
Abstract
The use of growth factor composites from platelets has been introduced to many areas of clinical applications and studies. With the richest source of growth factors (GFs), beneficial effects have been shown on tissue regeneration and wound healing. However, animal and clinical studies have revealed inconsistent outcomes with the use of platelet-derived growth factors (PDGFs), which were likely due to variations in the presence and concentrations of GFs between various sources. Autologous PDGFs are considered to be safer, but they are limited by the feasibility of large-scale production to be used extensively in the acute phase, greater surface area, or general cosmetic applications. This study employed a simple process to obtain growth factor composites from activated platelets of porcine origin, namely skin renewal growth factors (SRGF). The functions of SRGF were subsequently evaluated on cultured human fibroblasts, keratinocytes, and melanocytes. Our data revealed that SRGF significantly promoted the proliferation of fibroblasts, accompanied by increased expression of collagens (types I, III, IV, and VIII) and proteoglycans. Diminished proliferation and arrested differentiation of keratinocytes were evidenced by the attenuated expression of laminin V and keratin 10. In addition, SRGF also suppressed the growth of melanocytes and reduced the expression of microphthalmia-associated transcription factor (MITF), tyrosinase, and paired box 3 (PAX3), which mediates melanogensis. Our results suggest that SRGF possesses beneficial properties and is a promising and cost-effective composition for the development of a safe cosmetic agent or topical products for skin regeneration. The development of SRGF may also provide an alternative strategy for tissue engineering.
Introduction
Cutaneous wound healing is a complex process involving re-epithelialization, granulation tissue formation, and tissue remodeling in all skin components, including epidermis, dermis, hypodermis, blood vessels, and connective tissue, leading to wound closure and repair of tissues. GFs and cytokines are key players in the regulation of these events in which they mediate the coordinated action of keratinocytes, fibroblasts, endothelial cells, and inflammatory cells, which eventually promote wound healing and tissue repairing. The application of exogenous GFs to a wound has been shown to increase the rate of wound closure, the number of cells in the healing area, the growth of blood vessels, the deposition rate of collagen, and the strength of the scar. 10 Several growth factors and cytokines, including TGF-β, PDGF, keratinocyte growth factor (KGF), and insulin, have demonstrated beneficial effects on the wound healing process. 11 –13 However, evaluation of recombinant GFs in clinical use is still underway. 14
The use of PRP in wound healing represents the application of multiple GFs in biologically more similar ratios than a single GF or a combination of GFs. PRPs of autologous or homologous origins have been widely applied in studies. 7 Although the preparation process is relatively simple, many confounding variables, including differences in PRP characteristics and GF contents, can lead to inconsistent outcomes of application and even draw criticisms regarding the efficacy of PRP. 15
Autologous PRP is considered safer than allogeneic or homologous preparations because it is more likely to be free from concerns for transmissible diseases. 3,15 –17 However, the potential use of autologous RPR in acute phase, greater surface area, or general cosmetic applications is limited by the feasibility of large-scale production. The provision of a safe and effective PRP preparation produced by an economical and easy process would be highly favored.
Therefore, the aim of this study was to evaluate an activated PRP preparation, namely skin renewal growth factors (SRGF), developed from porcine whole blood due to its availability and its characteristics feasible for quality control. Furthermore, the effectiveness of SRGF was determined by cultured human fibroblasts, keratinocytes, and melanocytes exposed to SRGF containing medium with various concentrations of TGF-β1 as a quantitative indicator.
Materials and Methods
Preparation of SRGF
Porcine whole blood was collected and subjected into a MCS+ plasma collection system (Haemonetics Corp., Braintree, MA) to isolate the platelets. For platelet activation, bovine thrombin (Sigma) and CaCl2 solution were added into the plasma. The activated plasma was shaken at 100 rpm for 30 min, and the SRGF solution was then collected by centrifugation at 3500 rpm for 10 min. As PRP is enriched with TGF-β1, the level of TGF-β1 in SRGF was determined using enzyme-linked immunosorbent assay (ELISA; Quantikine ELISA kit R&D, Germany) and used as an indicator to quantify the growth factors. The SRGF solution was then diluted to various concentrations (according to TGF-β1=100, 200, 500 pg/mL, and 1 ng/mL) with basal medium containing 1% fetal bovine serum (FBS). Both basal medium and SRGF-containing medium used for cell growth were changed every 2 days for the following assays.
Cell cultures
Adult human dermal fibroblasts (hDFa; Cascade Biologics, Portland, OR) were grown at a density of 5×105/mL in Dulbecco modified Eagle medium (DMEM) supplemented with 10% FBS (GIBCO) and 1% pencillin-streptomycin, and then placed in an incubator with humidified atmosphere at 37°C, 5% CO2. Human dermal keratinocytes (HEKa; Cascade Biologics, Portland, OR) and human epidermal melanocytes (HEMa-LP; Cascade Biologics, Portland, OR) were maintained in M154 basal medium (Cascade Biologics, Portland, OR) and medium 254 containing human melanocyte growth supplement-2 (HMGS-2; Cascade Biologics, Portland, OR) and antibiotic/antimycotic solution for routine cultures, respectively.
