
Abstract
This study explored the curative effect and underlying mechanisms of a traditional Chinese medicine compound prescription, Bushen-Yizhi formula (BSYZ), in ibotenic acid (IBO)-induced rats. Morris water maze and novel object recognition tests showed that BSYZ significantly improved spatial and object memory. Brain immunohistochemistry staining showed that BSYZ significantly up-regulated expression of choline acetyltransferase (ChAT) and nerve growth factor (NGF) in the hippocampus and cortex. The protein tyrosine kinase high-affinity receptor TrkA was slightly increased in the hippocampus and cortex, and significantly enhanced in the nucleus basalis of Meynert (NBM) after BSYZ intervention. The immunoreactivity of the p75 low-affinity receptor in BSYZ-treated rats was significantly strengthened in the cortex. Similar expression trends of nerve growth factor (NGF), TrkA, and p75 mRNA were observed in the hippocampus and cortex. Additionally, BSYZ reversed IBO-induced disorders of acetylcholine (ACh) levels, ChAT, and cholinesterase (ChE) in the cortex, which was consistent with the changes in mRNA levels of ChAT and acetylcholinesterase (AChE). Expression of ChAT and AChE proteins and mRNA in the hippocampus was up-regulated, whereas the apoptosis-relative protein cleaved caspase-3 was decreased after administration of BSYZ. Moreover, changes in cell death were confirmed by histological morphology. Thus, the results indicated that the BSYZ formula could ameliorate memory impairments in IBO-induced rats, and it exerted its therapeutic action probably by modulating cholinergic pathways, NGF signaling, and anti-apoptosis. Overall, it is suggested that the BSYZ formula might be a potential therapeutic approach for the treatment of Alzheimer's disease (AD) and other cholinergic impairment-related diseases.
Introduction
C
Nerve growth factor (NGF), through its two typical receptors the protein tyrosine kinase high-affinity receptor (TrkA) and p75 low-affinity receptor (p75), plays an important role in neuronal survival and growth. TrkA knockout in animals might have a selective impact on cognitive activity. 5,6 p75 positively regulated NGF-triggered TrkA signaling pathways that mediate neuronal survival in hippocampal neurons. 7 It should be noted that p75 might also modulate cell death. Overexpression of p75 contributed to the excitotoxin-induced death of rat basal forebrain cholinergic neurons by an apoptotic-like mechanism. 8 Although the mechanisms are manifold, NGF still might be a promising target for the treatment of dementia. Outcomes of the first application of NGF intracranial infusion in an AD patient indicated that verbal episodic memory and cholinergic deficits were improved with no adverse effects. 9 NGF-infused rats showed enhanced recognition memory, increased expression of NGF and TrkA, as well as increased cell proliferation in the dentate gyrus. 10
Bushen-Yizhi formula (BSYZ) is a traditional Chinese medicine compound recipe containing She Chuang Zi (Cnidium monnieri (L) Cuss., fruit), Ren Shen (Panax ginseng C. A. Mey., rhizome), Zhi He Shou Wu (preparata of Polygonum multiflorum Thuna., radix), Mu Dan Pi (Paeonia suffruticosa Andr., cortex), Nv Zhen Zi (Ligustrum lucidum Ait., fruit), and Gou Qi (Lycium barbarum L., fruit) (Ait. patent no. ZL 200610112916.1). A multi-center, randomized, double-blind, controlled clinical trial of BSYZ for treatment of AD indicated significant increase of Mini-Mental State Examination (MMSE) scores after intervention with BSYZ.
11
In our previous studies, we established a rat dementia model using
Materials and Methods
Preparation of BSYZ extracts
She Chuang Zi (Cnidium monnieri (L.) Cuss., fruit), Ren Shen (Panax ginseng C. A. Mey., rhizome), Zhi He Shou Wu (preparata of Polygonum multiflorum Thuna., radix), Mu Dan Pi (Paeonia suffruticosa Andr., cortex), Nv Zhen Zi (Ligustrum lucidum Ait., fruit), and Gou Qi (Lycium barbarum L., fruit) were purchased from Guangxi Yifang Chinese Herbal Medicine Department and identified by Professor Chen Jiannan, pharmacognosist of the School of Chinese Materia Medica, Guangzhou University of Chinese Medicine. A voucher specimen was deposited at Guangzhou University of Chinese Medicine with the registration number 20121209. The BSYZ extracts were provided by School of Chinese Materia Medica, Guangzhou University of Chinese Medicine and stored at 4°C until used. The doses of BSYZ extracts were expressed as grams of the original raw herbs per kilogram of body weight.
The proportion of six raw herbs was 3:3:2:2:2:2. The extraction process comprised three parts: (1) She Chuang Zi was refluxed twice (for 2 hr each) with 2.5 L of 95% ethanol, respectively. The filtrate was reclaimed by ethanol and stewed for overnight to collect the precipitates. (2) Mu Dan Pi was distilled for 3 hr to gather the distillate, and the residue was refluxed twice (for 1.5 hr each) with 3 L and 2.5 L, successively. The filtrate was subjected to precipitation with ethanol to obtain the supernatant. (3) The other four herbs were refluxed twice (for 2 hr and 1.5 hr, successively) with 2 L of 80% ethanol; after cooling to room temperature, the combined extract was filtered and condensed. All of the concentrated extracts were dried to obtain powder by spray-drying before adding the fine power of She Chuang Zi precipitates. Finally, the mixture was mixed with an amount of medical starch and loaded into capsules.
