
Abstract
The emergence of immune disorders in aging is explained by many factors, including thymus dysfunction, decrease in the proportion and function of naïve T cells, and so forth. There are several approaches to preventing these changes, such as thymus rejuvenation, stem cells recovery, modulation of hormone production, and others. Our investigations of heterochronic parabiosis have shown that benefits of a young immune system, e.g., actively working thymus and regular migration of young hematopoietic stem cells between parabiotic partners, appeared unable to restore the immune system of the old partner. At the same time, we have established a progressive immune impairment in the young heterochronic partners. The mechanism of age changes in the immune system in this model, which may lead to reduced life expectancy, has not been fully understood. The first age-related manifestation in the young partners observed 3 weeks after the surgery was a dramatic increase of CD8+44+ cells population in the spleen. A detailed analysis of further changes revealed a progressive decline of most immunological functions observable for up to 3 months after the surgery. This article reviews possible mechanisms of induction of age-related changes in the immune system of young heterochronic partners. The data obtained suggest the existence of certain factors in the old organisms that trigger aging, thus preventing the rejuvenation process.
Introduction
Accumulated data demonstrate an inverse relationship between immune status, response to vaccination, and health, on the one hand, and age, on the other. These data suggest that the immune system becomes less effective with advancing age, and this decline is clinically relevant. 5 The immune system plays an important role not only in immunity (immune protection) but also in the regulation of the tissue homeostasis (immune physiology). Immune physiology shifts can result in pathology such as autoimmune, metabolic, and degenerative diseases and in the infertility and intrauterine growth retardation, fetal morbidity, and mortality. 6 This is why restoration of the immune function that has been reduced by age is so important.
The immune system is a convenient object for exploring regeneration. There is an intriguing concept that if stem cells can be induced to make new lymphocytes or antigen-presenting cell (APC) subpopulations, these fresh cells may be able to rejuvenate the function and restore immune system defects. 7 Mayak et al. have demonstrated that age-dependent changes in hematopoietic niche cells can be reversed by exposure to young blood. 8 These results disagree with the data obtained in our laboratory (A. Sidorenko et al.), likely due to a difference in the duration of parabiosis. 9 In this paper, we will discuss research investigating the immune system in heterochronic parabionts and the dynamics of its age-related changes.
Materials and Methods
Mice and parabiosis
CBA/Ca, FVB, and FVB.Cg-Tg(GFPU)5Nagy/J mice were bred and maintained at the Institute of Gerontology Research Animal Facility. For the parabiosis, animals (males or females with matching controls) were typically joined at 2–3 (young) and 22–23 (old) months of age. The parabiosis surgery was performed exactly as described previously 10 and in accordance with the guidelines established by the Institute of Gerontology Administrative Panel for Lab Animal Care. At 3, 6, weeks and 3 months after the initiation of parabiosis, mice were euthanized and subjected to tissue analysis. All parabiont pairs with tumors were excluded from the study. To confirm efficient blood mixing in parabiotic mice, the percentage of green fluorescent protein–positive (GFP+) cells among leukocytes in the thymus, spleen, and bone marrow (BM) was analyzed in each wild-type animal connected to the GFP+ parabiotic partner.
Isolation of cellular populations
Spleens and thymuses were removed from mice and disrupted by mechanical dissociation. Red blood cells (RBCs) were lysed from each cellular suspension and then used for flow-cytometric analysis. BM was flushed from the femurs of mice with Hanks' balanced salt solution (HBSS; BioTest Laboratory, Ukraine) supplemented with 2% fetal calf serum (FCS; Sangva, Ukraine). RBCs were lysed during a 3-min incubation in 0.15 M ammonium chloride and 0.01 M potassium bicarbonate solution on ice.
Flow cytometry
The antibodies used in these studies included anti-CD8α/Lyt-2 (Texas Red conjugate, clone 53-6.7), anti-CD4/L3T4 (PE-Cy7, GK1.5), anti-CD3ε (fluorescein isothiocyanate [FITC] conjugate, clone C363.29B [YCD3-1]), anti-CD44-Pgp-1 (allophycocyanin conjugate, clone KM201) (PickCell Laboratories, Netherlands), monoclonal antibody (mAb) to FoxP3 (phycoerythrin conjugate, clone 3G3, Abcam), mAb to CD25 (PE-Cy5 conjugate, clone PC61.5, eBioscience), rabbit antibody to GFP (Santa Cruz), goat anti-rabbit immunoglobulin G (IgG) (tetramethyl rhodamine iso-thiocyanate [TRITC] conjugate, PickCell Laboratories, The Netherlands). Briefly, single-cell suspensions were resuspended in phosphate-buffered saline (PBS) with 2% FBS (Sangva, Ukraine) at 2×107 cells/mL. Cells were incubated with antibodies for 30 min on ice in the dark. Cells were washed in PBS and fixed with 1% paraformaldehyde in PBS. In the case of using antibodies to GFP and to FoxP3, cells were treated after fixation according to the protocol for intracellular staining. Events were analyzed using a CyAn™ ADP analyzer (DAKO). Data acquisition was performed using Summit Software. Dead cells were excluded on the basis of forward and side light scattering.