Assessment of cell viability
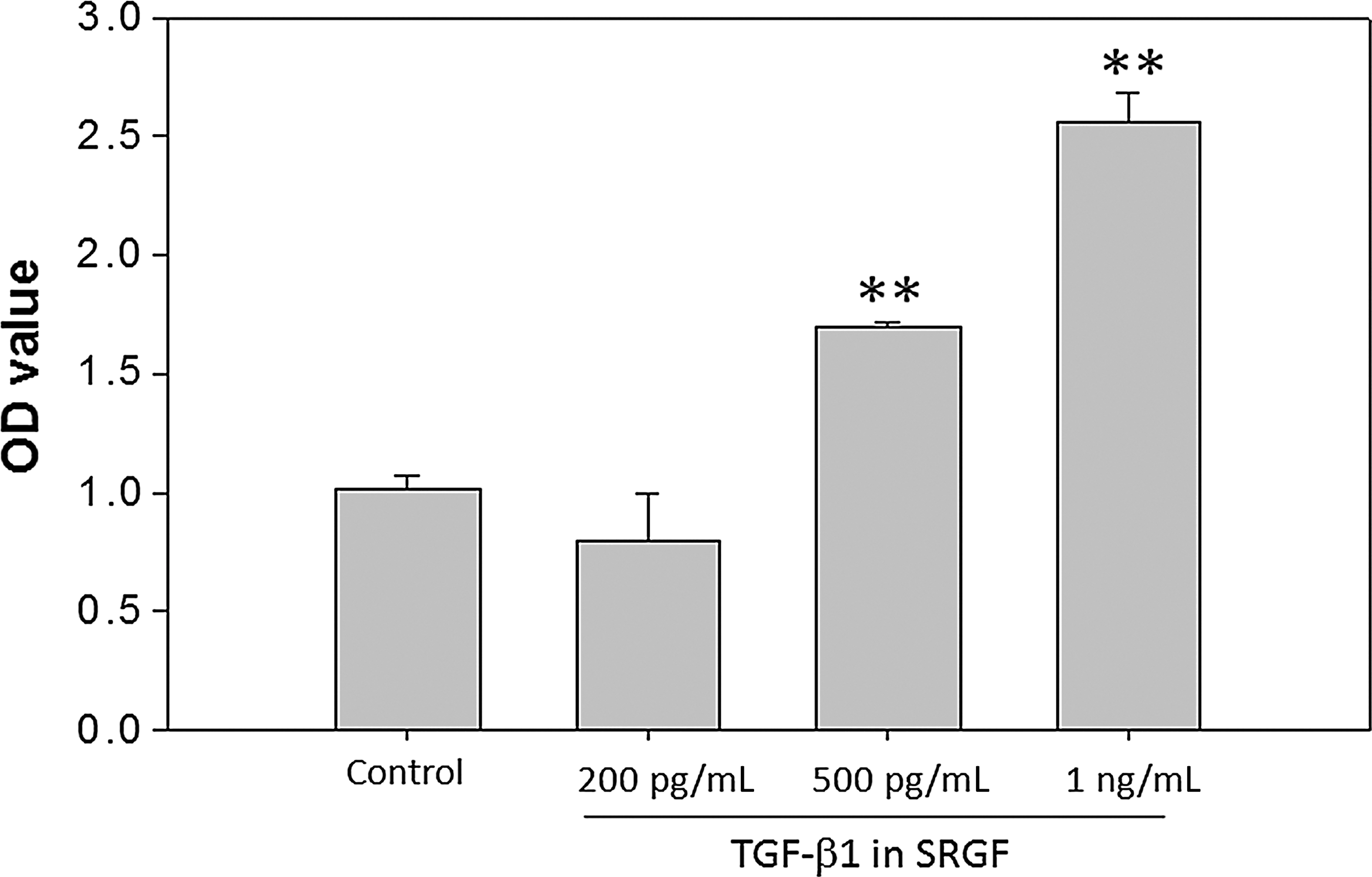
Cell viability was judged by changes in permeability states. The numbers of viable cells were counted on a hemocytometer with the use of a phase-contrast microscope after staining with 0.1% Trypan Blue. To determine the concentration of SRGF that provided optimal performance for following assay, quantitative measurements of cell viability of were performed using a 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT; Roche, Germany) colorimetric assay. Cells were seeded into 96-well plates at a density of 4×103 cells/200 μL per well, in the presence or absence of SRGF (the concentrations of TGF-β1 ranged from 100 pg/mL to 1 ng/mL). Cell proliferation was determined by measuring optical densities (OD) 4 hr after the MTT reaction using a Multiskan PC (Thermo Labsystem, Finland). The quantity of formazan product was directly proportional to the number of living cells, as measured by the absorbance at 570 nm. Cell survival curves were then plotted against incubation time.
Reverse-transcription polymerase chain reaction
Total RNA was extracted from harvested sub-confluent monolayer cultures using TRIzol® reagent (Invitrogen Life Technologies, Carlsbad, CA) and subjected to reverse transcription polymerase chain reaction (RT-PCR) for examining specific gene expression. Specifically, cDNA was synthesized by reverse transcription using SuperScript™ III reverse transcriptase (Invitrogen Life Technologies) and an oligo d(T) 12 –18 primer. Four micrograms of RNA was added into a final volume of a 21-μL reaction mixture containing 10 mM deoxyribonucleoside triphosphate (dNTP) mix, 10×RT buffer, 25 mM MgCl2, 0.1 M dithiothreitol (DTT), 1 μL of RNase inhibitor, and 1 μL of RNase H. For PCR amplification, 6 μg of cDNA was added to a 50-μL reaction mixture containing 2.5 mM dNTP, 25 mM MgCl2, upstream/downstream primers (see Table 1), and Taq DNA polymerase (Invitrogen-Life Technologies). The DNA amplification was performed in a Touchgene Gradient Thermal Cycler (Techne, Cambridge, UK) with a cycle of 5 min at 95°C for initial denaturation, followed by 35 cycles of denaturation at 94°C for 1 min and extension at 72°C for 1 min, and a final extension at 72°C for 5 min. The annealing temperatures were optimized for different genes and primers. PCR products were then separated on 1% agarose gels (Agarose I, AMRESCO, OH) and detected with ethidium bromide staining. Images were analyzed using FloGel-I (Fluorescent Gel Image System, TOP BIO Co., Taiwan). Quantification was always normalized using glyceraldehyde 3-phosphate dehydrogenase (GAPDH) as an internal control.
RT-PCR, Reverse transcription polymerase chain reaction; TGF-β1, transroming growth factor-β1; MITF, microphthalmia-associated transcription factor; PAX3, paired box 3; GAPDH, glyceraldehyde 3-phosphate dehydrogenase; Col, collagen; F, forward; R, reverse.
Alcian Blue staining for proteoglycan accumulation
Human fibroblasts were cultured in the basal or SRGF-containing medium. After 7-day cultures, confluent cells on 96-well plates were fixed in 3.7% formaldehyde and stained with 0.05% Alcian Blue in 0.018 M acetic acid containing guanidine-HCl and Triton X-100 for 3 hr. The fixed cells were then washed in 0.018 M acetic acid, followed by gradual elution of the bound dye by dissociated solution. To quantify the amount of dye incorporated, the eluted solutions were examined using a spectrophotometer at 675 nm (Thermo Labsystem, Finland) with the use of dissociated solution as blank.
