
Abstract
Human growth hormone-2 (GH-2) is a 191-amino-acid protein also known as human placental hormone. During pregnancy, continuous secretion of GH-2 appears to have important implications for physiological adjustment to gestation, especially in controlling levels of maternal insulin-like growth factor 1. To compare the physiological activity of GH-2 between lipid-free and lipid-bound states, GH-2 was expressed and incorporated into proteoliposome. GH-2 was expressed and purified using a pET28(a)-GH-2 vector in an Escherichia coli system. Purified GH-2 was then characterized and synthesized into reconstituted high-density lipoprotein (rHDL). The expression yield of GH-2 was 20–30 mg by BL21 (DE3) cells in 1 liter of Luria-Bertani broth. Purified GH-2 of at least 98% purity (23 kDa) was incorporated into rHDL with human apolipoprotein A-I (ApoA-I) and palmitoyloleoyl phosphatidylcholine (POPC) at a 1:1:95 (GH-2:ApoA-I:POPC) molar ratio. Structural analysis revealed that GH-2 had a 44% α-helix content and a wavelength maximum fluorescence (WMF) of 349 nm in a lipid-free state. In a lipid-bound state, the WMF of GH-2 was ∼4 nm blue-shifted (345 nm), with 50% of α-helix content. The lipid-bound GH-2 showed enhanced anti-atherosclerotic activity and anti-senescence activity with inhibition of fructose-mediated glycation. A fin regeneration experiment using zebrafish (17 weeks old, n=9) showed that lipid-bound GH-2 enhanced regeneration efficiency by 44% compared to native GH-2 (in the lipid-free state) without any notable side effects. GH-2 has anti-oxidant activity to enhance tissue regeneration as well as to exert anti-diabetic activity. Incorporation of GH-2 into rHDL can enhance structural stability and tissue regeneration efficiency in vertebrate models, indicating a synergetic effect between GH-2 and ApoA-I in rHDL.
Introduction
H
Although the physiological role of GH-2 is still under-investigated, the main physiological functions of GH-2 are known to include somatotropic activation for fetal development during pregnancy. 5 Interestingly, GH-2 is found only in the maternal circulation and disappears immediately after parturition, 6 although it can stimulate more fetal growth. 7 Regulation of GH-2 secretion is more complicated and different from that of GH-1. Hyperglycemic induction in pregnant women with diabetes has been associated with the disappearance of GH and human placental lactogen. 8 Although secretion of GH-2 is maintained at high levels during pregnancy, its functions and detailed mechanism of action are not known. GH-2 is found in the maternal circulation from week 6 of gestation 9 and gradually increases to peak values from weeks 34 to week 37. During pregnancy, the protein level of placental GH (GH-2) increases up to 20–30 ng/mL in the maternal serum. 10 Although the main function of GH-2 is promotion of fetal growth, GH-2 might exert other beneficial functions for maternal adaptation and protection. It is plausible that GH-2 might provide protective effects against metabolic stress, such as oxidation and glycation in the maternal blood. In addition to increasing growth in the fetus and in children, GH-2 is important for tissue regeneration in adults, maintenance of bone and muscle homeostasis, and wound healing. Reduced GH-2 levels have been found in pregnant women with the smallest fetuses. 9 However, the functions of GH-2 during pregnancy for maternal health have not yet been reported, especially regarding lipoprotein metabolism.
It has been reported that GH-2 is increased in the maternal and fetal sera of patients with pre-eclampsia, 10 and its physiological role is supposedly related to trophoblast invasion, fetal growth, and maternal adaptation to pregnancy. 11 Especially in patients with diabetes mellitus (DM), tissue regeneration activity was impaired in an adult zebrafish model, indicating tissue regeneration activity, was deprived by elevated advanced glycation end products (AGE).
Many reports have suggested that both low-density lipoprotein cholesterol (LDL-C) and high-density lipoprotein cholesterol (HDL-C) are delivered into the placenta from the maternal circulation in mice 12 and humans, 13 whereas GH-2 is found only in the maternal circulation. In a fasting state, HDL is a major carrier of cholesterol in reverse cholesterol transport for its excretion. HDL possesses potent anti-oxidant activity in the serum whereas apolipoprotein A-I (ApoA-I), a principal protein of HDL, suppresses atherosclerosis and cardiovascular disease. Due to the anti-oxidant and anti-glycation activities of ApoA-I, serum levels of ApoA-I and HDL are important for maintenance of health status. 14
Glucose inhibits human GH-2 secretion in vitro, 15 and pregnant subjects with type 1 DM show reduced maternal GH-2 levels along with increased fetal GH-2 levels. 16 Insulin resistance has been observed in transgenic mice expressing human GH-2 as well as a two-fold increase in body weight by the third trimester of pregnancy. 17 These studies imply that GH-2 plays physiological roles in lipid and glucose metabolism for fetal development during pregnancy. However, there has been no report investigating the additional functions of GH-2 in the maternal serum.
Therefore, the functional roles of GH-2 were examined in this study in lipid-free and lipid-bound states using human cells and zebrafish. In the current study, we expressed and purified human GH-2 using an Escherichia coli expression system and characterized its physiological functions in the lipid-free and lipid-bound state.
Materials and Methods
Materials
Palmitoyloleoyl phosphatidylcholine (POPC; cat. no. 850457) was obtained from Avanti Polar Lipids (Alabaster, AL). Restriction enzymes were obtained from New England BioLabs (Beverly, MA). The pET28a(+) expression vector as well as E. coli BL21 (DE3) were purchased from Novagen (Madison, WI).
Expression and purification of GH-2
The human GH-2 gene was cloned from pDNR-LIB (cat. no. hMU013780, Human Gene Bank of Korea, Daejeon, Korea) using PCR with the designed primer pair (forward, 5′-ATGGTACCATGGATATGTTCCCAACCATTCCCT TA-3′; reverse, 5′-ATGGTAATACTCGAGGAAGCCACAGCTGC-3′) to generate XhoI and NcoI sites for construction of the expression vector. The sub-cloned cDNA was inserted into pET28a expression vector and verified by DNA sequencing using a Sequentator (ABI7500, ABI, Foster City, CA). The expressed polypeptide in mature form was 191 amino acids long, and a His tag (eight amino acids, L-E-HHHHHHH) was attached to the carboxyl terminal. The His-tagged GH-2 gene was expressed and purified using a Ni2+-nitrilotriacetic acid column chromatography (Peptron, cat. no. 1103-3, Daejeon, Korea), as described in our previous report. 18,19
Protein sequencing
Protein samples for sequencing were electrotransferred onto a polyvinylidene fluoride, (PVDF) membrane (Immobilon-P) according to the standard protocol. The amino-terminal amino acid sequence of the excised band was determined using an Applied Biosystems model 491A sequencer (Foster City, CA) located in the Korea Basic Research Institute (Daejeon, Korea).