T cell proliferation assays
Splenocytes were cultured in the RPMI-1640 medium with glutamine supplemented with penicillin (100 U/mL), streptomycin (100 μg/mL), HEPES (25 μmol/mL, Sigma-Aldrich), and 10% heat-inactivated FCS (Sangva, Ukraine). In stimulation assays, the medium was additionally supplemented with phytohemagglutinin P (PHA-P; 10 μg/mL, Sigma-Aldrich). Splenocytes were seeded (2×106 cells in 0.2 mL of the culture medium) in 96-well round-bottomed plates (Sarstedt, Germany). The plates were incubated in a humid environment with 5% CO2 at 37°C for 3 days. Cell proliferation was evaluated by MTT assay. In brief, 10 μL MTT (5 mg/mL; Sigma-Aldrich) was added to each assay well and incubated for 2 hr, protected from the light. Formazan crystals were dissolved by resuspension in 100 μL of acidic isopropanol (0.4% (vol/vol) HCl in isopropanol), and the optical density (OD) was read on a Labsystems Multiskan Ascent ELISA analyzer using a test wavelength of 540 nm and a reference wavelength of 690 nm. Stimulation indexes were calculated by dividing OD values measured for stimulated cultures by those measured for unstimulated cultures (the mean of two replicates).
Phagocytosis assay.
Adherent cells were prepared immediately after splenocytes isolation by incubation of 15×106 cells in 35-mm dishes (Sarstedt, Germany) in the RPMI-1640 medium with glutamine, supplemented with penicillin (100 U/mL), streptomycin (100 μg/mL), HEPES (25 μmol/mL; Sigma-Aldrich), and 20% heat-inactivated FCS (Sangva, Ukraine) for 60 min at 5% CO2 and 37°C followed by three washes with RPMI/10% FBS. For latex bead uptake assays, latex beads (DiaeM, Russia) were suspended in the assay buffer at 2.5×108/mL and sonicated for 20 sec immediately before use. Adherent cells were incubated for 30 min with bead suspension at a final concentration of 2×108/mL and washed three times with HBSS/0.3% FBS. Dishes were fixed with 4% paraformaldehyde in PBS for 10 min, washed three times in PBS, and incubated for 5 min at room temperature with Romanovsky–Giemsa azure–eosin staining. The dishes were then dried and studied under a Zeiss light microscope. Assessment of the number of macrophages involved in the phagocytosis was performed by counting the number of cells with ingested particles for the total number of macrophages in each section. Obvious lymphocytes and granulocytes were excluded.
Statistical analysis
Data analysis using the Student t-test, one-way analysis of variance (ANOVA), and Mann–Whitney U-test was performed using Statistica 7.0 Software. A value of p<0.05 was used as a cutoff for statistical significance.
Results
In the first stage, we evaluated the status of the immune system in heterochronic parabionts 3 months after the operation. We assumed that if immune system aging is revealed through just a deficit of some component (serum factors, cells, etc.), then we could suppose that the migration of young stem cells in combination with young serum factors would contribute to the rejuvenation of the thymus. The restoration of the thymus, in turn, would help to restore the repertoire of peripheral T cells and regulatory T cells. And all of these factors would lead to the lymphoid organs and restoration of rejuvenation and immunological functions in the old parabiotic partners. The main processes that we expected from the young partners were the preservation of thymus functions, maintenance of stability of the peripheral T cells repertoire, and preservation of the immune system function at the level of young control animals. We did not find any signs of the thymus rejuvenation in the old partners. Also, we observed a significant reduction of the thymus weight in the young heterochronic partners without significant changes in thymocyte subpopulations (Fig. 1).