Quantification of melanin synthesis
The quantification of synthesized melanin was performed in triplicate and repeated at least twice. After the HEMa-LP cells were cultured in the basal or SRGF-containing medium in 96-well plates for 7 days, cells were washed with phosphate-buffered saline (PBS). The wells were decanted by inversion and emptied by tapping the inverted plates on paper towels. The cells were then lysed with 1 mL of 1 N NaOH and pipetted repeatedly. Two hundred milliliters of each crude cell extract was transferred into 96-well plates for quantification assay. Relative melanin content was determined by optical density at 405 nm using a spectrophotometer (Thermo Labsystem, Finland).
Statistical analysis
Statistical analysis was performed with the SPSS ver. 12.0 (SPSS Inc., Chicago, IL) for Windows. Results were determined from a minimum of three independent experimental data with triplicate measurements per experiment. Data are expressed as mean±standard error of the mean (SEM). Student t-test tests were used for comparison between groups. A probability of less than 0.05 was considered to be statistically significant.
Results
SRGF promoted the proliferation of cultured human fibroblasts
The progression of cellular proliferation of cultured human fibroblasts, hDFa, was determined by treatment with 100, 200, 500, 750 pg and 1 ng per mL TGF-β1 of SRGF-containing medium for 7 days. Compared to the control group, the cells treated with SRGF initiated to proliferate within 3 days and increased remarkably in numbers by 5 and 7 days (Fig. 1). Fibroblast proliferation induced by SRGF was shown in time- and dose-dependent manners according to the indicated concentrations of TGF-β1. On the day 7, cells exposed to higher concentrations of SRGF exhibited significantly higher rates of proliferation than control cells that were grown in medium supplemented with 1% FBS. The increases of fibroblast numbers were correlated to the concentrations of SRGF (Fig. 2A); the cells were fully confluent and almost 5 times more cells after treatment with SRGF containing 1 ng/mL of TGF-β1 (Fig. 2B). These results indicate that SRGF promoted the proliferation of fibroblasts, with 1 ng/mL of TGF-β1 producing maximal stimulation of proliferation. No sign of cell toxicity, such as rounding or loss of adherence, was observed for up to 7 days.

