, high-power time-lapse movies of single cells treated with both L-NAME and SP600125 were obtained at 48 hr following myofiber isolation. A key observation over short periods was that the satellite cells treated with either L-NAME or SP600125 did not move but nevertheless did undergo blebbing. L-NAME–treated fibers revealed two features. First, the number of blebs that were seen developing on the cells was reduced and, second, that they were larger when they did form (albeit not as large as those observed in SP6001250-treated cells). Examination with high-power, time-lapse, phase-contrast microscopy of SP600125-treated cells revealed that cellular blebs were increased in size dramatically and reduced in number, and the cell displayed disorganized erratic small movements from side to side as opposed to small organized bleb formation observed on control cells (see Supplementary movie 1C,D, respectively).
Actin cytoskeletal remodelling and ROCK1 activity are required for satellite cell blebbing
We explored the role of actin cytoskeletal remodeling in bleb formation by first monitoring blebbing in satellite cells at 48 hr following fiber isolation. Then we watched each cell following the introduction of cytochalasin D, an inhibitor of actin polymerization. 30 We found that in all cases examined in the presence of cytochalasin D, cells extended their blebs; however, the blebs then failed to retract (Fig. 6A–C).
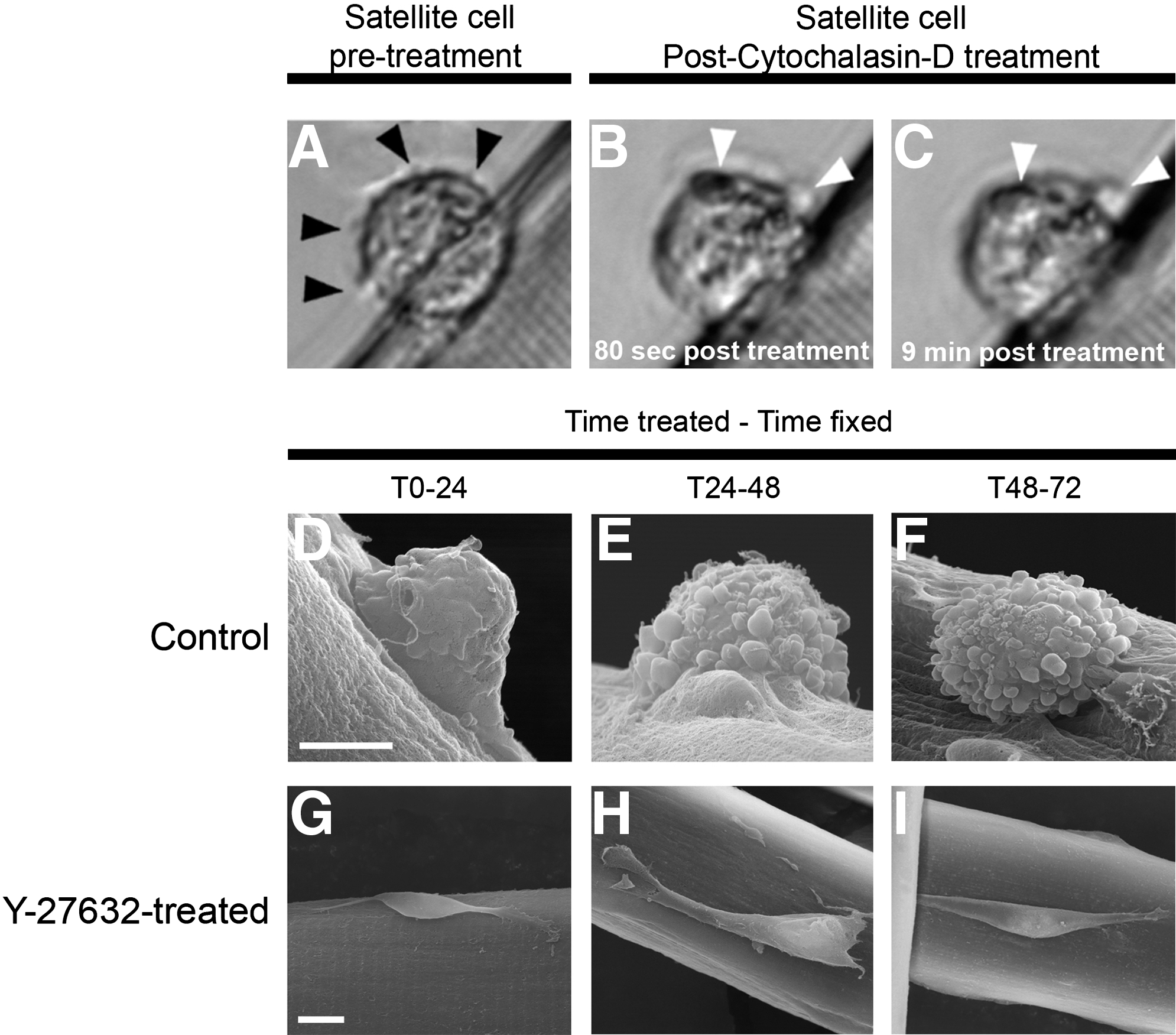
Actin polymerization is required for the maintenance of bleb formation on satellite cells.