Circular dichroism and fluorospectroscopy
Average α-helix contents of the proteins in lipid-free and lipid-bound states were measured by circular dichroism spectroscopy using a J-700 Spectropolarimeter (Jasco, Tokyo, Japan) located in the Korea Research Institute of Bioscience and Biotechnology (Daejeon, Korea). Spectra were obtained from 250 to 190 nm at 25°C in a 0.1-cm path-length quartz cuvette at a bandwidth of 1.0 nm, speed of 50 nm/min, and response time of 4 sec. Samples of lipid-free proteins were diluted to 0.07 mg/mL to avoid self-association, whereas lipid-bound proteins were diluted to 0.1 mg/mL. Four scans were accumulated and averaged. The α-helical content was calculated from the molar ellipticity at 222 nm using a mean residue weight of 117.3 for GH-2. 20
Characterization of Trp fluorescence and isothermal denaturation
The wavelength of maximum fluorescence (WMF) of Trp residues in the ApoA-I protein was determined from uncorrected spectra obtained using an LS55 spectrofluorometer (Perkin-Elmer, Norwalk, CT) in conjunction with WinLab software package 4.00 (Perkin-Elmer) and a 1-cm path length Suprasil quartz cuvette (Fisher Scientific, Pittsburgh, PA). Briefly, samples were excited at 295 nm to avoid tyrosine fluorescence, and the emission spectra were scanned from 305 to 400 nm at room temperature. The effects of the addition of urea on the secondary structures of GH-2 and ApoA-I in the lipid-bound state were monitored by observing WMF, as in our previous report. 21
To induce denaturation of GH-2 and ApoA-I in lipid-free and lipid-bound states, each protein was treated with urea and incubated with 25°C for 72 hr, because denaturation of the proteins is a very slow process.
Purification and oxidation of LDL
LDL (1.019<d<1.063) was purified from healthy human plasma (Blood Bank of Yeungnam University Medical Center, Daegu, Korea) by ultracentrifugation (100,000×g) for 22 hr at 4°C. Oxidized LDL (oxLDL) was produced by incubating the LDL fraction with CuSO4 (final concentration, 10 μM) for 4 hr at 37°C. oxLDL was then filtered (0.2-μm filter) and analyzed by a thiobarbituric acid reactive substances (TBARS) assay to determine the extent of oxidation, as previously described. 22
Synthesis of reconstituted HDL
Reconstituted HDL (rHDL) was prepared by the sodium cholate dialysis method 23,24 at an initial molar ratio of 95:5:1:1 for POPC:cholesterol:ApoA-I:GH-2, respectively. The size and hydrodynamic diameter of rHDL particles were determined by 8%–25% native polyacrylamide gradient gel electrophoresis (PAGGE, Pharmacia Phast system) by comparison with standard globular proteins (GE Healthcare, Uppsala, Sweden).
Glycation of ApoA-I in lipid-free and lipid-bound states
To compare glycation sensitivity, ApoA-I from each species in a lipid-free or lipid-bound state was incubated with
Western blotting
To compare the cross-reactivity of each ApoA-I protein with antibody specific to the human variant, equal amounts (2.5 μg of protein) of ApoA-I were loaded and electrophoresed on 15% sodium dodecyl sulfate polyacrylamide electrophoresis (SDS-PAGE) gels, followed by detection using GH-2 antibody (AAS13420E; Antibody Verify, Las Vegas, NV), anti-human full-length ApoA-I goat antibody (ab7613; Abcam, Cambridge, UK), and donkey anti-goat immunoglobulin G horseradish peroxidase (HRP) (SC2020, Santa Cruz Biotechnology, Santa Cruz, CA) as a secondary antibody (diluted 1:2000). ApoA-I concentrations in rHDL and lipid-free states were determined by Lowry assay as modified by Markwell et al. 26 with bovine serum albumin as a standard.
Inhibition of LDL oxidation
To determine the extent of oxidation, purified human LDL was incubated with 10 μM CuSO4 for up to 3 hr in the presence of ApoA-I in a lipid-free or rHDL state (final concentration, 2 μM). During incubation, the quantity of conjugated dienes formed was measured by following the absorbance at 234 nm (Abs234) and 37°C 27 using a Beckman DU 800 spectrophotometer (Fullerton, CA) equipped with a MultiTemp III thermocirculator (Amersham, Uppsala, Sweden).
To verify the spectroscopic data, oxidized samples were subjected to electrophoresis on 0.5% agarose gels to compare their electromobilities. 28 Migration of each lipoprotein is known to depend on its intact charge and size. Gels were then dried and the bands stained with 0.125% Coomassie Brilliant Blue.
Acetylation of LDL
Acetylation of LDL (acLDL) was performed using saturated sodium acetate and acetic anhydride, according to a previously described method. 29 After acetylation and subsequent dialysis, acLDL protein content was determined and filtered through a 0.22-μm filter (Millex; Millipore, Bedford, MA). To detect phagocytosis of acLDL, a fluorescent cholesterol derivative (22-[N-7-nitrobenz-2-oxa-1,3-diazol-4-yl]amino-23,24-bisnor-5-cholen-3-ol [NBD-cholesterol], Molecular Probes, cat. no. N-1148; 70 μg of NBD-cholesterol/mg of ApoA-I) was added to the acLDL particles.
LDL phagocytosis assay
THP-1 cells, a human monocytic cell line, were obtained from the American Type Culture Collection (ATCC, cat. no. TIB-202™, Manassas, VA) and maintained in RPMI-1640 medium (Hyclone, Logan, UT) supplemented with 10% fetal bovine serum (FBS) until needed for experimentation. Cells that had undergone no more than 20 passages were incubated in medium containing phorbol 12-myristate 13-acetate (PMA; final concentration, 150 nM) in 24-well plates for 24 hr at 37°C in a humidified incubator (5% CO2, 95% air) to induce differentiation into macrophages. The differentiated and adherent macrophages were then rinsed with warm PBS and incubated with 400 μL of fresh RPMI-1640 medium containing 1% FBS, 50 μL of acLDL (1 mg of protein/mL in phosphate-buffered saline [PBS]), and 50 μL of PBS or each protein (final concentration, 2 μM) for 48 hr at 37°C in a humidified incubator. After incubation, cells were washed with PBS three times and then fixed in 4% paraformaldehyde for 10 min. The fixed cells were stained with Oil Red O staining solution (0.67%) and then washed with distilled water. THP-1 macrophage-derived foam cells were then observed and photographed using a Nikon Eclipse TE2000 microscope (Tokyo, Japan) at 600×magnification.
Cellular senescence assay
Primary human dermal fibroblasts (HDFs) were cultured in Dulbecco's modified Eagle medium (DMEM; Life Technologies; Gaithersburg, MD). HDFs were plated in DMEM at 1×105 cells per 100-mm culture plate and cultured at 37°C in a 5% CO2 humidified incubator, as described in our previous report. 30 HDFs were exposed at passage 11 (approximately 40% confluence) to the indicated concentrations of GH-2 (4, 13, 21, or 42 nM) for 30 days with sub-culture until passage 18. The extent of aging as well as cellular senescence-associated (SA) β-galactosidase (β-Gal) activity were compared by comparing the intensity and area of staining, as previously described. 31
Zebrafish
Zebrafish and embryos were maintained according to standard protocols. 32 Maintenance of zebrafish and all procedures were approved by the Committee of Animal Care and Use of Yeungnam University (Gyeongsan, Korea). Fish were maintained in a system cage at 28°C under a 10:14-hr light cycle with consumption of normal Tetrabit (Tetrabit Gmbh D49304, 47.5% crude protein, 6.5% crude fat, 2.0% crude fiber, 10.5% crude ash, containing vitamin A [29770 IU/kg], vitamin D3 [1860 IU/kg], vitamin E [200 mg/kg], and vitamin C [137 mg/kg]; Melle, Germany).