Proliferative effect of skin renewal growth factors (SRGF)-containing medium with indicated concentrations of transforming growth factor-β1 (TGF-β1) (100, 200, 500, 750 pg/mL and 1 ng/mL) on the human dermal fibroblasts (hDFa) was determined by 3-(4, 5-dimethylthiazol-2-yl)-2, 5-diphenyl tetrazolium bromide (MTT) assay after 1, 3, 5, and 7 days of incubation time and compared to untreated control (CTRL) cells. The data are represented as a mean±standard error of the mean (SEM) of three independent replicates. OD, Optical density.

Skin renewal growth factors (SRGF)-induced proliferation on the human dermal fibroblasts (hDFa). (
SRGF induced collagen expression and proteoglycan synthesis of human fibroblasts
To test the potential function of SRGF in regulating collagen production in cultured hDFa, the collagen gene expression was analyzed and quantified by RT-PCR after 7 days of incubation. As shown in Fig. 3, when the expression levels were normalized to GAPDH as a housekeeping gene in fibroblasts, the mRNA expression patterns of collagen subtypes I, III, IV, and VII were all significantly increased by 7 days of SRGF (1 ng/mL of TGF-β1) induction (p<0.05; Fig. 3). The results indicated that the relative collagen synthesis of human fibroblasts was accelerated, nearly doubled in comparison to the control group, by the addition of SRGF in culture medium. As shown in Fig. 4, proteoglycan synthesis in fibroblasts was substantially increased in the presence of SRGF with 500 ng/mL and 1.0 ng/mL of TGF-β1 than control at 7 days as evidenced by Alcian Blue staining, although cells treated with lower concentrations of SRGF (TGF-β1=200 ng/mL) displayed a slight decrease.
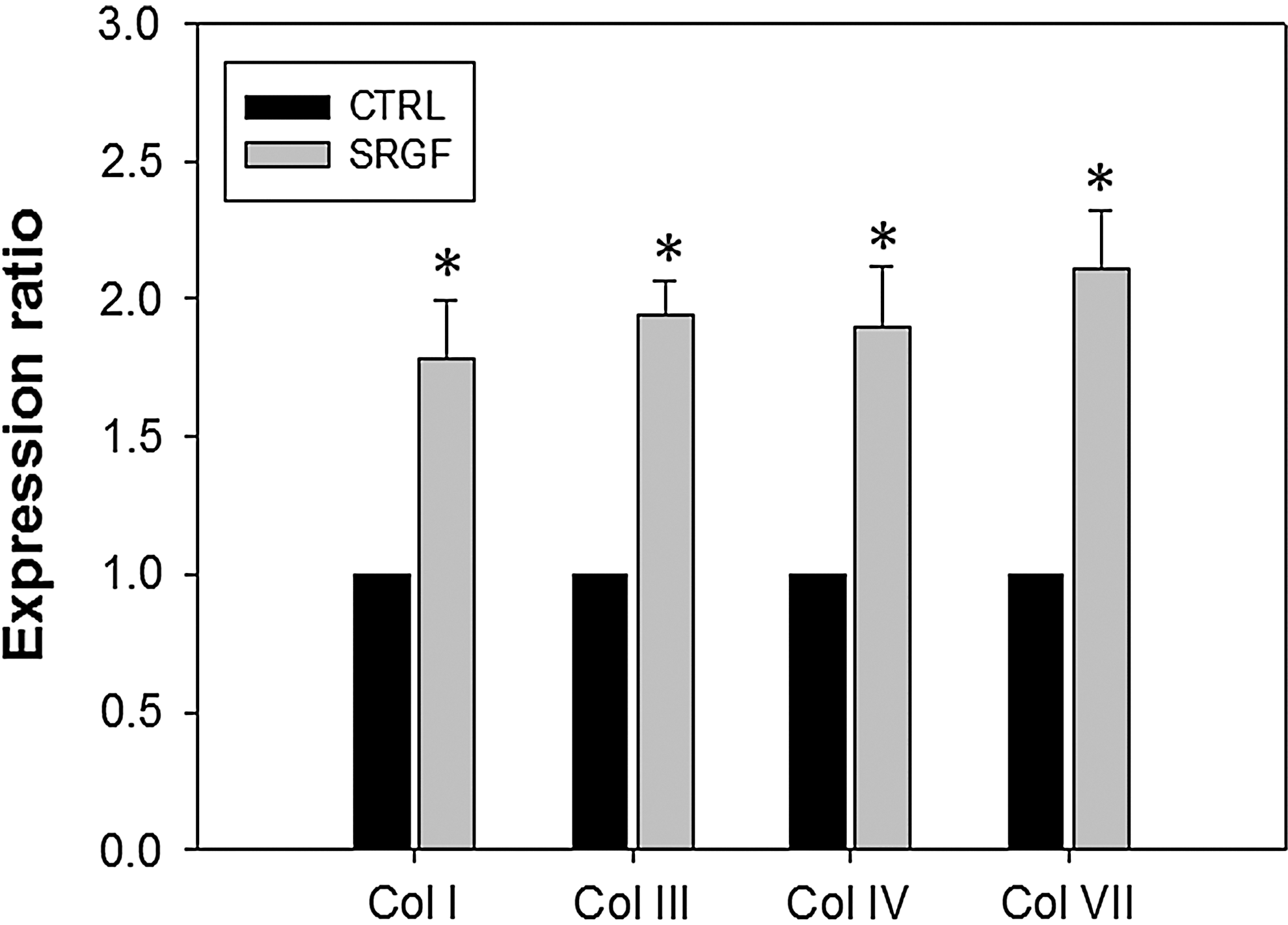
Skin renewal growth factors (SRGF) up-regulates the mRNA levels of collagens in human dermal fibroblasts (hDFa). Relative quantities of collagen mRNA were expressed as mean±standard error of the mean (SEM) of the ratio of the human fibroblasts (hFB) treated with SRGF (normalized to glyceraldehyde 3-phosphate dehydrogenase [GAPDH]/untreated control (CTRL) (normalized to GAPDH). Reverse transcription polymerase chain reaction (RT-PCR) was used to evaluate the differences in gene expression between untreated control and collagen types. Expression levels of collagen (Col) subtypes I, III, IV, and VII were found significantly higher for hFB-treated with SRGF than that untreated (*p<0.05).