To investigate the role of ROCK in satellite cell bleb formation, we incubated myofibers in the presence of the ROCK inhibitor Y-27362 at three separate time points—T0–24, T24–48, and T48–72 hr, respectively—and carried out SEM analysis to establish satellite cell morphology. Our data showed that in the presence of Y-27362, all satellite cells viewed under SEM at all time points analyzed adopted a smooth surface and produced lamellipodia-like structures (Fig. 6D–F and Supplementary movie 1E). We measured the speed of satellite cell migration following Y-27362 treatment between T48 and T56 hr following myofiber isolation and found that velocity was reduced from 41.3 μm/h in control conditions to 17.2 μm/hr in the presence of Y-27632 (see Supplementary movie 1E).
Inhibition of NO and PCP on satellite cell development
We next determined whether the decrease in migration rates seen following the introduction of L-NAME or SP600125 was due to an acceleration in the differentiation program. To this end, we cultured fibers in either inhibitor and profiled the progeny after 72 hr of culture for myogenin as a marker for differentiation and for Pax7 as a marker of the self-renewing myogenic stem cell population. We found that introduction of either inhibitor reduced the level of differentiation (Fig. 5D). In conclusion, we can state that the decrease in migratory rates and the loss of blebbing is not due to the ability of the inhibitors to drive premature differentiation.
NO and PCP signaling and migration of cultured myoblasts.
We finally examined whether NO or PCP cascades regulated migration of muscle cells in vitro. To this end, we examined the effects of the inhibitors on the movement of C2C12 cells. This myoblast cell line was grown under proliferation-maintaining conditions at all times during the experiment and introduction of either L-NAME or SP600125 resulted in a significant decrease in the rate of migration (Supplementary Fig. 2). One difference was that SP600125 in these cultured cells caused a greater reduction in migration compared to L-NAME. We also found that simultaneous addition of both compounds resulted in a greater degree of inhibition of migration than either compound on its own (See Supplementary movie 1F).
Discussion
Plasma membrane blebs are dynamic cytoskeleton-regulated structures that have long been known to be involved in apoptosis, but their role in cell migration has been overlooked until recently. The ability of cells to use membrane blebs for their motility has lately become an area of intense interest following the observation that primordial germ cells in vivo migrate exclusively through a bleb-mediated mechanism. 31 Furthermore, new studies have shown that tumor cells can adopt a bleb-driven, amoeboid-type of movement through extracellular matrix. 32,33 This bleb-driven migration produces faster migration speeds than the well-studied lamellipodia-driven process. In this study, we provide the first evidence that resident muscle stem cells use this newly discovered mechanism for migration along myofibers.
An essential feature in the regeneration process is thought to be the requirement of satellite cells to migrate to the point of lesion, 7,8 although most studies on satellite cell migration have been performed using myoblast cell lines in culture. 11 –15 Recently, primary satellite cells moving on the surface of myofibers have been observed using time-lapse microscopy, and a number of unexpected properties were reported in an important study by Siegel et al. 16 In particular, satellite cells on fibers showed a rapid migration compared to myoblast cell lines. Furthermore, numerous differences were noted between satellite cells and cultured muscle cell lines. The most significant finding of our study is that it represents the first report of satellite cells moving on their in vivo substrate in an amoeboid or bleb-dependent mechanism, which is independent of lamellipodia formation. In addition to this startling finding, we also confirm the rapid migration of satellite cells along myofibers and establish that the cell movement is more rapid than any other cell line tested, including the epithelial MCF10A cell line, which is widely used in migration studies due to its motile properties. 34,35 Previous studies have suggested that satellite cell asymmetric division occurs in, and is regulated by the stem cell niche environment, and occurs beneath the basal lamina of the myofiber. 24 However, our immunofluorescence, confocal, SEM, and time-lapse analysis all suggest that the initial division of all observed satellite cells occurs away from the original niche and only once the cells have emerged from beneath the basal lamina. Moreover, these experiments revealed that satellite cells were always stationary prior to and during division, and images captured during these phases using SEM showed no membrane blebbing, again evidence for blebbing being involved in migration. Interestingly, we have observed that the initial satellite cell division always occurs in the direction away from the myofiber surface, suggesting indeed that if asymmetric divisions are occurring, they may be regulated by an interaction with the outside, and not the inside, of the basal lamina. Further studies addressing this important issue are currently underway.