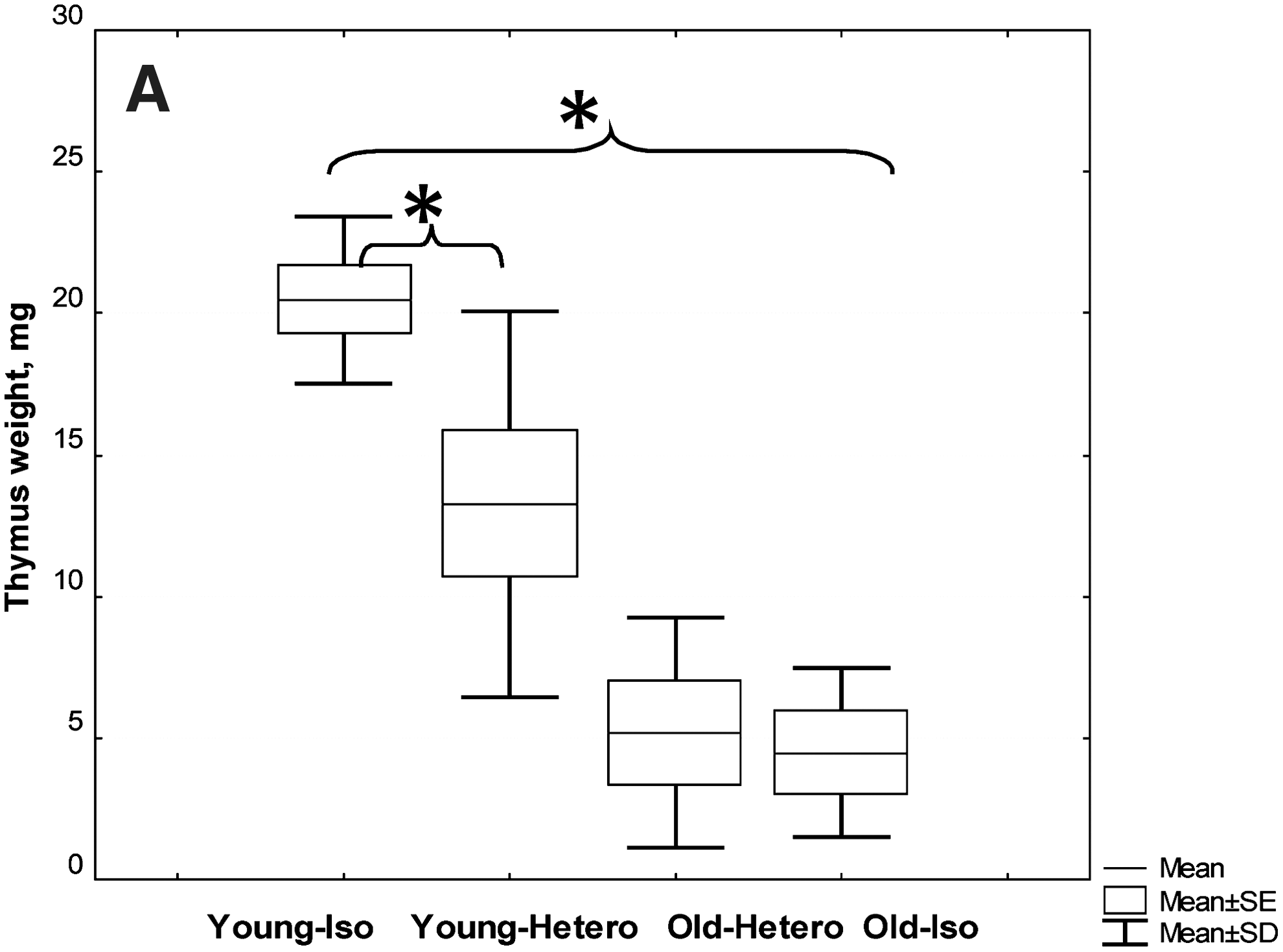
CBA/Ca female mice at 3 months after the parabiosis initiation were euthanized and subjected to thymus analysis. All pairs with tumors were excluded from the study. Thymus mass (

To check the status of the immune system, we used simple parameters that varied well with age, such as CD4 and CD8 T cell subpopulations in the spleen and proliferative response T cell to PHA in vitro. At first, we found that the proliferative activity of splenocytes in response to PHA after 3 months of the parabiosis did not change in the old heterochronic partners and decreased in the young ones. Similar changes were observed in the composition of T cell subpopulations in the spleen; the CD4/CD8 ratio was reduced. With regard to T memory–like cells (it should be noted that we used only one marker, CD44), their number increased in the young partners (Table 1). The most optimistic result in such a pessimistic picture was shown by regulatory T cells; their number decreased significantly in the old heterochronic partners to young level (Fig. 2).