Proteoglycan synthesis in human dermal fibroblasts (hDFa) is promoted by skin renewal growth factors (SRGF)-containing medium in the presence of transforming growth factor-β1 (TGF-β1) (200, 500 pg/mL and 1 ng/mL) for 7 days. Proteoglycan synthesis was measured by Alcian Blue staining and expressed as optical density (OD) at 630 nm. Results are indicated as mean±standard error of the mean (SEM) of data normalized to total cell numbers from three independent replicates. (**) p<0.01 versus untreated control.
SRGF inhibited proliferation and differentiation of human dermal keratinocytes
To understand the effects potentially exerted by SRGF in skin regeneration, we investigated the response of keratinocytes grown in the SRGF-containing medium. Total RNA was subsequently extracted and used for RT-PCR. The results demonstrated that SRGF inhibited the growth of keratinocytes in a dose-dependent manner (Fig. 5A), which was coupled with significantly elevated expression of TGF-β1 in keratinocytes (Fig. 6). The reduced keratinocyte differentiation was accompanied as evidenced by the attenuated expression of laminin V and keratin 10 (Fig. 6). The decreased levels of laminin V indicated that there was reduced migration, cell polarization of inactivated keratinocytes, which was also observed in Fig. 5B.
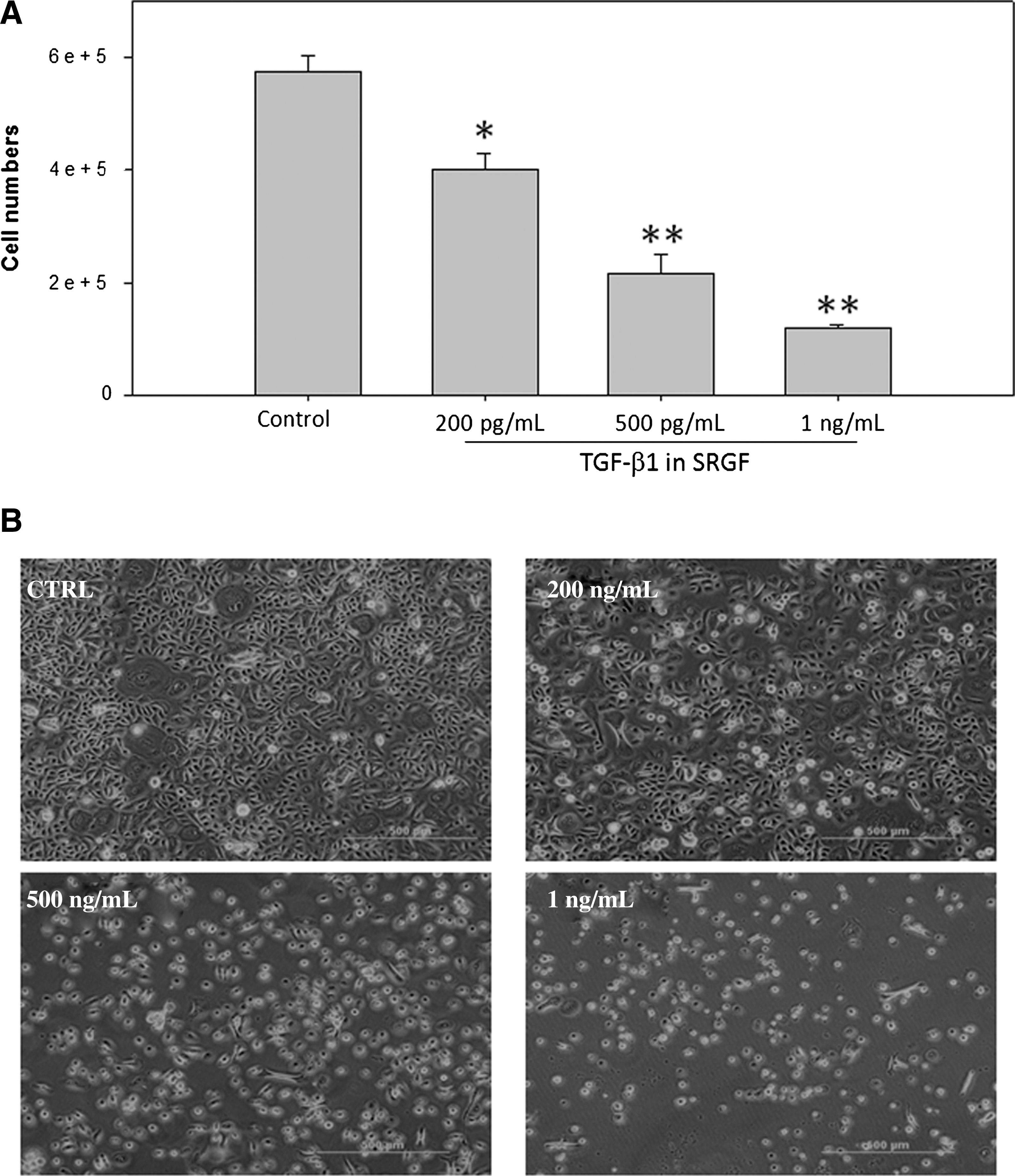
Skin renewal growth factors (SRGF)-induced changes on the human dermal keratinocyte (HEKa) proliferation. (
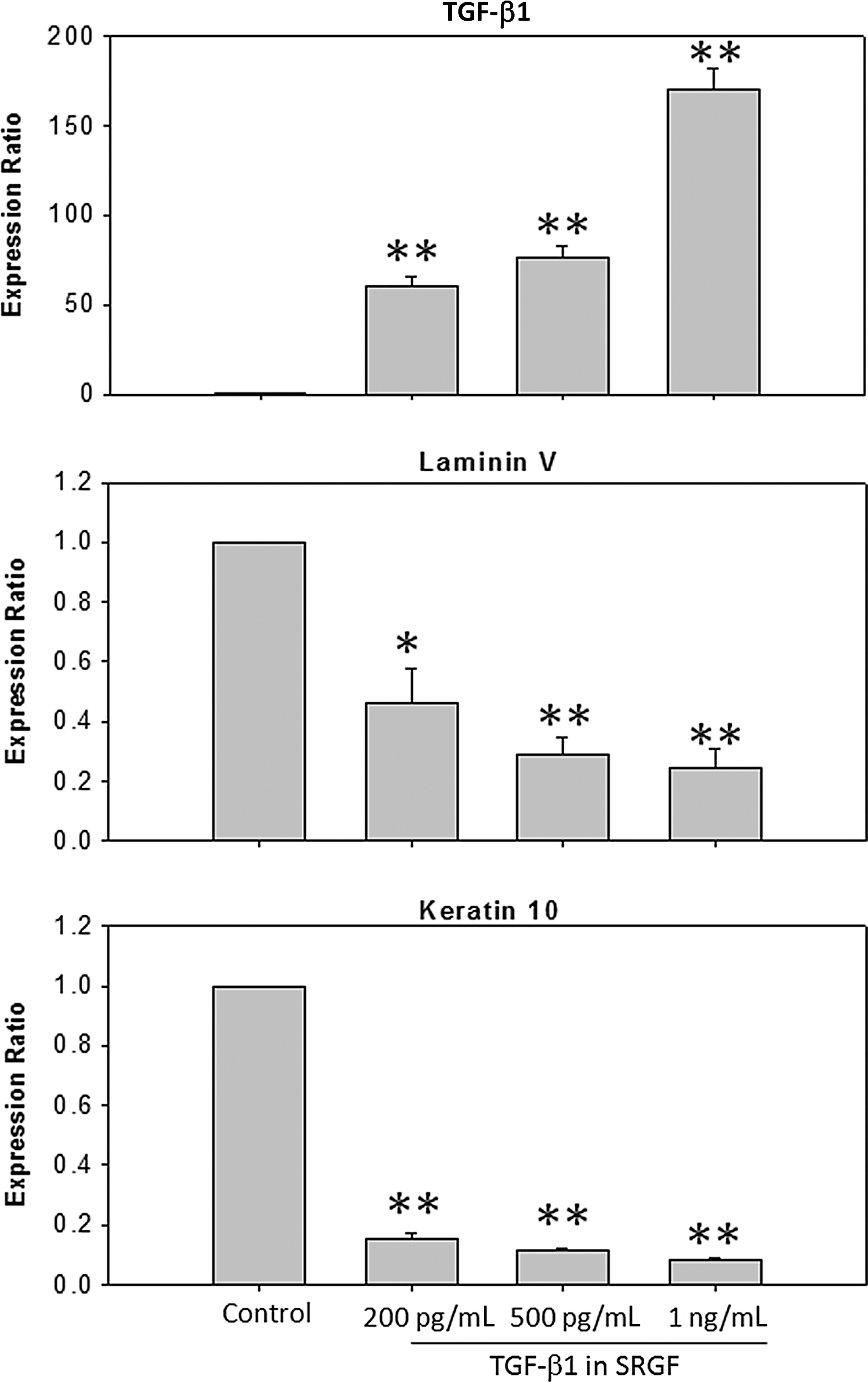
Exposure of cultured human dermal keratinocytes (HEKa) to skin renewal growth factors (SRGF) resulted in altered expression ratios of transforming growth factor-β1 (TGF-β1), laminin V, and keratin 10. Relative quantities of mRNA were expressed as mean±standard error of the mean (SEM) of the ratio of the HEKa cells treated with SRGF/untreated control. The mRNA levels were measured by reverse transcription polymerase chain reaction (RT-PCR) and normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH). TGF-β1 expression is up-regulated by SRGF-containing medium in a concentration-dependent manner, whereas differentiation was inhibited as evidenced by decreased expression of laminin V and keratin 10. (*) p<0.05; (**) p<0.01.