Reverse-phase high-performance liquid chromatography analysis of BSYZ's chemical composition
The BSYZ formula was qualitatively analyzed by high-performance liquid chromatography (HPLC) using a Dionex Summit HPLC system equipped with a PDA-100 detector, a P680 pump, an ASI-100 automatic sampler, and a STH585 thermostatic column compartment. As illustrated in Fig. 1, the eight compounds of the main peaks of the HPLC map were identified as paeoniflorin (peak 1, tR8.66 min), 2,3,5,4′-tetrahydroxystilbene-2-O-β-
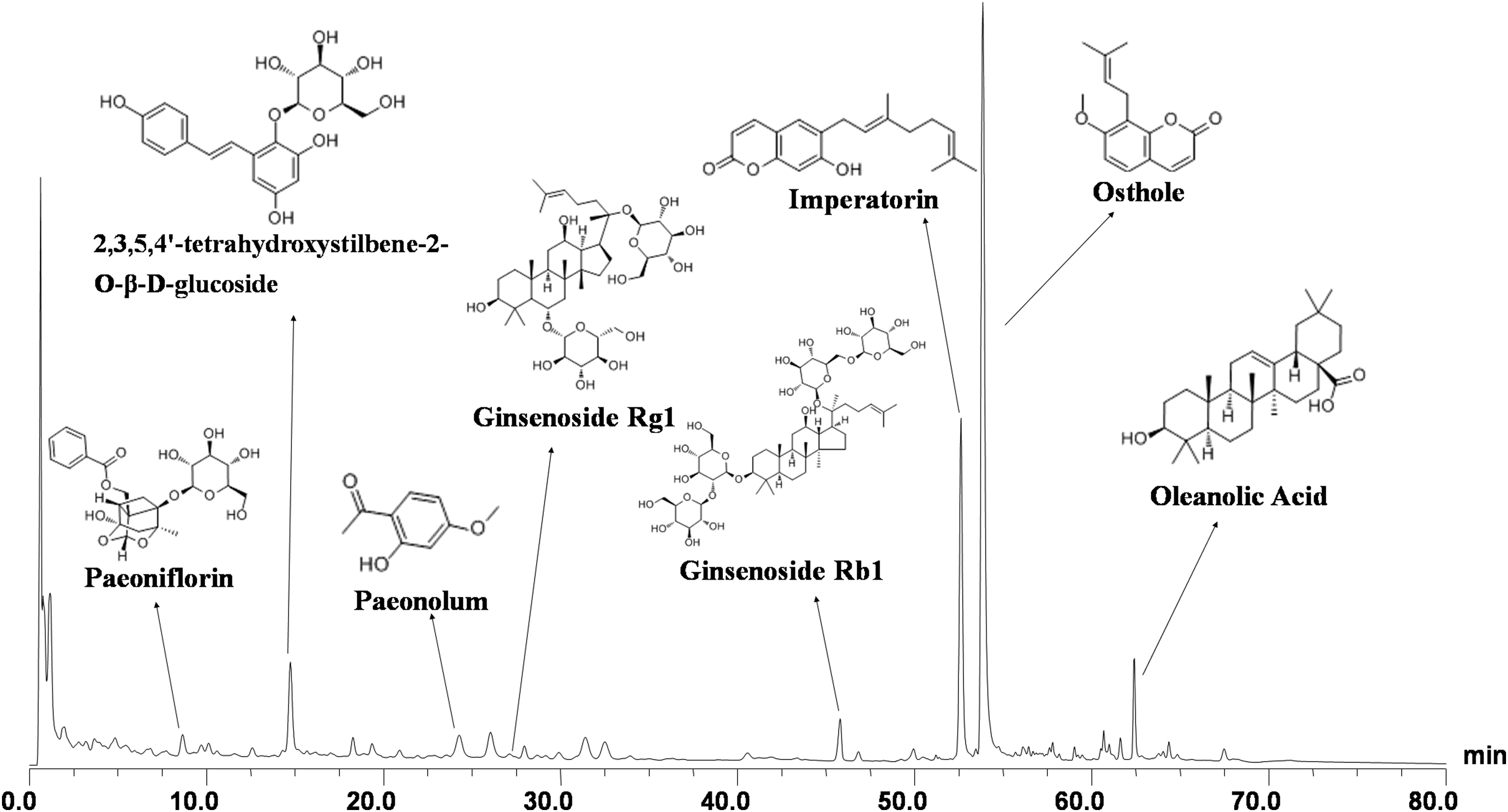
Chemical profile of Bushen-Yizhi Formula (BSYZ) analyzed by high-performance liquid chromatography (HPLC). Ten batches of BSYZ were analyzed by HPLC with ultraviolet (UV) detection and similar profiles were observed. The samples showed good correlation and shared a similar chromatographic pattern with the similarity indexes higher than 0.986. The analysis was performed with a Gemini-C18 110A (150 mm×2.00 mm, 5 μm) at 35°C. Eight compounds were eluted (eluent A, acetonitrile; eluent B, water-phosphoric acid, 100:0.1, vol/vol) at a flow rate of 0.8 mL/min using a gradient program.
Animals and housing
Sprague–Dawley rats (3-month-old, weighing 200–250 grams, male) were purchased from the Experimental Animal Center of Guangzhou University of Chinese Medicine (Guangzhou, China), and housed in a room at constant temperature (22±2°C) and under a 12-hr light/dark cycle. Food and water were freely available. All procedures were undertaken as required by the animal ethics committee of Guangzhou University of Chinese Medicine and in accordance with the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health.
Surgery and drug administration
Animals were randomly divided into five groups: Sham group (bilateral NBM lesion by saline), IBO group (bilateral NBM lesion by IBO), low-dose BSYZ group (IBO+BSYZ 1.46 grams/kg per day), middle-dose BSYZ group (IBO+BSYZ 2.92 grams/kg per day), and high-dose BSYZ group (IBO+BSYZ 5.84 grams/kg per day). Rats were anesthetized with sodium pentobarbital (50 mg/kg, intraperitoneally [i.p.]) and fixed on a stereotaxic apparatus (RWD Life Science Co., Ltd. Shenzhen, China). IBO (International Laboratory, USA) was dissolved in normal saline at a concentration of 5 μg/μL. For the IBO and BSYZ groups, 1 μL of IBO solution (5 μg/μL) was infused to both sides of the NBM (A 1.0 mm; L±2.7 mm; V 7.8 mm) of rats 16 using a microsyringe for 5 min and then left in place for 5 min. Rats in the sham group received the same volume of normal saline instead of IBO. Rats were allowed 3 days to recover after the surgery and received oral administration of BSYZ at different doses (1.46, 2.92, and 5.84 grams/kg) and normal saline (sham and IBO groups) once a day for 4 consecutive weeks.
Behavioral tests
Before behavioral tests were undertaken, rats were habituated to the environment of the testing room for 3 days. All tests were conducted during the light cycle. The Morris water maze test and novel object recognition test were carried out to evaluate the spatial memory and object memory of rats.