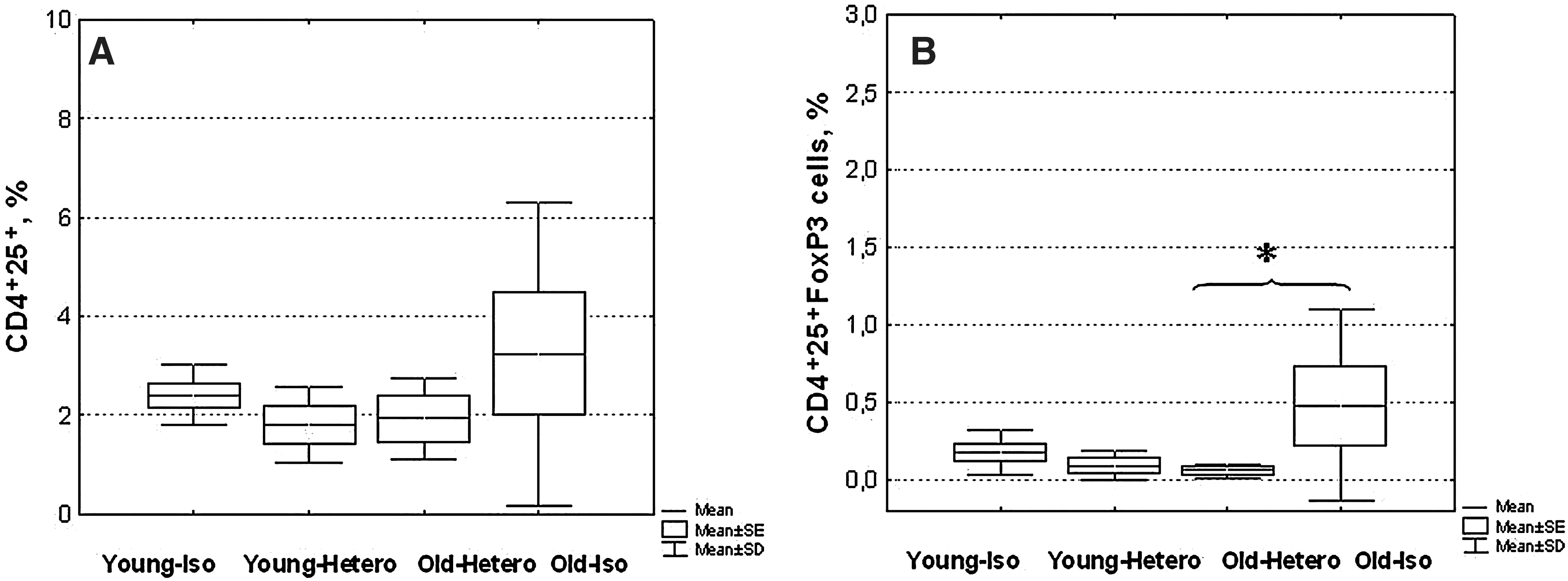
CBA/Ca female mice at 3 months after the parabiosis initiation were euthanized and subjected to splenocyte analysis. All pairs with tumors were excluded from the study. Numbers of CD4+25+ cells (
For the parabiosis, CBA/Ca male mice were typically joined at 2–3 (young) and 22–23 (old) months of age. At 3 months after the parabiosis initiation, mice were euthanized and subjected to immune parameters analysis. All pairs with tumors were excluded from the study.
p(t)<0.05 comparing young isochronic animals.
p(t)<0.05 comparing isochronic animals the same age.
p(t)<0.01 comparing isochronic animals the same age.
p(t)<0.01 comparing young isochronic animals.
To sum up, we found that the migration of young stem cells in combination with young serum factors did not contribute to the thymus rejuvenation. We did not note any recovery of the peripheral T cell repertoire, only normalization of regulatory T cells. And all of these factors did not lead to the restoration of immunological functions in the old parabiotic partners. The main changes observed in the young partners were: A decrease of the thymus mass, changes in the peripheral T cell repertoire, and a decrease of PHA-stimulated lymphocyte proliferation. We observed changes in the immune function of the young heterochronic partners, which were similar to aging. Therefore, we obtained an accelerated aging model for the immune system, which gives a unique opportunity for tracing the emergence of age-related changes in an accelerated mode.
For tracing of age-related changes animals were operated on at different times with intervals of 3 and 6 weeks to produce parabiotic pairs with different periods of coexistence. It was found that the decrease in the proliferative activity of splenocytes in response to PHA during the animals' coexistence was observed only in the young heterochronic partners and only after 12 weeks of coexistence (Fig. 3A). The phagocytic activity of macrophages in young heterochronic partners' spleen decreased as slowly as PHA-stimulated T cell proliferation (Fig. 3B).