SRGF suppressed the growth and melanogensis of melanocytes
Effects of SRGF on melanocyte growth and melanin production were assessed after 7 days of treatment. Co-culture with higher concentrations of SRGF (TGF-β1=500 ng/mL and 1 ng/mL) significantly inhibited proliferation and decreased dendricity of melanocytes compared with controls (Fig. 7A,B). Although SRGF at a lower concentration (as indicated, TGF-β1=200 ng/mL) conversely increased the growth in cell numbers, reduced dendricity in morphological features and decreased melanin production were consistent trends. The expression levels of PAX3, a key transcriptional factor that promotes proliferation and stimulates melanin synthesis in mature melanocytes, 18,19 were repressed by SRGF in a dose-dependent manner (Fig. 8). At higher concentrations of SRGF, significantly reduced expression of PAX3 mRNA, accompanied by remarkably decreased mRNA levels of microphthalmia-associated transcription factor (MITF) and tyrosinase, was detected (Fig. 8). The decreased expression of MITF elicited by SRGF correlated well with the decreased expression of tyrosinase that is regulated by MITF. 20,21 The inhibitory effect of SRGF on melanogenesis was subsequently studied by measuring melanin synthesis. In the presence of SRGF for 7 days, the melanin content of melanocytes was significantly lowered (Fig. 9), showing 68.1% at 200 pg/mL, 60.6% at 500 pg/mL, and 57.6% at 1 ng/mL of indicated TGF-β1, compared with the control group. Together, these results suggest that SRGF inhibited melanocyte proliferation and suppressed the expression of MITF, tyrosinase, and PAX3, which mediates melanogensis.
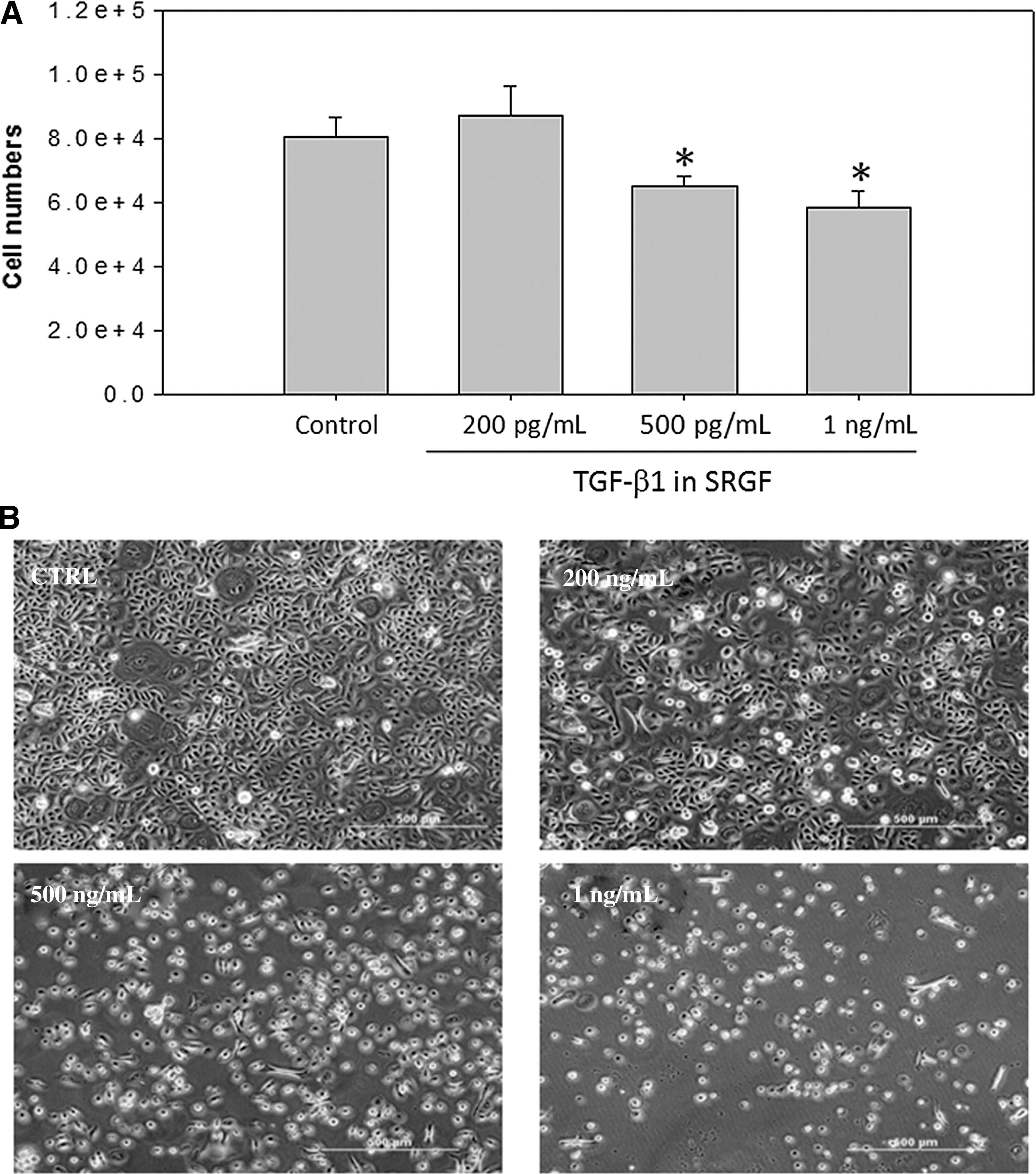
Skin renewal growth factors (SRGF)-induced changes on the human epidermal melanocytes (HEMa-LP) proliferation. (
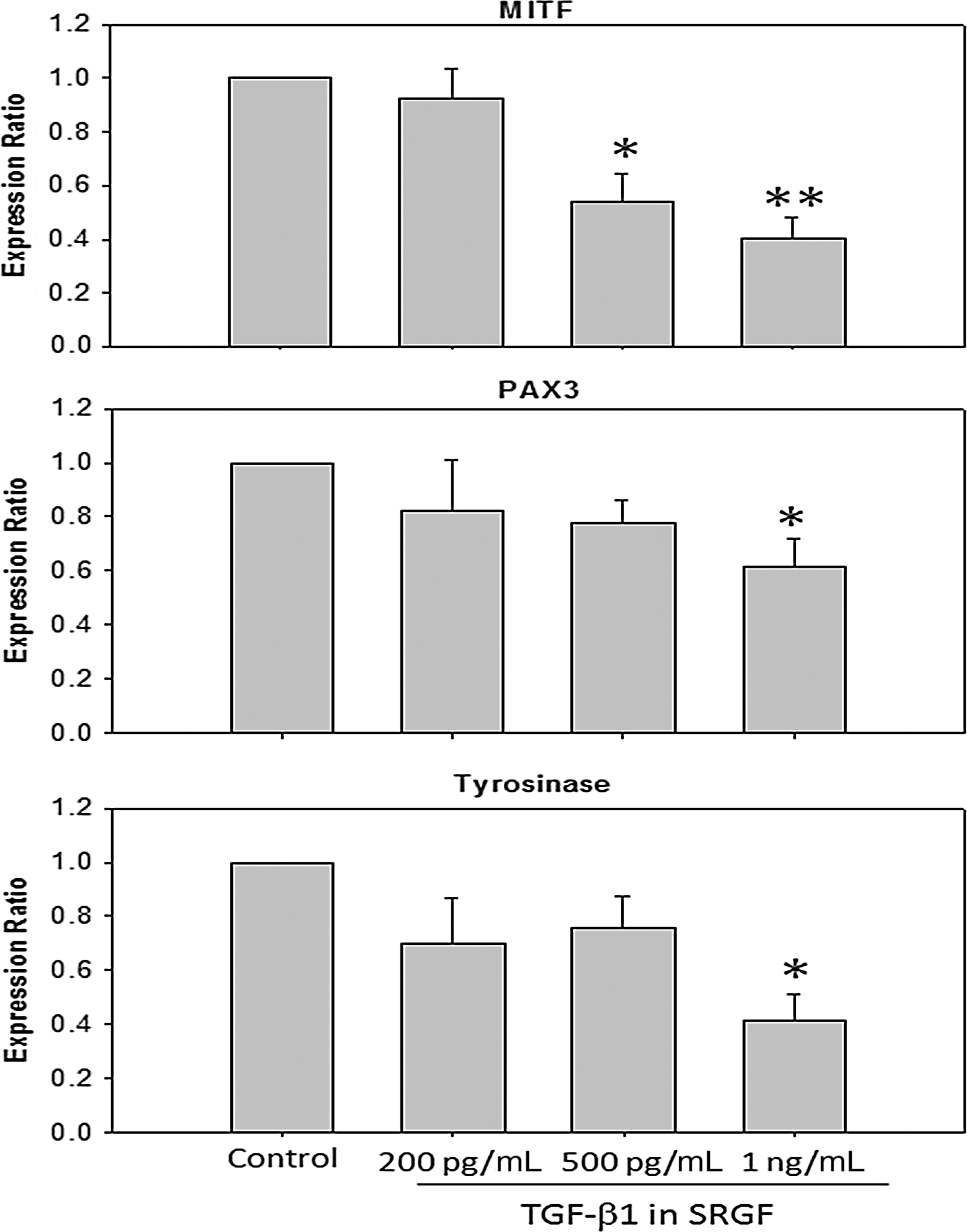
Reverse transcription polymerase chain reaction (RT-PCR) analysis of the expression of mRNA for microphthalmia-associated transcription factor (MITF), paired box 3 (PAX3), and tyrosinase in the human epidermal melanocytes (HEMa-LP). Relative quantities of mRNA were expressed as mean±standard error of the mean (SEM) of the ratio of the HEMa-LP cells treated with skin renewal growth factors (SRGF)/untreated control. MITF, PAX3, and tyrosinase are significantly down-regulated by SRGF-containing medium in concentration-dependent manners. The mRNA levels were normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH). (*) p<0.05; (**) p<0.01.