We also report that satellite cell migration starts even before the cells emerge from the basal lamina, with cells moving beneath the basal lamina at a relatively slow speed (27 μm/hr), before stopping, emerging onto the surface and then start moving again at a much higher speed (50 μ/hr). Given that cells emerge onto the surface of the myofibers with smooth membranes showing no sign of blebbing and that there are significant differences in the speed of migration on the surface compared to beneath the basal lamina, it is possible that the satellite cells employ different mechanisms of migration depending on the environmental context in which they find themselves. Our time-lapse movies support this notion by capturing images of elongated cells displaying lamellipodia like extensions in the direction of cell movement. The movement of satellite cells beneath the basal lamina may be a response to activation of the cells that leads them to search out potential exit locations to emerge onto the surface of the fiber, suggesting that their quiescent niche may not be optimum to enable exit from the basal lamina.
Perhaps the most startling behavior observed is the formation of blebs on the surface of actively migrating satellite cells. In the early stages after satellite cells emerge onto the fiber, SEM reveals that the emerging satellite cells have smooth surfaces. In some cases the beginnings of polarized blebbing can be observed shortly after emergence while at time points in which the cells are most motile (24–48 hr) all of the satellite cells, with the exception of a few that had recently undergone cell division, showed very substantial membrane blebbing. High-magnification time-lapse imaging of satellite cells moving along fibers also revealed substantial membrane blebbing occurring in all motile cells. Blebbing motility, also sometimes known as amoeboid motility, has recently begun to attract significant interest as an alternative mechanism of cell movement to that based on the more widely studied lamellipodia-based motility. A number of recent studies have suggested that bleb-based cell migration may be more common than is currently appreciated and may be a particular feature of cell migration in vivo (for reviews, see refs. 26 and 36). Live studies in zebrafish embryos have shown that primordial germ cells use blebs to migrate, 31 whereas studies in tumor cells migrating through extracellular matrix (ECM) have revealed that tumor cells can employ a blebbing based mechanism of motility, distinct from lamellipodia-based migration, to migrate through the ECM in a mechanism that does not require total matrix degradation. 33,37,38 We now report the first instance of bleb-directed cell migration occurring in adult mammalian cells on their native substrate performing a physiological role.
Bleb formation appears to follow a well-defined life cycle that can be divided into three phases—initiation, expansion, and retraction. The total life cycle of a bleb has been documented to last for 1–2 min and produces membrane blebs that expand up to 2 μm in size. The dynamics of blebs formation in satellite cells appears to conform to these phases and have a life cycle of around 1 min and can grow to a size of 0.7 μm. In addition to observing blebbing occurring in migrating satellite cells, we have also observed significant blebbing occurring in entire clusters of cells. Because these clusters are also capable of rapid migration along the myofiber, it is interesting to note that they appear to employ the same mechanism of migration as the individual cells.