Fin regeneration
The wound-healing effect of GH-2 was investigated using an streptozotocin (STZ)-induced adult zebrafish diabetes model, similar to our previous report. 30 Experimental zebrafish approximately 12-weeks-old were anesthetized by submersion in 2-phenoxyethanol (Sigma P1126; St. Louis, MO) in system water (1:1000 dilution). Control zebrafish were injected with the same volume of citrate buffer. For fin regeneration studies, STZ-induced zebrafish were anesthetized, and their tail fins were cut with a scalpel close to the proximal branch point of the dermal rays within the fin. After amputation, 10 μL each of protein GH-2 (2 μg of protein) and/or ApoA-I (2 μg) in lipid-free and lipid-bound states was injected into the tail muscle near the urostyle (n=9 for each group). Following injection, fish consumed a normal diet and were observed in a 28°C system incubator. Images of regenerating fins from live zebrafish were taken at 24-hr intervals up to 144 hr under a stereomicroscope (Motic SMZ 168; Hong Kong) and photographed using a Moticam 2300 CCD camera.
Statistical analysis
All data are expressed as the mean±standard deviation (SD) of at least three independent experiments with duplicate samples. Comparisons between results were made by the Student t-test using the SPSS program (v. 12.0; SPSS, Inc., Chicago, IL). Statistical significance was defined as p<0.05.
Results
Purification and characterization of GH-2
Purified GH-2 (23.7 kDa) was revealed to be at least 98% pure by densitometric scan analysis (Fig. 1A), and its sequence was identified by amino-terminal protein sequencing. Western blotting using a GH-2–specific antibody also confirmed the protein (Fig. 1B).
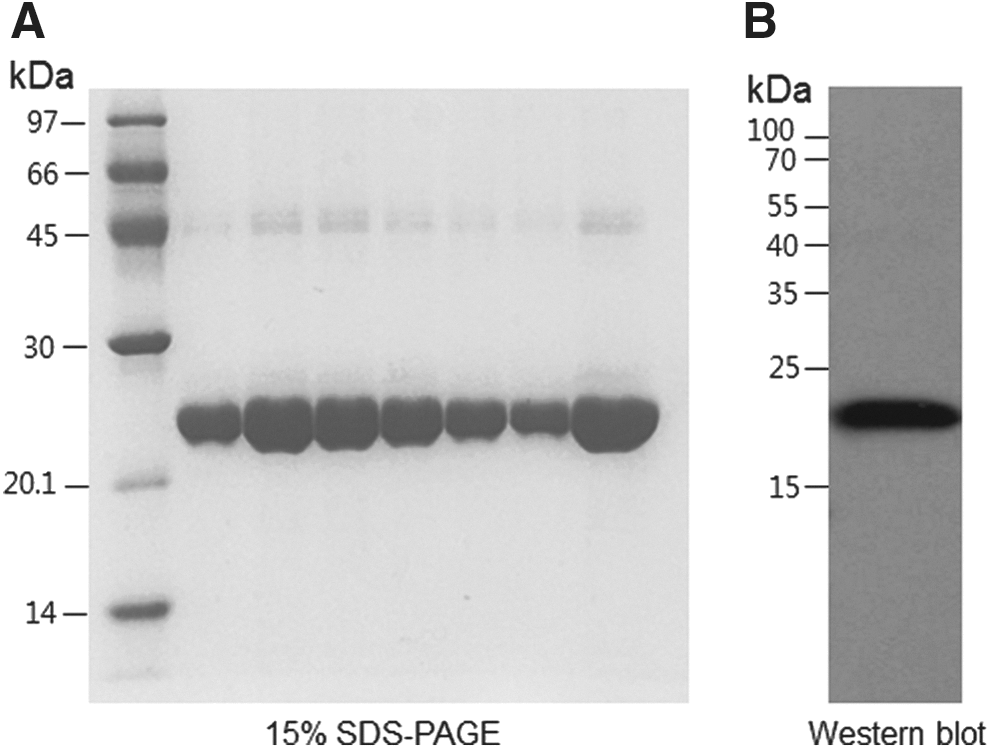
(
In a nondenaturing state, GH-2 showed a single band at around 79 Å in a lipid-free state (Fig. 2), whereas in a lipid-bound state, GH-2 showed two bands at 79 and 82 Å. This result indicates that proteoliposome formation occurred while binding to phospholipids and cholesterol influenced mobility. ApoA-I in a lipid-bound state showed a band at 95 Å, as in our previous report. Synthesized rHDL containing both ApoA-I and GH-2 showed three bands at 96, 82, and 79 Å, indicating formation of rHDL. Extinction coefficient at 280 nm (ε280) was measured as 20,145 based on ultraviolet (UV) spectroscopy using the Suprasil quartz cuvette (1-cm path length), whereas the calculated ε280 of GH-2 was 26,400 using Protean software (v. 5.7.4., Madison, WI). Lipid-free GH-2 and ApoA-I had α-helical contents of 44% and 52%, respectively, as determined by circular dichroism spectroscopy. Using fluorospectroscopy, WMF levels of GH-2 were 349 and 345 nm in lipid-free and lipid-bound states, respectively, whereas ApoA-I showed WMF levels of 344 and 335 nm. These results indicate that the Trp residues in GH-2, 89th and 129th, transitioned to a more hydrophobic state are via phospholipid binding. However, which Trp moved remains unclear.