Skin renewal growth factors (SRGF)-containing medium showed melanin production inhibitory effect on melanogensis in human epidermal melanocytes (HEMa-LP). Melanin content was detected by melanin assay. These data are representative results of three independent experiments. Data are reported as mean±standard error of the mean (SEM). (*) p<0.05; (**) p<0.01. OD, Optical density.
Discussion
Tissue repair is a complex process. Strategies to improve regeneration involving delivering the optimal combination of GFs, targeting the appropriate cells that contribute to the maintenance of structural integrity, and re-establishment of matrix scaffold pose significant challenges for tissue engineering. 22 Although significant progress in this area has been made, the hurdles of consistent manufacturing, preservation, and shipping in compliance with regulations remain challenging for further clinical and commercial advancements. Human PRP has been shown to induce wound healing and skin repair, thus indicating that a porcine platelet-derived wound healing formula may constitute an advantageous alternative to a wider range of applications than the use of recombinant GFs and human PRP if proven to be economically available and practically effective. 4,5,7,8 The wound healing formula is termed SRGF, and its mechanisms for dermal regeneration were examined in this study.
PRP is widely applied in many clinical fields and has attracted attention of dermatologists for its various growth factors (including TGF-β1, VEGF, PDGF, bFGF, etc.) released by the α-granules. TGF-β1 is found in many different cell types, but its highest concentration is in platelets. 23,24 Normal function of TGF-β1 represents a multifunctional regulator of cell proliferation, differentiation, migration, and survival that affect multiple biological processes, including development, carcinogenesis, fibrosis, wound healing, and immune responses. 25 –29 In addition, TGF-β1 markedly inhibits the growth of keratinocytes at concentrations greater than 2 ng/mL under conditions of low Ca2+. 30 Cultured melanocytes are highly sensitive to the inhibitory effect of all TGF-β isoforms. 31 Furthermore, TGF-β1 suppresses the transcription of PAX3, which down-regulates the expression of MITF in melanocytes. 19 Therefore, TGF-β1 is considered a key cytokine in wound healing and hypertrophic scarring. Clinical applications of PRP with moderate platelet enrichment are efficacious in healing. 32,33 Because PRP is enriched with TGF-β1 and the optimal concentration of porcine PRP was unclear, we used it as the quantitative indicator to optimize the preparation and to evaluate the effectiveness of SRGF in the current study.