Morris water maze test
The equipment for the Morris water maze test was a black circular pool (180 cm in diameter and 50 cm in height), a camera monitor (above the center of the pool), and a video-tracking system (Guangzhou Feidi Biological Technology Co. Ltd., China). The water (30 cm in depth, 22–26°C) was opacified with non-toxic black paint. The pool was divided into four equal quadrants, one containing the platform and the others including four starting points.
On days 1–5, a black circular platform (10 cm in diameter) was kept in the center of one quadrant and submerged 1 cm below the surface of the water. During this procedure, four 60-sec trials were administered each day from four different start points. The escape latency (the time to find the platform) was recorded. If the rat failed to find the platform within 60 sec, it was guided toward the platform, and the escape latency was recorded as 60 sec. After staying on the platform for 20 sec, the next trial was continued. At the end of the last trial of each day, the rat was dried and returned to its home cage. On day 6, the platform was removed and each rat was allowed to swim freely for 60 sec from the same position, opposite to the platform location. The number of crossings of the previous platform position was recorded.
Novel object recognition test
The apparatus for the novel object recognition test consisted of a bright testing arena (50 cm long, 50 cm wide, and 50 cm high) and a video camera (above the center of the arena) that was connected to a computerized video tracking system (Shanghai Xinruan Information Technology Co. Ltd. China).
The novel object recognition test was carried out as described 17 with minor modifications. Briefly, on day 1, rats were acclimated to the empty testing arena for 10 min in turns, and subsequently returned to their home cage for 24 hr. On day 2, there were two stages. For the first stage, two identical objects were affixed to opposite positions (northeast and northwest) of the arena and rendered immovable. Rats were allowed to explore freely for 5 min and returned to their home cages for a 1-hr inter-stage interval. Any rats that failed to reach the encoding criterion within 5 min were removed from the study. For the second stage, one of the familiar objects was replaced by a novel object, and rats were placed back into the arena for 5 min. The time spent on exploring the familiar object (Tf) and novel object (Tn) were recorded with the rat's snout within 2 cm of the object. The objects and arena were thoroughly cleaned between each trial. Object recognition memory was analyzed by calculating discrimination index and novel object preference index: Discrimination index=(Tn−Tf) / (Tn+Tf), 18 novel object preference index=Tn / (Tn+Tf). 19
Brain sections and tissue preparation
After finishing the behavioral tests, three rats in each group were selected randomly, anesthetized with sodium pentobarbital, and decapitated. Brains were removed, fixed in 4% paraformaldehyde in 0.1 M phosphate buffer (phosphate-buffered saline [PBS], pH 7.4) for 24 hr, dehydrated with a graded series of ethanol, and embedded in paraffin blocks. Coronal sections were cut at 4-μm thickness for immunohistochemistry and histological examination. The rest of the rats were anesthetized and decapitated. Brains were rapidly removed on ice and stored at −80°C until analysis.
Immunohistochemistry
Sections were dewaxed and rehydrated in xylene/alcohol serial solutions. Following blocking with 3% hydrogen peroxide for 10 min and normal goat serum for 10 min at 37°C, sections were incubated with anti-choline acetyltransferase (ChAT) antibody (1:200, Proteintech™, USA), anti-NGF antibody (1:200, Abcam, UK), anti-TrkA antibody (1:200, Abcam, UK), and anti-p75 antibody (1:200, Abcam, UK) at 4°C overnight. After washing in PBS three times, sections were incubated with horseradish peroxidase-conjugated secondary antibodies (1:2000, Cell Signaling Technology, USA) for 1 hr at 37°C. Following a reaction with diaminobenzidine (Tiangen Biotech Co, Ltd., China), the sections were dehydrated, cleared in ascending alcohol/xylene solutions, and cover slipped with permanent mounting medium. Sections were imaged using a Leica DM4000B Microscope (Leica, Germany) connected to a computerized system with Leica QWin Plus software (Leica, Germany). The mean positive staining area percentage was measured at an original magnification of 200×by using LEICA QWIN plus software.
Histological examination
Sections were used for Hematoxylin & Eosin (H&E), Nissl staining, and terminal deoxynucleotidyl transferase dUTP nick end labeling (TUNEL) assay according to the manufacturer's instructions. A Haematoxylin-Eosin Kit (Boster, China), Nissl Staining Solution (Beyotime Institute of Biotechnology, China), and FragEL™ DNA Fragmentation Detection Kit (Merck, USA) were used.
Enzyme-linked immunosorbent assay quantification of acetylcholine concentrations
For the quantitative determination of acetylcholine (ACh) concentration, the cortex was used according to the manufacturer's instructions for the Rat Acetylcholine ELISA kit (CUSABIO Biotech Co., Ltd., China).
Biochemical analysis
For assays of ChAT and cholinesterase (ChE) activities, the cortex was homogenized with ice-cold saline to make a 10% (wt/vol) homogenate. After centrifugation at 3000×g for 10 min at 4°C, the supernatant was collected for determination according to the manufacturer's instructions of the commercial kits (Nanjing Jiancheng Institute of Biological Engineering, Nanjing, China). The absorbance was read using a Universal Microplate Spectrophotometer (Bio-Rad, USA).