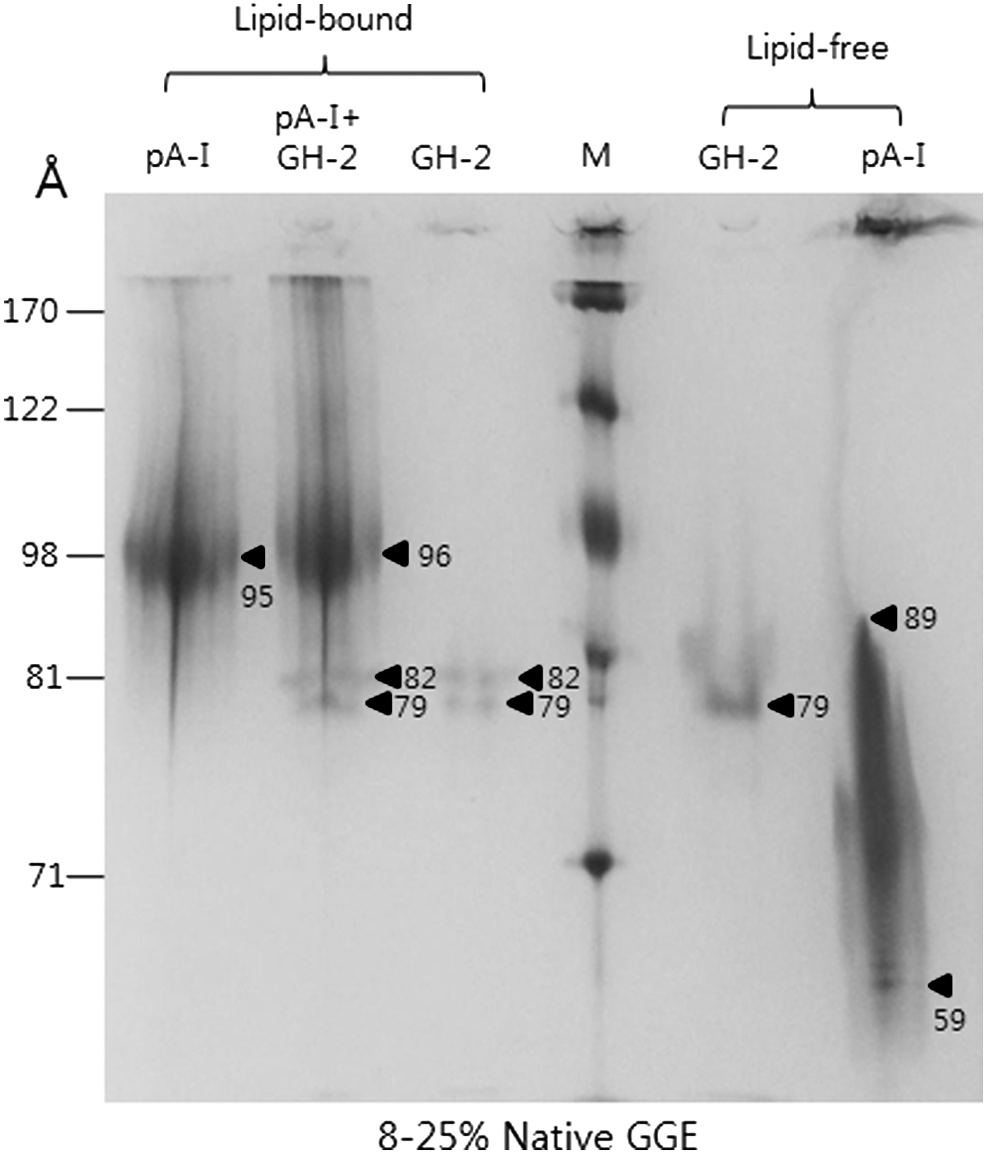
Electrophoretic patterns of growth hormone-2 (GH-2) and apolipoprotein A-I (ApoA-I) in lipid-bound and lipid-free states (8%–25% native polyacrylamide gradient gel electrophoresis [GGE]). Synthesis of reconstituted high-density lipoprotein (HDL) with ApoA-I and GH-2 using palmitoyloleoyl phosphatidylcholine (POPC):cholesterol:protein at a 95:5:1 molar ratio. In a non-denaturing state, each protein was electrophoresed on 8%–25% native gel.
Synthesis of rHDL with GH-2 and ApoA-I
In a lipid-bound state, GH-2 alone showed an α-helical content of 50%, confirming association with phospholipids as well as binding to GH-2. As shown in Fig. 2, ApoA-I in a lipid-bound state showed a band at 95 Å, as in our previous report. Synthesized rHDL containing ApoA-I and GH-2 showed bands at 96, 82, and 79 Å, respectively, indicating formation of rHDL. In a lipid-bound state, GH-2 and ApoA-I-GH-2-rHDL showed WMF of 338 nm, indicating that co-existence of ApoA-I and GH-2 could disrupt Trp exposure in each of the proteins in rHDL.
Phospholipid-binding assay
A dimyristoyl phosphatidylcholine (DMPC) clearance assay revealed that clearance half-times (T1/2) for GH-2 and ApoA-I were 15 and 4 min, respectively, as shown in Fig. 3. Interestingly, GH-2 did not bind efficiently with DMPC until 10 min into the initial phase, and rapid binding was observed at around 15 min. Upon DMPC binding, WMF was blue-shifted 3–4 nm, suggesting Trp moved to a more hydrophobic face to associate with more phospholipids.
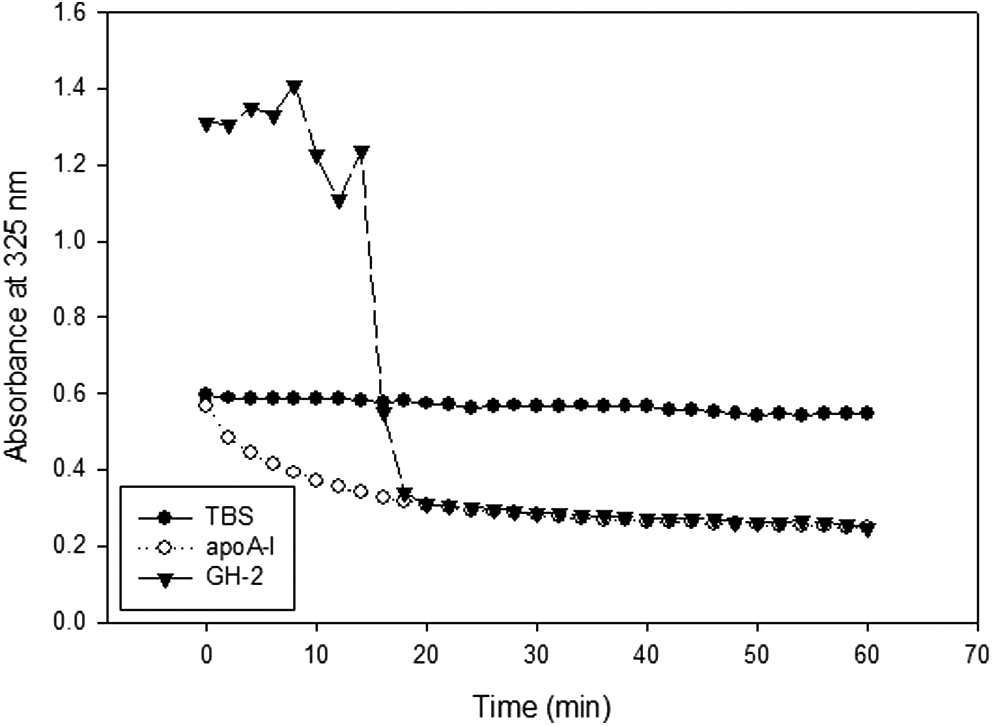
Phospholipid (dimyristoyl phosphatidylcholine)-binding abilities of growth hormone-2 (GH-2) and apolipoprotein A-I (ApoA-I). Absorbance at 325 nm was monitored at 24.5°C at 2-min intervals during 60 min.