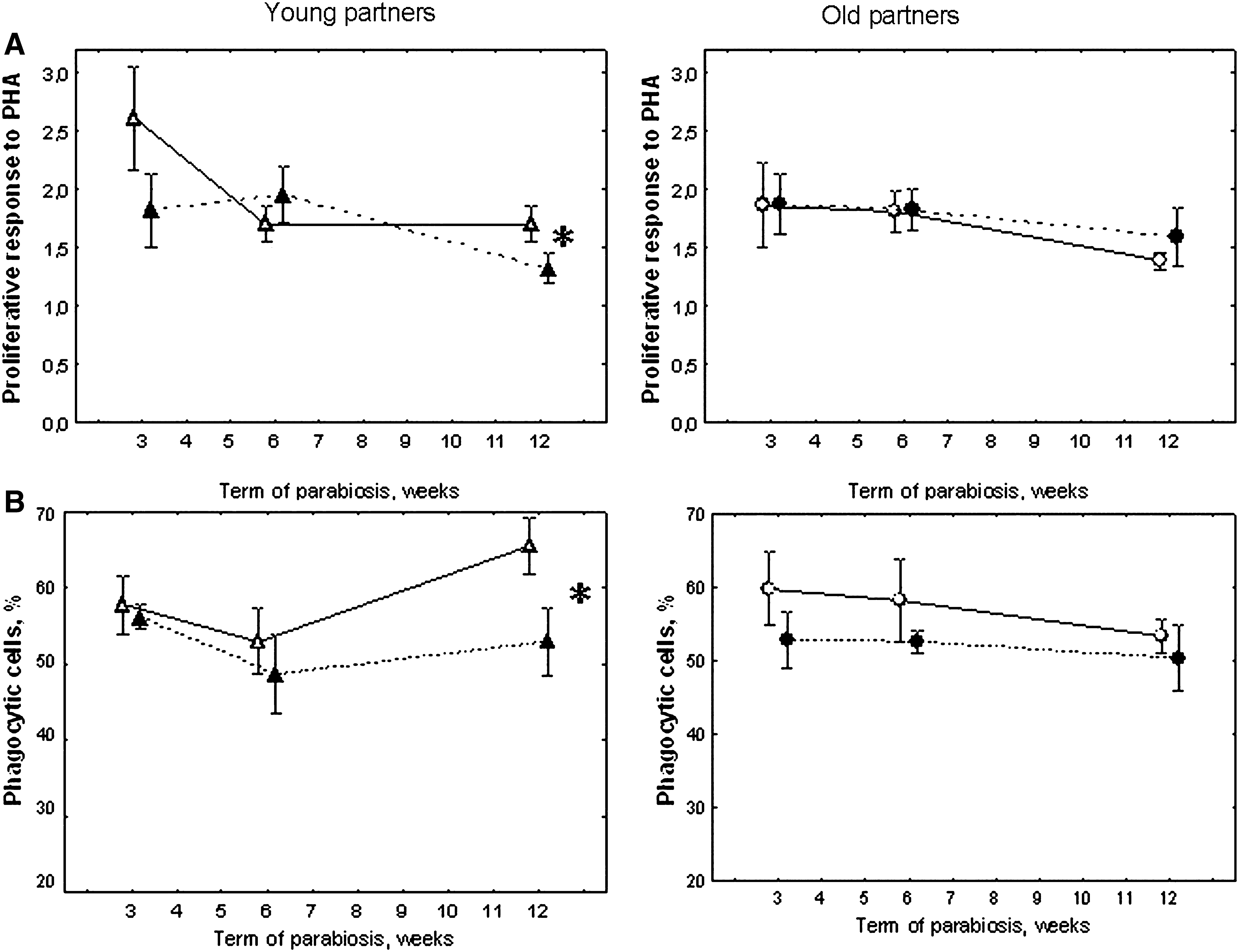
The dynamics of changes in the proliferative response of T lymphocytes to PHA in vitro (
The dynamics of changes in T cell subpopulations varied more notably. The first changes were observed after 3 weeks of coexistence and were accompanied by CD4+ cell decline in the young heterochronic partners' spleen (unpublished results). After 6 weeks, the CD4/CD8 ratio in the spleen was reduced as well (Fig. 4A). Similar changes were also observed in a population of memory-like cells. The most significant changes were induced in the population of CD8+44+ T cells. Their number was much higher in the spleen of the young heterochronic partners 3 weeks postsurgery and remained at this level throughout the observation period (Fig.4B). CD4+44+ T cells grew in numbers in animals of this group only after the 12th week of coexistence (Fig. 4C).