Real-time PCR
Brain samples (hippocampus and cortex) were homogenized and RNA was extracted using RNAiso Plus (Takara Biotechnology [Dalian] Co., Ltd. China). Total RNA was quantified with the NanoDrop 2000 spectrophotometer (Thermo Scientific, MA) and reverse transcribed into complementary DNA using PrimeScript™ RT Master Mix-Perfect Real Time (Takara Biotechnology [Dalian] Co., Ltd. China) according to the manufacturer's instructions. Real-time quantitative PCR was performed with SYBR® Premix Ex Taq™ II (Tli RNaseH Plus) (Takara Biotechnology (Dalian) Co., Ltd. China) and the CFX96™ real-time PCR Detection System (Bio-Rad). Each sample was analyzed in triplicate reactions of 25 μL with the following protocol: 95°C for 30 sec, 40 cycles of 95°C for 5 sec, and 62°C for 30 sec. The following primers were designed and synthesized by Life Technologies: ChAT, forward, 5′-GTGAGACCCTGCAGGAAAAG-3′, and reverse, 5′-TGGTCATTGGTGTCTTGGAA-3′; AChE, forward, 5′-CAGCAATACGTGAGCCTGAA-3′, and reverse, 5′-TTCCAGTGCACCATGTAGGA-3′; β-actin, forward, 5′-GTCGTACCACTGGCATTGTG-3′, and reverse, 5′-CTCTCAGCTGTGGTGGTGAA-3′. Primers targeting NGF, TrkA, and p75 were designed from the published gene sequences and synthesized by Life Technologies: NGF, forward, 5′-TGCATAGCGTAATGTCCATGTTG-3′, and reverse, 5′-CTGTGTCAAGGGAATGCTGAA-3′) 20 ; TrkA, forward, 5′-CCATGCTACAGCACCAACAC-3′, and reverse, 5′-AAGGACCAGGAGCCACATC-3′ 21 ; p75, forward, 5′-TGCACAGCGACAGTGGCATC-3′, and reverse, 5′-TCCTCACGCTTGGTCAGGGG-3′. 22 β-actin was used as internal control. mRNA expression was analyzed using CFX Manager™ Software provided by the CFX96™ Real-Time PCR Detection System (BioRad Laboratories, Inc.).
Western blotting
Brain samples (hippocampus) were homogenized in ice-cold RIPA buffer (Nanjing Bio-Box Biotechnology, Inc., China) with protease and phosphatase inhibitors. The lysates were incubated for 15 min on ice and centrifuged at 12,000×g for 15 min at 4°C. The supernatant was collected, and the protein determination was carried out by using a bicinchoninic acid (BCA) assay kit (Beyotime Institute of Biotechnology, China). Equal amounts of protein (40 μg) for each sample were loaded onto 12% sodium dodecyl sulfate (SDS)-polyacrylamide gels. Proteins were separated and transferred to polyvinylidene difluoride membranes (Millipore Corporation, USA). After