Inhibition of oxidation by GH-2
In the presence of Cu2+, oxLDL showed the fastest electromobility due to an increased negative electric charge and ApoB fragmentation. However, lipid-free GH-2 inhibited Cu2+-mediated oxidation, as shown in Fig. 4A. GH-2 treatment (1 and 2 μg; final concentration, 1.7 μM) completely inhibited oxidation similar to the control without Cu2+, indicating that GH-2 could bind to Cu2+ and exert its anti-oxidant effects. Similarly, HDL oxidation mediated by Cu2+ was inhibited by GH-2 up to 1 and 2 μg (0.8 and 1.7 μM, respectively) in the mixture, as shown in Fig. 4B.
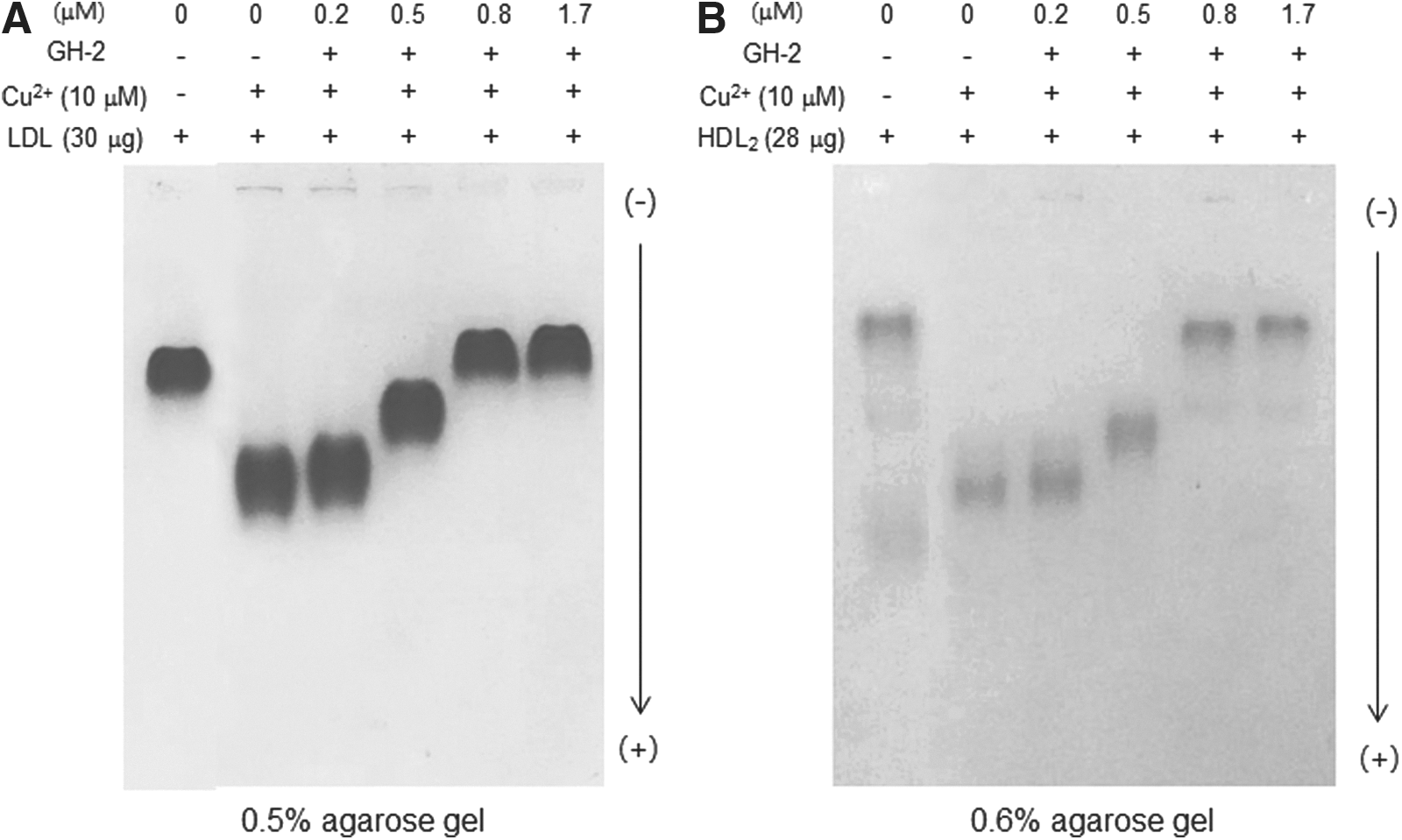
Growth hormone-2 (GH-2) inhibited cupric ion mediated oxidation of low-density lipoprotein (LDL) and high-density lipoprotein (HDL). Comparison of electromobility among LDL (
Inhibition of glycation by GH-2
Fluorescence of the glycated HDL product increased 10.7-fold from the initial level after 72 hr of incubation in the presence of fructose (Fig. 5A), whereas ApoA-I was multimerized with smear band intensity (Fig. 5B). This suggests that glycation occurred via the Maillard reaction. However, GH-2 treatment inhibited glycation of HDL in a dose-dependent manner (42 and 420 nM GH-2 resulted in 33% and 44% inhibition, respectively, compared with fructose-treated HDL). GH-2-treated HDL showed less multimerization in a dose-dependent manner, as shown in Fig. 5B.