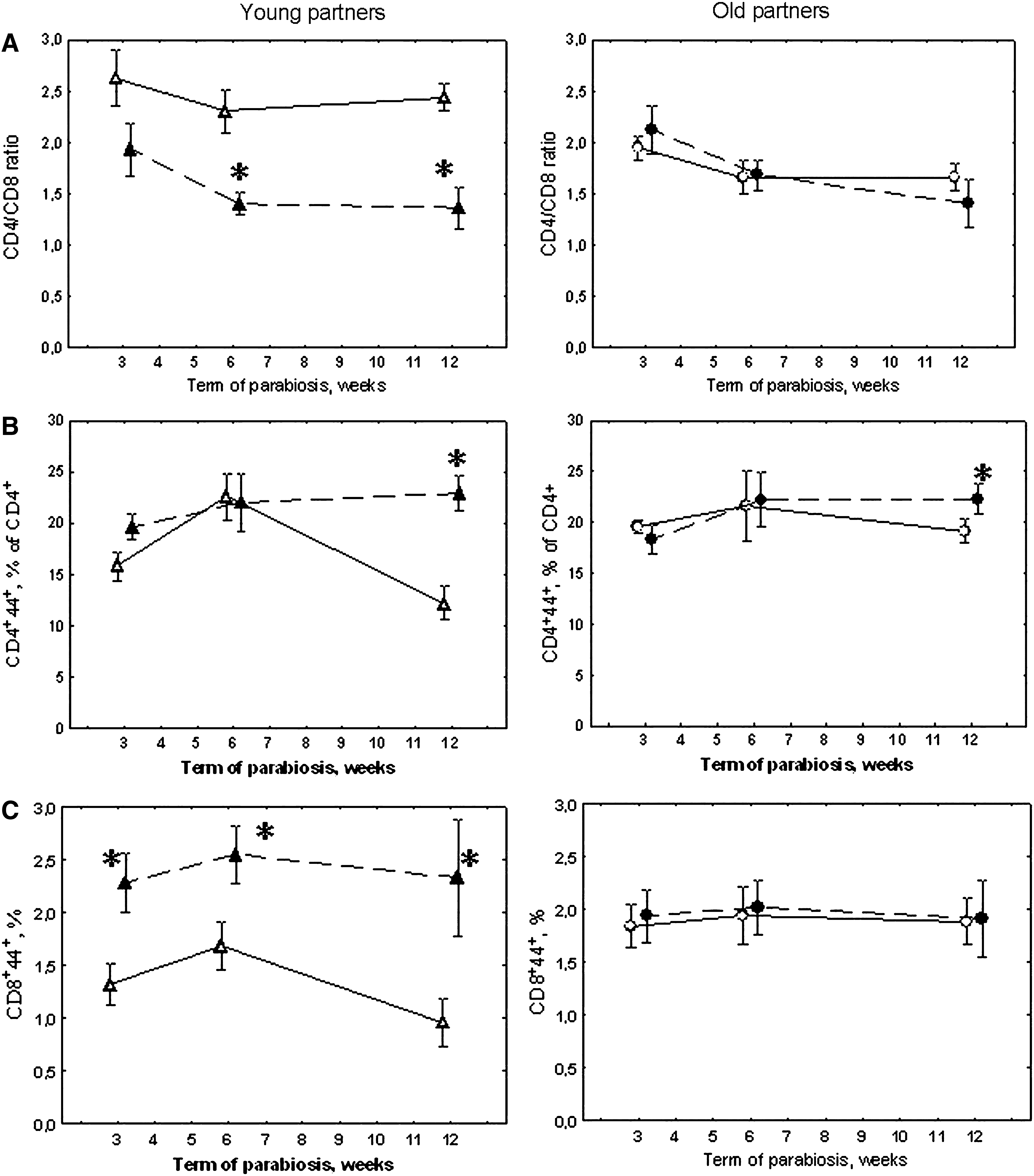
The dynamics of changes in the CD4/CD8 ratio (
As a result, we obtained a sequence of age-related changes of the immunological parameters in the young heterochronic partners. First, after 3 weeks of the co-existence, the T cell subpopulation was disturbed in the spleen; the number of CD8+44+ cells was increased. After 6 weeks, the CD4/CD8 ratio was decreased. By the 12th week, there was also a decrease in functional activity of T lymphocytes, which was a reduction of their proliferative response to PHA, and in number of phagocytic cells in the spleen.
We suggested two possible schemes for the development of the age-related changes: (1) The predominant migration of old T cells in the young organism; and (2) the migration of young T cells in the old organism, and the interaction with the old microenvironment with a subsequent increase of the T cell homeostatic differentiation, or young microenvironment aging.
We checked the lymphoid cell migration between heterochronic partners and did not find any difference in the number of donor cells in the thymus of heterochronic parabionts (Fig. 5A). We noted a significant increase of young SP CD4−8+ donor cells in the thymus of old partners as compared to the young isochronic controls (Fig. 5D). In the spleen of wild-type animals, we did not detect any difference in the number of donor cells in general (Fig. 5B), and in the number of CD4+ and CD8+ T cells in particular (unpublished data). We found a marked rise in the number of young donor CD8+44+ T cells among CD8 splenocytes in the old heterochronic partners relative to the young ones (Fig. 5E). Similar changes were also observed in BM cell populations, where a considerable increase of young donor CD8+44+ T cells among CD8 bone marrow cells in old heterochronic partners was found as compared to the young counterparts.

Cell migration analysis demonstrated the absence of differences in donor green fluorescent protein-positive (GFP+) lymphocyte number in the thymus () Mean; ( ) mean±standard error; (
) mean±standard error; ( ) mean±standard devation; WT, wild type; Y-y, young isochronic; Y-o, young heterochronic; O-y, old heterochronic groups.