blocking with 5% non-fat milk in Tris-buffered saline containing 0.1% Tween 20 (TBST) for 1 hr at room temperature, the membranes were incubated with primary antibodies (diluted in 5% bovine serum albumin [BSA]) overnight at 4°C. Anti-ChAT antibody (1:1000, Proteintech™, USA), anti-AChE antibody (1:1000, Bioworld Technology, Inc., USA), anti-caspase-3 antibody (1:1000, Cell Signaling Technology, USA), and anti-β-actin antibody (1:2000, Cell Signaling Technology, USA) were used. After three washes with TBST, membranes were incubated in horseradish peroxidase-conjugated anti-rabbit immunoglobulin G (IgG) antibody (1:2000, Cell Signaling Technology, USA) for 1 hr at room temperature and washed with TBST three times again. Images of immunoreactive bands based on an electrochemiluminescent reagent (Bio-Rad, USA) were obtained using a FluorChem FC3 imager (Alpha, USA) and quantified using ImageJ software (NIH Image, USA). The amount of each protein was normalized for the amount of the corresponding β-actin detected in the sample.
Statistical analysis
All statistical analysis was performed using Statistic Package for Social Science (SPSS) v. 17.0 software. In the Morris water maze test, differences in escape latency were analyzed by repeated measures analysis of variance (ANOVA) followed by least significant difference (LSD) post hoc test. Other data were analyzed by one-way ANOVA followed by the LSD post hoc test or nonparametric test where appropriate. Differences were considered statistically significant when values of p were<0.05.
Results
Effects of BSYZ on learning and memory impairments induced by IBO
In the Morris water maze test, IBO rats spent a significantly longer time to find the hidden platform than rats in the sham control group from day 2 to day 5 (day 2, p<0.05; day 3 to day 5, p<0.01) (Fig. 2A). Rats treated with both low and high doses of BSYZ spent significantly shorter escape latencies than IBO-induced rats on day 4 (p<0.05) and day 5 (p<0.05 for low-dose BSYZ, p<0.01 for high-dose BSYZ) (Fig. 2A). In the spatial probe trial, the times crossing the platform position in IBO-injected rats were significantly lower than that in sham group (p<0.01), whereas a significant increase in times was observed in high doses of BSYZ-treated rats (p<0.05) (Fig. 2B).
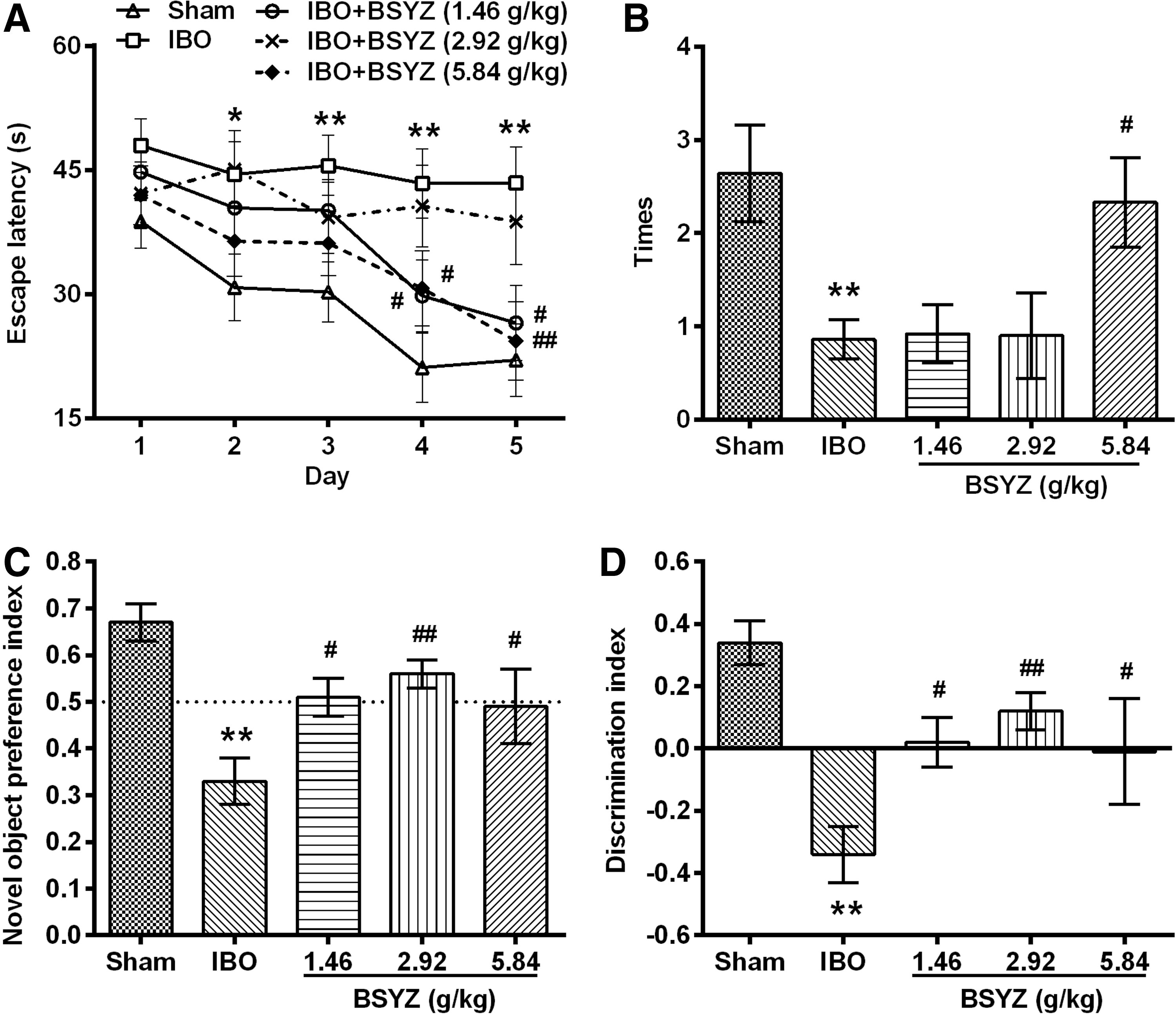