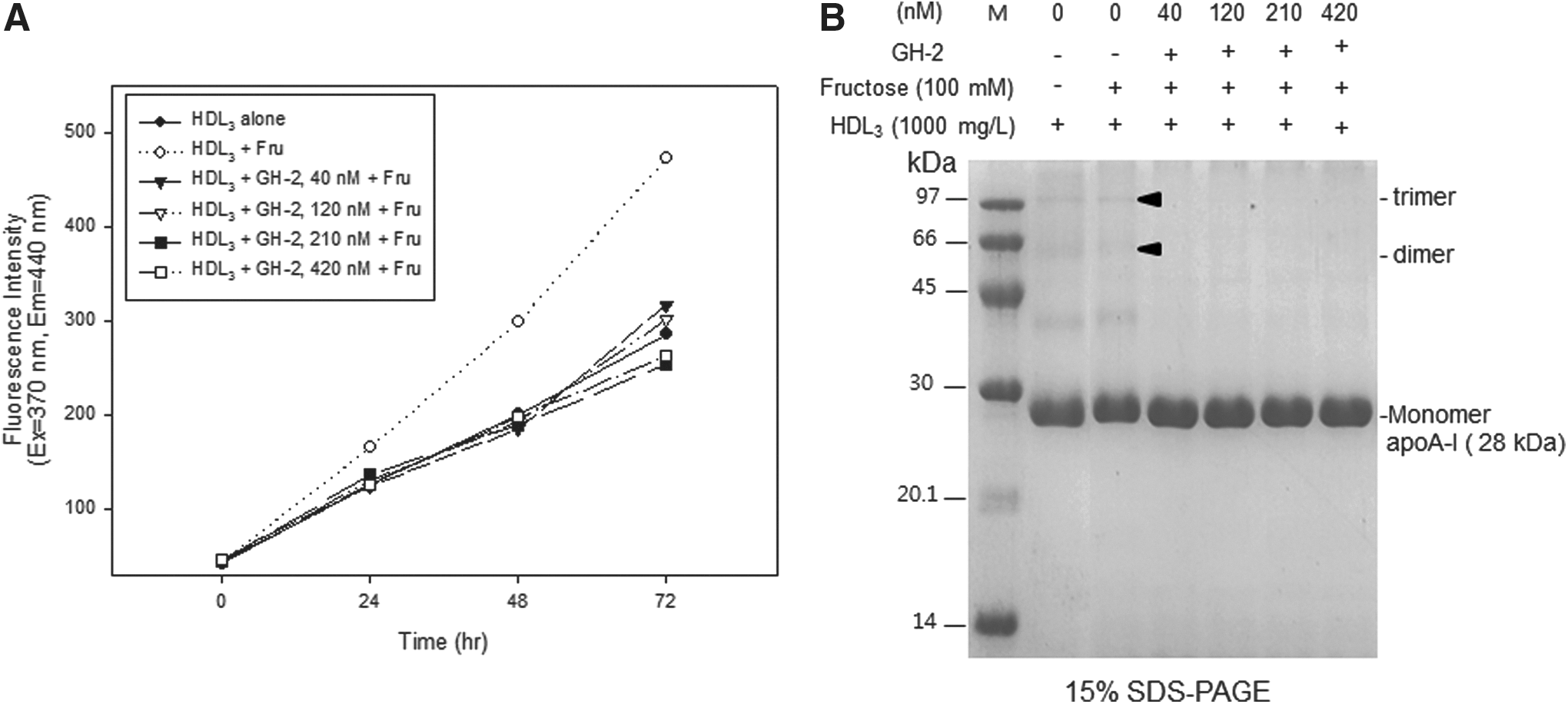
Inhibitory activity of growth hormone-2 (GH-2) against fructose-mediated glycation of high-density lipoprotein (HDL). (
Stabilization of secondary structure by GH-2
As shown in Fig. 6, GH-2 in a lipid-free state had no remarkable change on WMF during isothermal denaturation until 4 M urea treatment around 349 nm; the WMF increased up to 353 nm by a following treatment of 5 M urea. In a lipid-bound state, between 0.5 and 4 M urea, the WMF was increased up to 4 nm red-shift; initial WMF (344 nm) was rapidly elevated up to 348 nm by 4 M urea treatment, suggesting that Trp was very sensitive to lower concentrations of denaturant. In the presence of GH-2, ApoA-I showed no changes in WMF up to 1 M urea, indicating that ApoA-I contributed to the resistance against the denaturation.
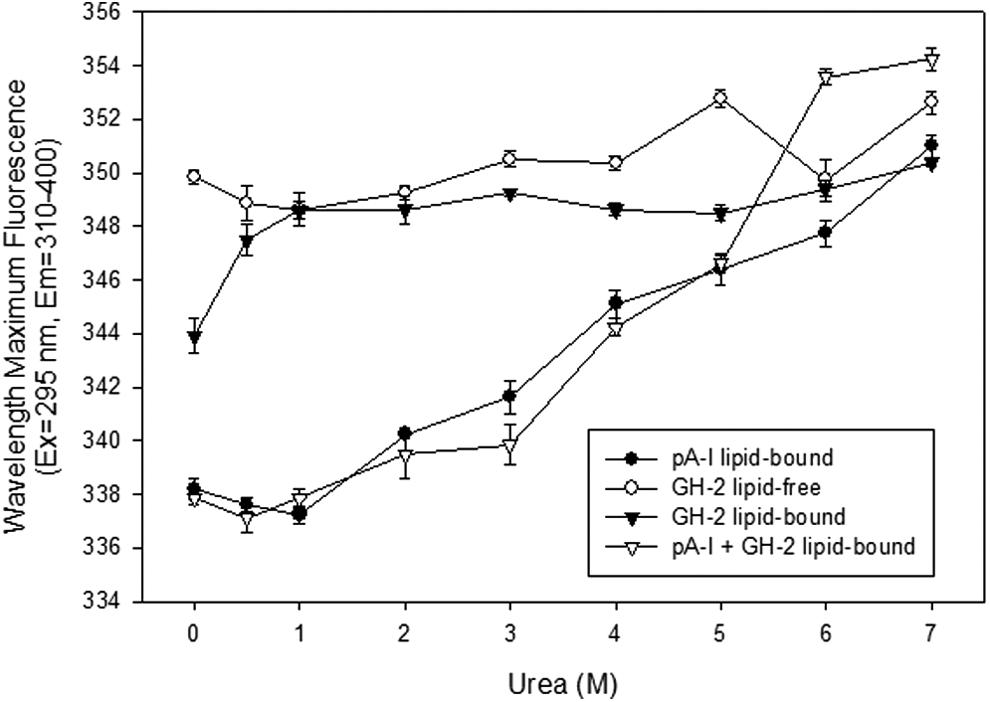
Isothermal denaturation patterns of growth hormone-2 (GH-2) and apolipoprotein A-I (ApoA-I) in lipid-free and lipid-bound states. Fractions of folded proteins were obtained from wavelength maximum fluorescence measurements as a function of urea concentration after equilibration for 72 hr at 4°C.
Inhibition of oxLDL uptake into macrophages by GH-2
Uptake of oxLDL into macrophages was confirmed by Oil Red O staining, as shown in Fig. 7b. However, GH-2 co-treatment resulted in less oxLDL uptake (Fig. 7b–e). The lowest dosage of GH-2 (final concentration, 17 nM; Fig. 7c) resulted in a 21% decrease in Oil Red O staining, whereas the highest dosage (final concentration, 170 nM; Fig. 7f) resulted in a 46% decrease. There was no difference between the 87 nM and 174 nM GH-2 treatments.