) mean±standard devation; WT, wild type; Y-y, young isochronic; Y-o, young heterochronic; O-y, old heterochronic groups.
Discussion
Age-related defects in T lymphocytes are well appreciated. It has been shown that age-related atrophy of the thymus largely depends on the reduced number of thymocyte precursors in BM as well as changes in the hormonal balance, e.g., increasing of the sex steroids level. 11 –13 The reduced number of double-positive (DP) thymocytes and the increased number of double-negative (DN) thymocytes in the old thymus indicates an occurrence of disturbances in their differentiation. We expected that the common blood circulation that enriches old blood with young hormones and growth factors and provides recycling of young stem cells from young to old parabiotic partners would contribute to the regeneration of the old thymus. Indeed, we observed a tendency to increasing the number of DP thymocytes and decreasing the number of DN thymocytes in the old heterochronic partners. However, these changes were not dramatic, and the thymus weight did not increase in animals of this group, which suggests that a more complicated pathway should be implemented to rejuvenate the old thymus and chronic administration of young hematopoietic stem cells is insufficient.
A significant decrease of the thymus weight, along with the absence of considerable changes in thymic cell subpopulations, was found in the young heterochronic parabiotic partners within 3 months of coexistence with the old partners. Such rapid atrophy of the thymus in the young heterochronic partners may indicate a more complicated mechanism of age-related thymic atrophy than just a decline in the hematopoietic precursor migration from BM 9 and requires further research.
With a view to assessing the peripheral immune system, we evaluated indicators that most significantly change with age, especially subpopulations of T cells and proliferative activity of spleen lymphocytes in response to mitogens. 14 It was found that after 3 months of the parabiosis, subpopulations of splenocytes did not change in old animals, whereas the CD4/CD8 ratio in the spleens of young partners decreased dramatically. The number of cells with the memory phenotype in young partners' spleen increased up to the old level. In addition, the proliferative index of splenocytes from animals of this group reduced to the level of old animals. In fact, the heterochronic parabiosis model proved to be a model of accelerated aging of the young immune system because the studied parameters in the young heterochronic partners did not differ from the old animals' level.
In this case, we had a unique possibility to study the dynamics of age-related changes in the immune system of heterochronic parabionts, which could be an answer to the question about the trigger of these age-related changes. We examined the parameters of the immune system in parabiotic partners at different times after the surgery. It turned out that the first changes of the parameters studied occurred within 3 weeks after the surgery. These changes related to such values as the number of CD8+44+ memory-like cells in the spleen; their amount increased significantly in the young heterochronic partners. After 6 weeks of coexistence, the CD4/CD8 ratio of cells in the spleen was reduced in the young partners, and, after 12 weeks, functional activity of splenocytes was also reduced (the proliferative activity of T cells to PHA and the percentage of phagocytic macrophages in spleen).
As seen from the data obtained, the first age-related change occurs in the composition of T cells in spleen of the young heterochronic partners, which may indicate either a perturbation of lymphoid cell differentiation or the preferential migration of old lymphocytes in the young partners. To test this supposition, we investigated the migration of lymphoid cells between the parabiotic partners. It should be noted that the parabiosis establishes common blood circulation by surgical pairing; i.e., creates a surgical union of twin animals or parabionts. 15 Parabiosis using GFP+ parabionts was used to investigate the migration of dermal fibroblast/myofibroblast progenitors, 16 lung progenitors, 17 diabetic wound healing, 18 non-BM progenitors, 19 endothelial progenitors, 20 and BM stem cells. 21
Due to a great biologic interest in parabiotic cell populations, the kinetics of the blood flow and cell distribution between parabionts was investigated in detail by Gibney et al. 22 They found no evidence for a "parabiotic barrier" based on the cell size or surface characteristics, and all peripheral blood cell populations in this study reached equilibrium within 14 days. The whole blood fluorescence analysis indicated that the mean exchange flow rate was 16 μL/hr or 0.66% of the circulating blood volume per hour. 22
In our experiment, the main purpose was to examine the characteristics of the cell migration between animals of different ages. We established that the total number of donor lymphocytes did not differ in heterochronic partners. This fact demonstrates the equivalence of the migration of hematopoietic and lymphoid cells between partners of different age. However, in old animals a significantly higher number was found of young donor, single-positive CD4−8+ cells in thymus along with the number of CD8+44+ cells among the CD8+ cells of spleen and BM without affecting any other studied cell population.
As mentioned above, the first changes in splenocyte populations of the young heterochronic partners in our experiment appeared 3 weeks after the surgery and were reflected in the increasing number of CD8+44+ memory-like cells in spleen. This fact, together with the results of the lymphoid cells migration experiment, may indicate that an increase in the number of CD8+44+ cells in the young heterochronic partners may be caused by: (1) The predominant migration of young CD8+44+ cells in the old partner, (2) antigen-specific, or (3) homeostatic differentiation in young T cells in the old partner. However, on the basis of the results obtained, we cannot answer the question of what exactly the mechanism is leading to this increase.