VEGF is involved in the regulation of angiogenesis during wound healing. 13,34 PDGF stimulates the proliferation of fibroblasts and smooth muscle cells and may participate in wound healing by stimulating dermal cells. 20 bFGF stimulates keratinocyte migration 21 and wound healing. 35 EGF family members activate an intracellular signaling cascade through their receptors to affect keratinocyte growth and migration and subsequent healing of skin wounds. 36 IGF-I also has a vital role in wound healing through its action on fibroblasts, 37,38 aside from stimulating fibroblast mitogenesis and ECM synthesis. 39 According to the reports above, these factors that are also in SRGF are commonly considered necessary for the initiation, propagation, and maintenance of wound healing, and are potentially useful for reducing skin wrinkles and rejuvenating the skin in addition to many clinical applications. Deregulation of these functions is associated with excessive connective tissue deposition including fibrosis or scarring in the skin. 40
Fibroblasts are the most common cells in connective tissue and they have a prominent role during wound repair. They proliferate and invade lesions, modulate local inflammation, generate extracellular matrix (ECM) to serve as a scaffold for other cells, and possess cytoskeletal elements that facilitate contractions of healing wounds. 41 Conventional anti-aging strategies, such as light, lasers, and topical treatments, typically aim to increase ECM synthesis through the activation of fibroblasts. Fibroblasts also produce many GFs and cytokines that act on other cell types, thereby modulating tissue repair and scarring. 42,43 Disorder of fibroblast proliferation would result in delayed wound healing or tissue fibrosis. 44,45
Under normal and non-wound conditions, the ECM consists primarily of collagens, the most abundant proteins in the body. Collagens are comprised of a group of diverse protein subfamilies containing at least 28 different collagen types that are encoded by more than 42 genes. 42,46 Collagens in the skin are primarily type I and type III, which provide structure, strength, and integrity. Type IV collagen is a component of epidermal and endothelial basement membranes, and collagen VII forms the anchoring fibrils in the skin. 47 Synthesis, deposition, and remodeling of ECM proteins are essential for the restoration of damaged tissue during the early and late phases of wound healing. Proteoglycans also serve as co-receptors for growth factors, participate in cell signaling, and support to regulate the activity of many other molecules. These activities enable glycosaminoglycans and proteoglycans to play key roles in cell adhesion and migration. To evaluate the effects exerted by SRGF, mRNA expression of collagens and proteoglycan synthesis were determined in this study. As shown in Figs. 3 and 4, SRGF may induce ECM remodeling by facilitating expression of both collagens and proteoglycans. This ability of SRGF to increase collagen could be beneficial and result in faster wound healing. In accordance with these results, the production and deposition of normal structural and functional collagen to make normal physiological connective tissue can be promoted by SRGF.
Numerous factors produced in and released from keratinocytes may be involved in regulating the proliferation and differentiation of mammalian epidermal melanocytes. 48 –50 A proper balance of proliferation and differentiation of basal cell population is substantial for maintaining tissue integrity for epidermal hemostasis. It is known that the balance between the continuous proliferation of basal keratinocytes, maturation, and cell death of terminally differentiation of keratinocytes is regulated by various factors that are involved in the “switch” from proliferation to differentiation. 54 According to previous studies, 52 retinoids induce proliferation in keratinocytes and consequently result in epidermal hyperplasia and modulate epidermal differentiation. In contrast, vitamin D3 and its analogs inhibited keratinocyte proliferation, which explains why these compounds are effective in treating hyperproliferative skin disorders, such as psoriasis. 53
The present study revealed that the growth and differentiation of cultured keratinocytes were considerably affected by SRGF-containing medium. In addition to TGF-β1 detected in SRGF medium, the growth inhibition was facilitated by up-regulated TGF-β1, which acted in an autocrine manner to further decrease the growth rate of keratinocytes. 54 Reduced proliferation and morphological alterations of keratinocytes (Fig. 6) observed in our study were the processes associated with the arrest of keratinocytes. Keratins are used as markers due to their specific localization in the skin layers during the process of differentiation. In normal skin, keratin 10 is considered to be an early differentiation marker that is expressed in maturing keratinocytes detached from the basal laminin. 55,56 Furthermore, the reduced expression of keratin 10 in line with elevated levels of TGF-β1 (Fig. 7) indicates that keratinocyte is restored to the basal phenotype. 57 As the result, reduced hyperplasia and early differentiation of keratinocytes would be a consequence of SRGF application. These findings indicate that SRGF exhibit a vital role in regulating skin regeneration.
Melanocytes are highly differentiated neural crest–derived cells responsible for synthesizing melanin in responding to environmental stimuli. 58 The melanin is transferred in melanosomes through dendritic processes to surrounding epidermal keratinocytes. Many GFs are thought to be involved in the regulation of the proliferation and/or differentiation of melanocytes. 59 MITF is the key transcription regulator of signaling that controls the proliferation and dendricity of melanocytes 60 and promotes melanin synthesis by the up-regulation of tyrosinase. 61 –63 The decreased expression of MITF elicited by SRGF correlates well with the decreased expression of tyrosinase (Fig. 8), suggesting that the activities of SRGF are associated with the down-regulation of MITF. 63,64 In addition, the properties of SRGF inhibiting melanocyte growth in concert with the diminished expression levels of PAX3 make SRGF an excellent candidate for the suppression of melanogenesis.
In conclusion, evidence is presented that the SRGF derived from porcine platelets shows potent and cell type–dependent proliferative and differentiation activities toward human fibroblasts, keratinocytes, and melanocytes, while having no observed toxic activity. The present study also demonstrated that SRGF possess beneficial properties and are a promising and cost-effective component for the development of a safe cosmetic agent or topical products for skin regeneration or used as adjuvant therapy with the standard treatment for wounds. SRGF may also provide an alternative strategy for tissue engineering.
Footnotes
Acknowledgments
The authors thank to SRGF Biotech Ltd. Co. for the provision of SRGF as a free gift. The auuthors also sincerely thank all volunteers, technical personnel, and colleagues.
Author Disclosure Statement
No competing financial interests exist.