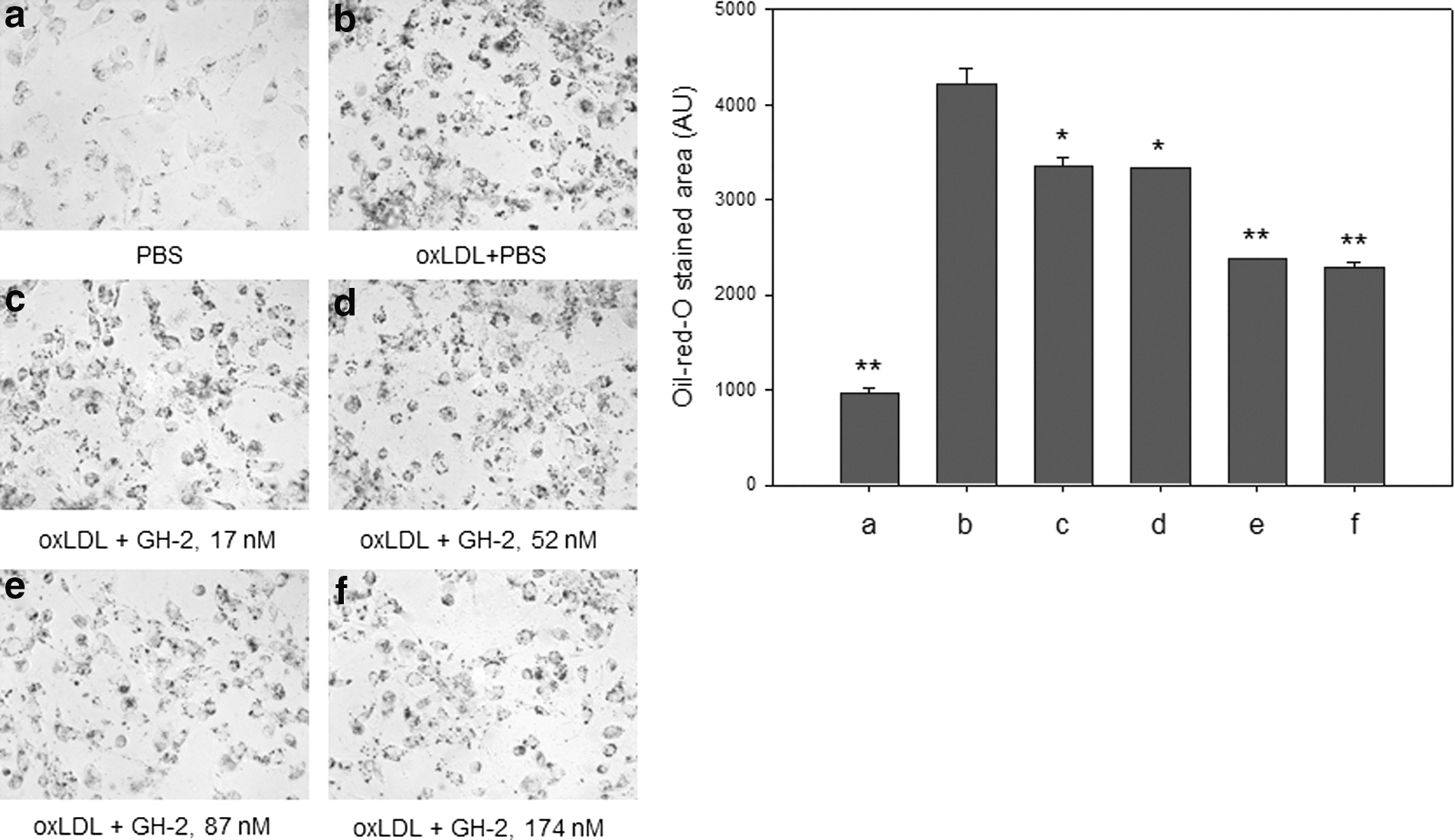
Anti-atherosclerotic activity of growth hormone-2 (GH-2). Oxidized low-density lipoprotein (oxLDL) was applied to differentiated human macrophage cells in the presence of GH-2. Uptake of oxLDL was inhibited by GH-2, as shown by Oil Red O staining. *, p<0.05; **, p<0.01.
Anti-senescence effect of GH-2
HDF cells showed cellular senescence upon incubation with fructose (final concentration, 5 mM), as visualized by blue intensity in SA-β-Gal assay at passage 11 (Fig. 8). However, GH-2 showed potent inhibitory activity in a dose-dependent manner in a lipid-free state. Treatment with GH-2 at 21 nM and 42 nM resulted in 57% and 62% lower blue intensity, respectively. This result indicates that GH-2 has inhibitory activity against cellular senescence, which correlates well with its anti-oxidant (Fig. 4) and anti-glycation activities (Fig. 5).

Anti-senescence activity of growth hormone-2 (GH-2). Cellular senescence was initiated by fructose treatment to human dermal fibroblast (HDF) cells. (
Cell number increased concomitantly with GH-2 dosage; 42 nM GH-2 increased cell numbers 1.7-fold compared to the fructose group as a control.
Wound-healing effect of GH-2
After amputation, ApoA-I or GH-2 was injected into the tail muscles of zebrafish to monitor fin growth over 144 hr, as shown in Fig. 9. In a lipid-free state, ApoA-I showed slightly more enhanced (similar) tissue regeneration activity than GH-2. However, in a lipid-bound state, GH-2 injection resulted in 13% higher tissue regeneration activity than ApoA-I, indicating that phospholipid binding by GH-2 is associated with its wound-healing effect.
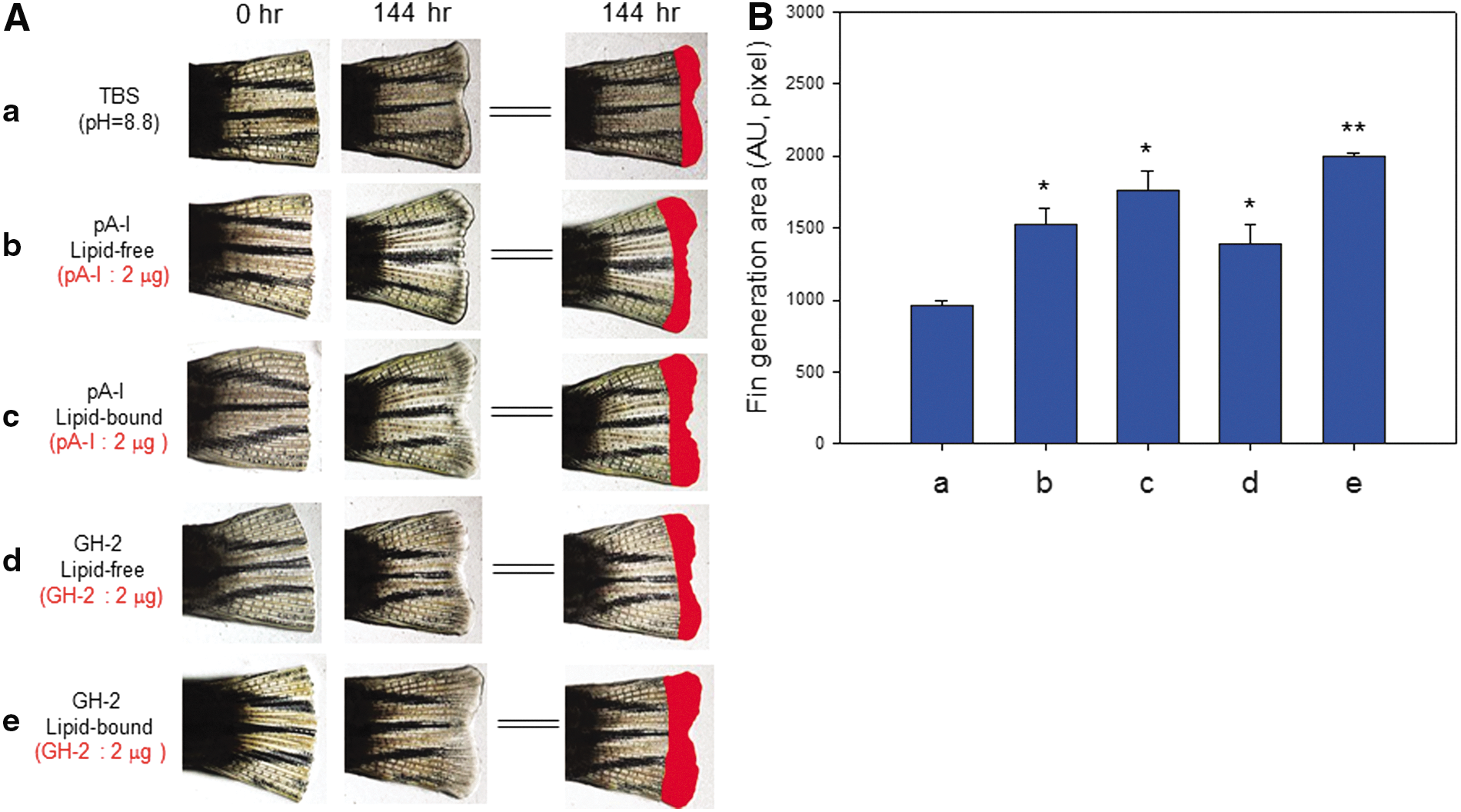
Tissue regeneration activity of growth hormone-2 (GH-2). Enhancement of zebrafish fin regeneration by GH-2 treatment and injection of reconstituted high-density lipoprotein (rHDL) (2 μg of GH-2), apolipoprotein A-I (ApoA-I), and rHDL (2 μg of ApoA-I). (
Injection of GH-2 (2 μg) resulted in two-fold greater fin growth in the lipid-bound state, indicating that the tissue regeneration activity of GH-2 could be enhanced in both lipid and protein formulations. In lipid-free and lipid-bound states, ApoA-I injection caused a similar extent of tissue regeneration. In a lipid-bound state, GH-2 showed up to 1.4-fold higher tissue regeneration activity than ApoA-I, as shown in Fig. 9.