Of note is that the population of CD8+44+ memory cells is short-lived and renews quickly. Parretta et al. found that both the doubling time and half-life of the CD8 memory population were 9 weeks versus 1 year for the naive subset. 23 These findings suggest that a higher turnover of memory CD8 T cells as compared with naïve CD8 T cells is mostly attributable to a higher proliferation rate in adult, mainly due to the homeostatic proliferation. 23,24 Yet by examining the rate of the in vivo T cell turnover in aged mice, a marked reduction in the turnover was revealed at the level of memory phenotype CD44hi CD8+ cells relative to young mice. On the basis of adoptive transfer experiments, the reduced turnover of aged CD44hi CD8+ cells reflected an inhibitory influence of the aged host environment, including APCs. 25,26 On the other hand, it was found that transferred young CD8 cells developed in young hosts were of a naïve phenotype, whereas the majority of those developed in the aged host displayed a memory phenotype with a high percentage of CD44hi cells. These results suggested that the aged microenvironment should have a significant impact effect on newly developed CD8 T cells. 27 Therefore, in our experiment, the rapid rise in the number of CD8+44+ cells in the young heterochronic partners may result from mutual influence of two environments—young for proliferation, and old for memory phenotype CD8 cells differentiation.
In our study, apart from increasing the number of CD8+44+ memory-like cells in the spleen of the young heterochronic partners, there was a reduction in the CD4/CD8 ratio, as well as a decrease in T cell proliferative activity in vitro. This fact is related to the emergence of age-related changes in the immune system and may be associated with thymus atrophy and impact of old APCs. 7,28 Another mechanism of age-related changes may be increasing of the number of regulatory T cells in the young heterochronic partners.
Regulatory T cells (Treg), a subset of CD4+ T cells, are critical for maintaining self-tolerance 29 and Treg function by decreasing the level of activation, proliferation, and cytokine production of effector T cells in mice and humans and by controlling stimulatory functions of dendritic cells. 30,31 Several researchers have reported that the frequency of FoxP3+CD4+ T cells significantly increases in multiple lymphoid tissues from aged mice. 32 –34 Also, it has been shown that FoxP3+ cells from aged mice have a greater in vitro suppressive activity on a per cell basis than their young counterparts. 32 During aging, thymic Treg output may decrease with a significant loss of thymic capacity to generate new T cells. Treg homeostasis has been shown to be sustained by alternative pathways, such as peripheral generation of Tregs or by greater resistance to the apoptosis. A Treg homeostasis imbalance would then predispose to immune dysfunction in aged individuals, explaining their higher risk of immune-mediated diseases, cancer, or infections. 35,36
In our investigation, the number of FoxP3+CD4+25+ T cells significantly increased in old isochronic animals. But, unexpectedly, the number of cells with this phenotype did not change in the young heterochronic partners. Meanwhile, in the old heterochronic partner, the number of FoxP3+CD4+25+ T cells decreased to the level of these cells in young animals. Reduction of the number of regulatory T cells in young heterochronic partners should theoretically lead to an increased immune response in animals. 32 Hence, in this case, some other mechanism should take place, because the magnitude of immune response to the sheep RBCs (SRBCs) in heterochronic partners stays at the level of old animals. 10 A possible mechanism that can explain the phenomenon obtained in the experiment is that germ-line FoxP3 mutation not only ablates Treg but also dramatically increases homeostatic T cell proliferation. 37 Homeostatic proliferation of T cells leads to the generation of effector/memory cells, which have the potential to cause harm to the host. The presence of Tregs resulted in a lower accumulation of T cells, enhanced apoptosis, and impaired differentiation to a cytokine-producing state. On basis of this fact, the authors concluded that Tregs play a major role in the control of homeostatic proliferation. 38 These facts, together with our data, suggest the possibility that a decrease in the number of regulatory T cells in heterochronic parabionts may provoke an increase in the homeostatic T cell differentiation and an increase of the number of T cells with a memory phenotype, and reduce their proliferative activity.
To this end, the assumption that immune system aging is an insufficiency of either its functions or factors (thymus, naïve T cells, hormones, and growth factors) seems groundless. The heterochronic parabiosis model has shown that the presence of common circulation between animals of different ages, which provides penetration of young stem cells, naïve T cells, hormones, and growth factors into an old body, does not lead to the rejuvenation of the old immune system. On the contrary, there is a change in the differentiation of young T cells that presumably enhances their homeostatic proliferation as impacted by old lymphoid niches. Reduction of the number of regulatory T cells may contribute to this process. This hypothesis needs thorough testing.
Footnotes
Author Disclosure Statement
No competing financial interests exist.