Effects of Bushen-Yizhi Formula (BSYZ) on learning and memory impairments induced by ibontenic acid (IBO).
In the novel object recognition test, IBO-injected rats showed significantly lower levels of novel object preference index (p<0.01) (Fig. 2C) and discrimination index (p<0.01) (Fig. 2D) than the sham group. BSYZ treatment rats showed significantly higher levels of these two indexes than IBO-induced rats (p<0.05 for low- and high-dose groups, p<0.01 for middle-dose group) (Fig. 2C, D).
Effects of BSYZ on ChAT, NGF, TrkA, and p75 immunoreactivity in brain
As shown in Fig. 3A and Fig. 4A, ChAT expression in IBO rats was significantly reduced in the hippocampus, cortex, and NBM as compared to the sham group (p<0.01, p<0.01, and p<0.05, respectively). BSYZ significantly enhanced ChAT immunoreactivity in the hippocampus (p<0.01 for all dose groups) and cortex (p<0.01 for all dose groups) compared with the IBO group. An obvious change in the size of ChAT-positive staining cells was observed in the NBM region, and a trend in increasing ChAT expression was detected in BSYZ-treated rats. There were no significant changes of ChAT expression in the thalamus among groups.
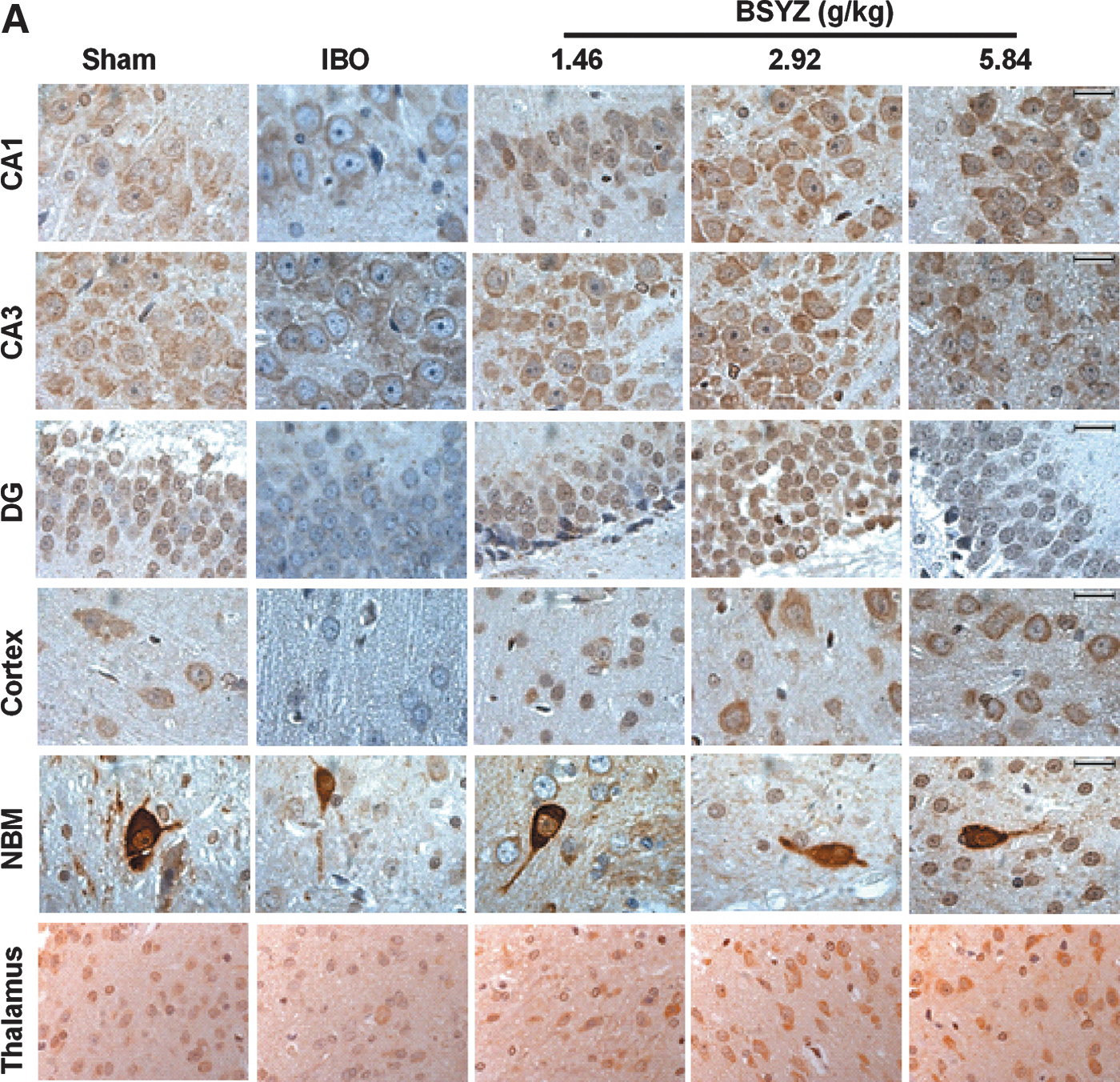
Photomicrographs of the immunohistochemical determination of choline acetyltransferase (ChAT), nerve growth factor (NGF), tyrosine kinase high-affinity receptor TrkA, and p75 low-affinity receptor in rat brains. (


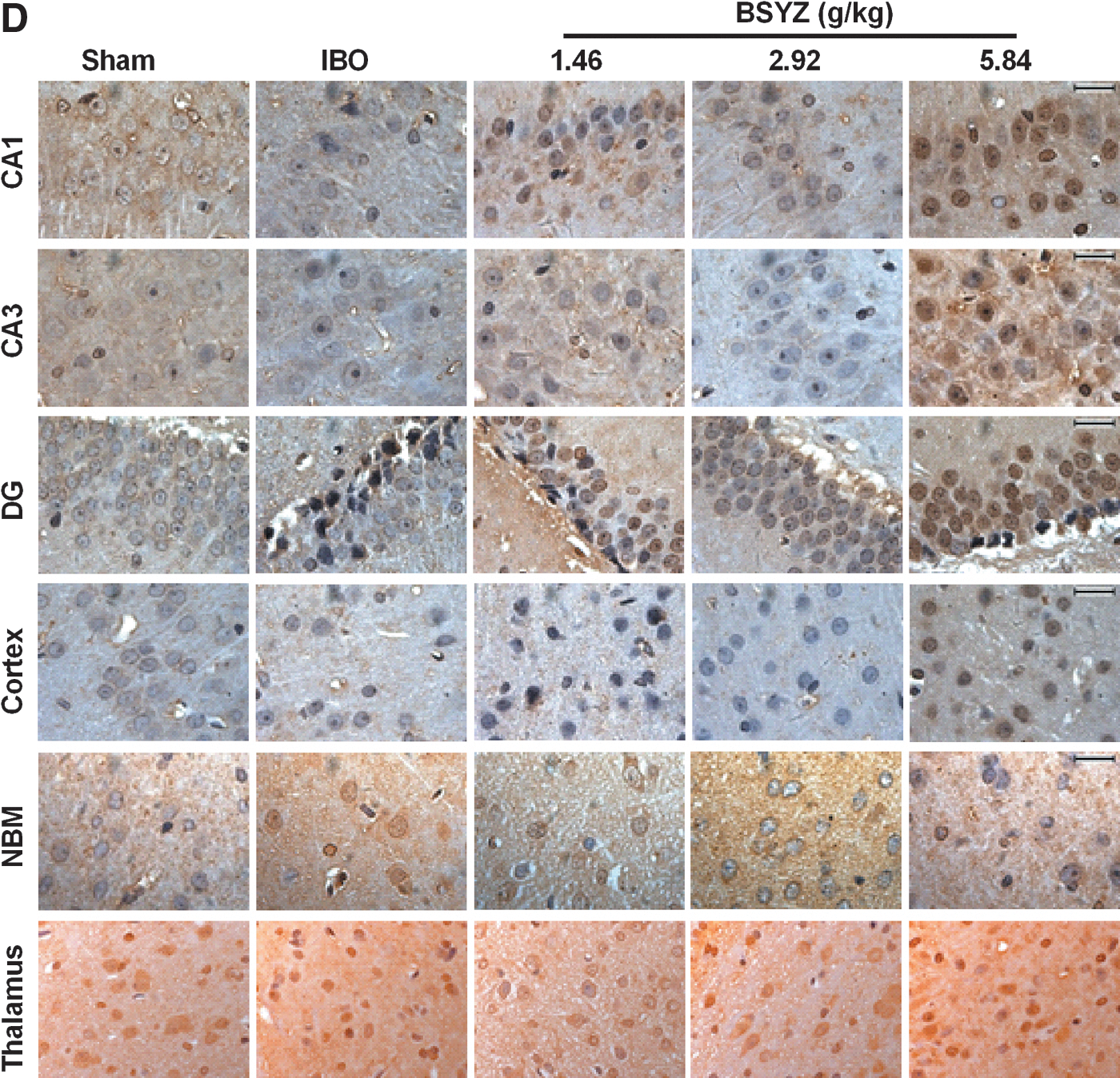
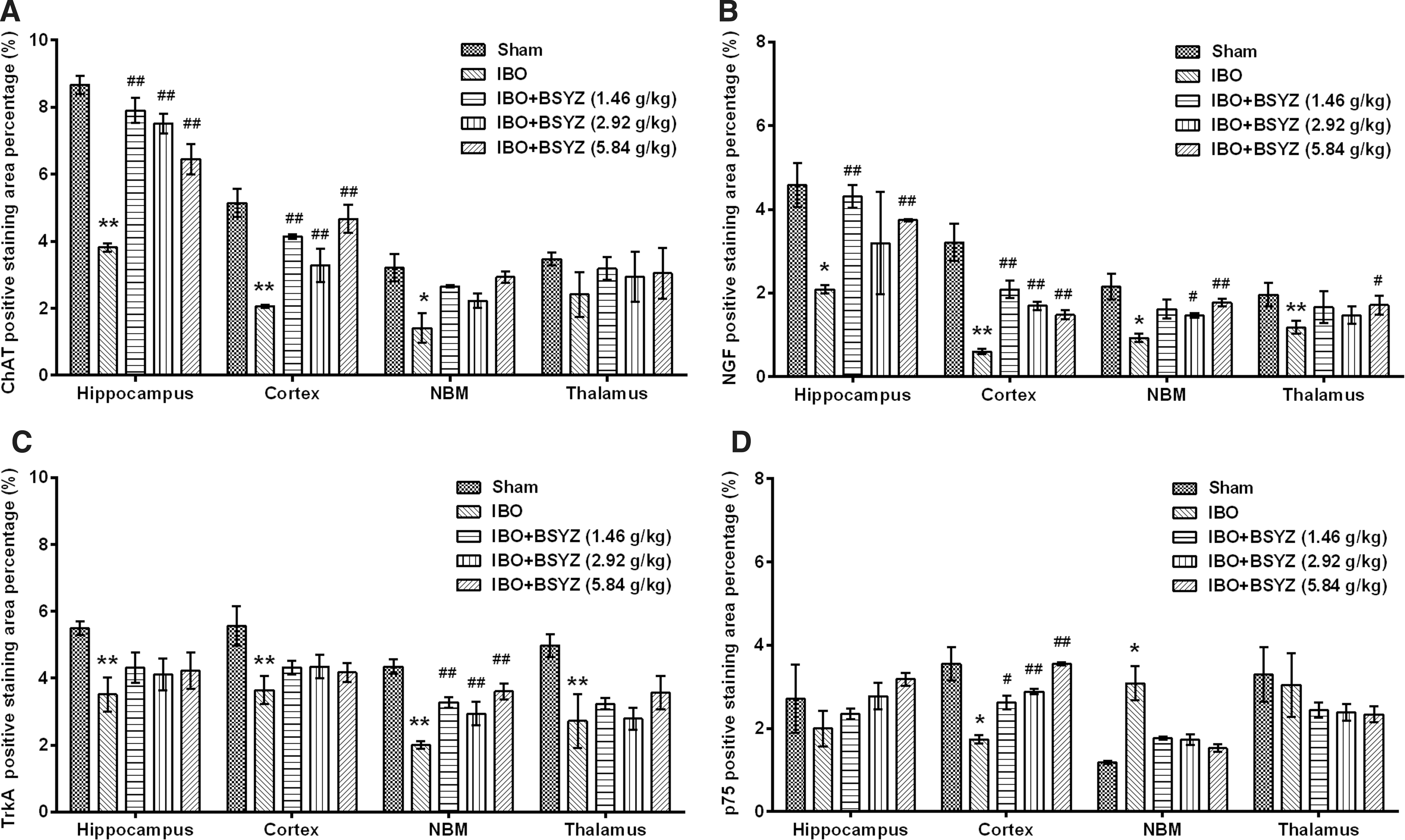
Effects of Bushen-Yizhi Formula (BSYZ) on choline acetyltransferase (ChAT), nerve growth factor (NGF), TrkA, and p75 expression in the brain by immunohistochemistry. (
As shown in Fig. 3B and Fig. 4B, NGF expression of IBO rats was significantly reduced in the hippocampus, cortex, NBM, and thalamus as compared to the sham group (p<0.05, p<0.01, p<0.05, and p<0.01, respectively), whereas BSYZ treatment significantly increased NGF expression in the hippocampus (p<0.01 for low and high dose groups), cortex (p<0.01 for all dose groups), NBM (p<0.05, and p<0.01 for middle- and high-dose group, respectively), and thalamus (p<0.05 for the high-dose group) compared with the IBO group.