Discussion
The current study characterized the function and structure of GH-2 both in vitro and in vivo. Purified GH-2 showed phospholipid-binding ability, resulting in formation of POPC-proteoliposome via movement of Trp at amino acid positions 89 and 129 and increase of α-helical content, as shown in Table 1 and Fig. 2. These structural changes indicate that Trp transitioned to a more hydrophobic face due to interactions with phospholipids and increased α-helical content. This is very similar to the properties of ApoA-I in a lipid-bound state, as explained in our recent report. 33 Interestingly, a low dose of GH-2 (final concentration, 800 nM) showed potent inhibitory activity against LDL and HDL oxidation (Fig. 4), indicating GH-2 might have potent anti-oxidant activity. In addition, low-dose GH-2 showed inhibitory activity against glycation of HDL in the presence of 100 mM fructose (Fig. 5). These results demonstrate that GH-2 prevents protein modification. Furthermore, its ability to suppress oxidation and glycation might contribute to anti-atherosclerotic (Fig. 7) and tissue regeneration activities (Fig. 9). These results suggest that GH-2 could protect the fetus from oxidation and glycation stresses as well as promote body growth during pregnancy. To our knowledge, this is the first report to characterize the functional and structural characteristics of GH-2 in vitro as well as the protective role of GH-2 against oxidation and glycation in the placenta.
Determined from circular dichroism spectroscopy.
Determined from 8% to 25% native-gradient gel electrophoresis with densitometric scanning analysis.
Generally, the serum level of GH-2 gradually increases up to 20–30 μg/liter in the late stage of pregnancy from an initial level of zero in non-pregnant females. 34 However, there is sufficient information on the physiological functions of GH-2 besides its role in fetal growth. Although the hormonal and regulatory activities of GH-2 during pregnancy are relatively well known, its physiological activities as a protein have been less explored. Because GH-2 is larger than most hormones, it is possible that the protein itself has functionality as an independent protein. The purification yield of GH-2 is low in an E. coli expression system, so biochemical and immunochemical characterization were never successful previously. 35 Solomon et al. reported the large-scale preparation and characterization of GH-V; 22K and 20K variants showed 56% and 41% α-helical content, respectively. 36 Generally, the maternal lipid profile undergoes increases abnormally in total cholesterol (TC) HDL-C, LDL-C, and triacylglycerol (TG) during pregnancy to provide sufficient lipid nutrition. 37 HDL-C and LDL-C levels increased by 18% and 65% in the third trimester compared from the first trimester, respectively, whereas serum TG levels increased 2.9-fold. These changes in the lipid profile indicate increased risk of metabolic burden with modification of lipoproteins. These increases are also related with higher incidence of insulin resistance and risk of gestational DM. 38 It might be possible that GH-2 can alleviate the metabolic stress brought about by higher lipid and carbohydrate contents and suppress incidence of preeclampsia.
Interestingly, women with gestational DM show significantly lower LDL-C than controls during pregnancy as well as higher serum TG levels. 39 Furthermore, women with pre-eclampsia and hypertension are known to have 17% higher TG content as well as 25% lower HDL-C than controls. These women also show 20% higher ApoB and 50% higher malondialdehyde (MDA) levels. 40 Therefore, GH-2 might exert resistance against oxidative stress in normal pregnant women. Mittal et al. reported that the serum GH-2 level is up to two-fold higher in women suffering from severe pre-eclampsia (23,076 ng/liter) compared to control women (12,157 ng/liter). Umbilical serum GH-2 concentration is also higher in pre-eclamptic women than in controls.
Using the Protean DNASTAR analysis software (v. 5.7.4, Madison, WI), the isoelectric point (pI) of GH-2 was calculated as 7.04, whereas DNA sequencing has shown the pI to be 8.9. 41 However, our isoelectrofocusing (IEF) experiment revealed that the pI of GH-2 was around 7.0, which is in good agreement with the Protean software analysis.
The purification yield of GH-2 was 10–15 mg/liter of culture, and hGH-V has been expressed from E. coli HB101 and purified by ion-exchange and exclusion chromatography. 35 Our expression and purification method using a pET vector and Ni2+ column chromatography was very efficient, with high yield and purity. Although its physiological role remains unknown, GH-2 is increased in the maternal and fetal sera of patients with pre-eclampsia. Because GH-2 should only be circulated in the maternal serum, it is possible to postulate that GH-2 contributes to the protection of maternal serum proteins against the glycation stress in pre-eclampsia. Future studies should be carried out to elucidate which Trp residue (position 89 or 129) is more responsible for changes in WMF.
There has been no report to elucidate the anti-oxidant activity of the GH-2, especially in the lipid-bound state. To enhance the physiological activity of GH-2, we synthesized reconstituted HDL containing GH-2 to compare anti-oxidant and anti-glycation activity. Incorporation of GH-2 into rHDL can enhance structural stability and tissue regeneration efficiency in vertebrate models, indicating a synergetic effect between GH-2 and ApoA-I in rHDL.
In conclusion, GH-2 has anti-oxidant and anti-glycation activities in vitro, and incorporation of GH-2 into proteoliposomes enhances structural stability and tissue regeneration efficiency in vertebrate models.
Footnotes
Acknowledgments
This work was supported by the Mid-carrier Researcher Program (2014-11049455) and Basic Science Research Program (2010-020910) through the National Research Foundation of Korea (NRF).
Author Disclosure Statement
No competing financial interests exist.