It has been proposed that blebbing-based motility may provide a number of advantages to cells moving through complex three-dimensional environments such as those found in muscle. In these environments, lamellipodia may have no obvious surface to follow; bleb protrusion can occur in any direction, making rapid changes in course possible, and blebs may provide a simple means of adaptation to the shape and structure of the local environment. In addition, bleb expansion is faster than lamellipodial protrusion, suggesting that this may allow more rapid migration of cells. In tumor cells, it has been proposed that the acquisition of amoeboid or blebbing-based motility is associated with a greater ability to move and to colonize distant sites in vivo. 39 These observations are particularly important in light of our results, which show that satellite cells can move along myofibers significantly faster than almost every cell line tested capable of moving across a two-dimensional surface using lamellipodia. In the context of muscle regeneration, bleb-based migration of satellite cells may provide a mechanism for rapid repair of damaged sites on muscle fibers. We suggest that a number of behavioral characteristics of satellite cells are governed by their local environment. An elegant study by Gilbert and colleagues has recently shown that substratum mechanical properties regulate the self-renewal capacity of satellite cells. 40 We suggest that the mechanism used by satellite cells for migration is also dictated by the substratum. We have examined satellite cells maintained on myofibers in conditions that support proliferation and have shown that they form blebs. In contrast, others have taken satellite cells away from the myofiber, cultured them in differentiation medium, and shown that they form lamellipodia. 41
NO is a gaseous signaling molecule that regulates diverse physiological and cellular processes. It promotes cell migration in numerous cell types. Treatment of isolated myofibers with the general NOS inhibitor L-NAME led to significantly reduced migration and to impaired bleb formation. Blebbing in L-NAME–treated cells was markedly reduced, with fewer, blebs occurring, suggesting that NO may play a role in the regulation of bleb formation. The role of NO in cell migration has exclusively been studied in cells employing lamellipodial motility rather than bleb-based motility, and many of the molecular targets for NO have been identified in these cells, including the integrins αVβ3 42 and β1. 43 It is possible that these NO-regulated proteins may also be involved in regulating bleb formation, or it may be that there are further targets to be identified that are specific to blebbing based motility.
Previously, we have shown that Wnt signaling plays a role in satellite cell proliferation dynamics 21 and wanted to determine the role noncanonical Wnt signaling might play in regulating satellite cell migration. JNK/SAPK has been shown to regulate cell migration 28 and is a component of the noncanonical Wnt signaling pathway. Using a selective inhibitor of JNK, a key component of the nonconical Wnt pathway, we have shown that, like NO, it plays an important role in regulating both bleb formation and satellite cell migration. Little is known about the regulation of bleb formation in migrating cells, and, while much work is needed to fully elucidate these regulatory mechanisms, we have established a role for both NO and noncanonical Wnt signaling in this process.
One important feature of our studies was that although both NO and the PCP pathway regulated the velocity of migration, neither had any impact on directionality of movement. Comparisons are invited between our findings and those of Siegel and colleagues. 16 In the latter study, the influence of guidance molecules was examined on migration velocity. They found that hepatocyte growth factor (HGF) altered migration trajectory but not velocity. Assimilating the findings of Siegel et al. 16 and those in the present study, it appears that the mechanisms regulating cell migration and guidance are independent processes.
The observation of these behaviors of satellite cells, particularly their reliance on bleb formation for migration, is fundamental to developing therapies for muscular dystrophies or aging-related muscle weakness, because bleb-based motility uses completely different pathways and regulatory mechanisms from lamellipodial-dependent migration. Studies using the myoblast cell line C2C12, for example, have suggested that N-WASP and WAVE2 activated downstream of phosphoinositide 3-kinase (PI3-kinase) play an important role in satellite cell migration. 13 However, these cells move in a lamellipodial-dependent mechanism and as such are a poor model for studying the regulation of migration in satellite cells. Our analysis of C2C12 migration demonstrates that L-NAME has less of an effect on these myoblast cells, whereas the PCP pathway plays a greater role in their movement, the opposite of our findings in primary adult satellite cells. These data support the hypothesis that the balance of pathways regulating bleb-based and lamellipodia-based migration is different and suggests separate mechanisms are in place. We show that inhibiting a key regulator of amoeboid movement causes cells to no longer display blebs but instead lamellipodia. This work supports the hypothesis that blebbing- and lamellipodia-based mechanisms act in an antagonistic manner. 44
Blebbing motility represents an important and exciting new avenue of research in the field of cell migration, especially of satellite cells, yet remains poorly studied compared to lamellipodial motility. This may be because its role in cell migration in vivo is underappreciated, possibly due to the technical difficulty of studying this type of migration, because it is only apparent in complex three-dimensional matrices or living tissues.
Footnotes
Acknowledgments
We would like to thank the University of Reading (K.P. and A.O.), the BBSRC (K.P. and H.C.H.), Systems Biology Laboratory UK (K.P. and H.C.H.), and the British Heart Foundation (FS/08/056; P.D. and A.P.) for generous funding to allow the completion of this study, and Dr. Chris Stain for assistance with SEM analysis.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.