As shown in Fig. 3C and Fig. 4C, TrkA expression of IBO rats was significantly reduced in the hippocampus, cortex, NBM, and thalamus as compared to the sham group (all p<0.01), whereas BSYZ treatment significantly strengthened TrkA immunoreactivity in NBM (p<0.01 for all dose groups). In the hippocampus, cortex, and thalamus regions, there was also slight enhancement of TrkA immunoreactivity compared with the IBO group.
As shown in Fig. 3D and Fig. 4D, p75 expression of IBO rats was significantly reduced in the cortex as compared to the sham group (p<0.05), whereas BSYZ treatment significantly enhanced p75 expression compared with the IBO group (p<0.05, p<0.01, and p<0.01 for low-, middle-, and high-dose groups, respectively). In the NBM, p75 expression in IBO group was significantly increased as compared to the sham group (p<0.05), and reduction of p75 expression was observed in the BSYZ group compared with the IBO group. In the hippocampus and thalamus, there were no significant changes of p75 expression among groups.
Effects of BSYZ on NGF, TrkA, and p75 mRNA expression in the brain
The level of NGF mRNA was significantly decreased in IBO-induced rats compared to the sham group (p<0.05 and p<0.01 for hippocampus and cortex, respectively) (Fig. 5A, D). However, an increase of NGF mRNA expression was observed after treatment with BSYZ compared with the IBO group (hippocampus, p<0.05 for both low- and high-dose groups; cortex, p<0.05 for both low- and high-dose groups) (Fig. 5A, D), and there was also a slight increment in the cortex for the middle-dose group (Fig. 5D).

Effects of Bushen-Yizhi Formula (BSYZ) on nerve growth factor (NGF), TrkA, and p75 mRNA expression in the brain. (
TrkA mRNA was significantly reduced in the IBO-treated rats compared to the sham group (p<0.05 for both hippocampus and cortex) (Fig. 5B, E). After treatment with BSYZ, the level of TrkA mRNA was obviously increased especially in the cortex compared with the IBO group (p<0.01 and p<0.05 for the middle- and high-dose groups, respectively), and there was a slight improvement in the low-dose group (Fig. 5E).
In the IBO group, p75 mRNA was significantly down-regulated in the cortex compared to the sham group (p<0.01) (Fig. 5F), whereas no significant reduction was observed in the hippocampus (Fig. 5C). However, administration of BSYZ at high dose significantly up-regulated p75 mRNA expression compared with the IBO group (hippocampus, p<0.05; cortex, p<0.01) (Fig. 5C, F).
Effects of BSYZ on markers of cholinergic neurons in brain
ACh concentration, ChAT activity, and ChE activity in the cortex were significantly reduced by IBO injection as compared to the sham group (p<0.01, p<0.05, and p<0.01, respectively), whereas BSYZ treatment significantly enhanced ACh concentration (p<0.05 for the low-dose group) and ChE activity (p<0.05 for low- and high-dose groups) compared with the IBO group (Fig. 6A–C). The mRNA levels of ChAT and AChE in IBO group were significantly decreased in the cortex compared to the sham group (both p<0.05) (Fig. 6D, E). However, administration of BSYZ at middle and high doses significantly increased the mRNA levels of ChAT (p<0.01 and p<0.05, respectively) (Fig. 6D) and AChE (both p<0.05) (Fig. 6E) compared with the IBO group. In the hippocampus, a significant reduction of ChAT mRNA and AChE mRNA expression was observed in IBO group rats (both p<0.01 compared to the sham group), and BSYZ treatment ameliorated these changes (ChAT, p<0.05 and p<0.05 for low- and middle-dose groups, respectively; AChE, p<0.05 and p<0.01 for low- and middle-dose groups, respectively) (Fig. 6D, E). The protein levels of ChAT and AChE were significantly reduced by IBO injection when compared to the sham group, and BSYZ treatment significantly up-regulated ChAT protein expression (p<0.05, p<0.01, and p<0.05 for low-, middle-, and high-dose groups, respectively) and AChE protein expression (p<0.05 for all dose groups) compared with the IBO group (Fig. 6F, G).
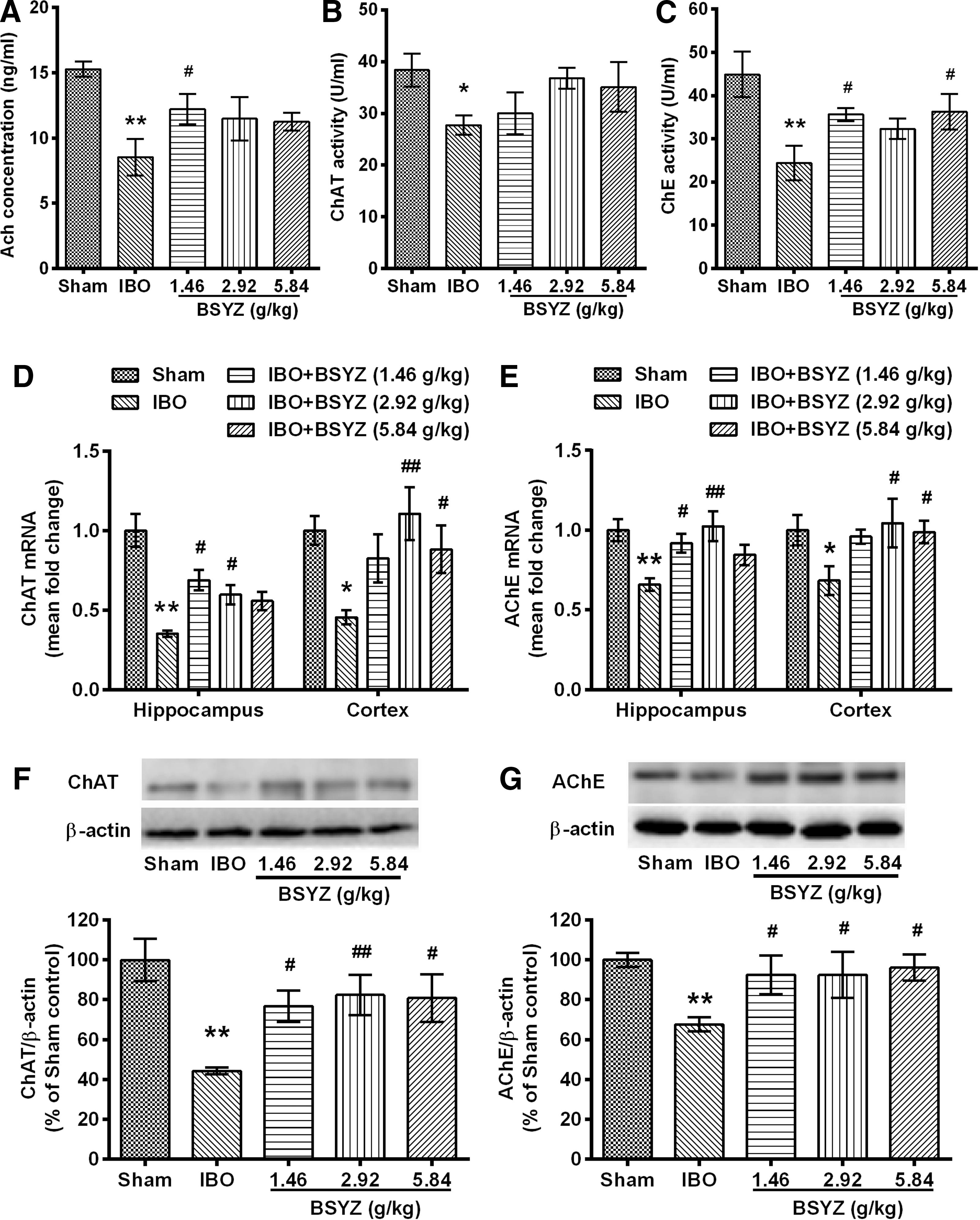
Effects of Bushen-Yizhi Formula (BSYZ) on acetylcholine (ACh), choline acetyltransferase (ChAT), and acetylcholinesterase (AChE) expressions in brain. (
Effects of BSYZ on neuronal apoptosis and histological morphology in brain
The apoptotic-related protein cleaved caspase-3 level in the IBO group was significantly higher than that in the sham group (p<0.01) (Fig. 7A), whereas BSYZ significantly down-regulated cleaved caspase-3 expression as compared with the IBO group (p<0.01 for all dose groups) (Fig. 7A). Figure 7, B and C, shows a similar neuronal apoptosis in TUNEL staining. The number of TUNEL-positive staining cells in the IBO group was significantly higher than that in the sham group (p<0.01). However, with the application of BSYZ, there were decreased numbers of TUNEL positive staining cells in both hippocampus and cortex (p<0.01, p<0.01, and p<0.05 for low-, middle-, and high-dose groups, respectively). Additionally, there were more TUNEL-positive staining cells in the CA1 region of the hippocampus and the cortex (Fig. 7C).
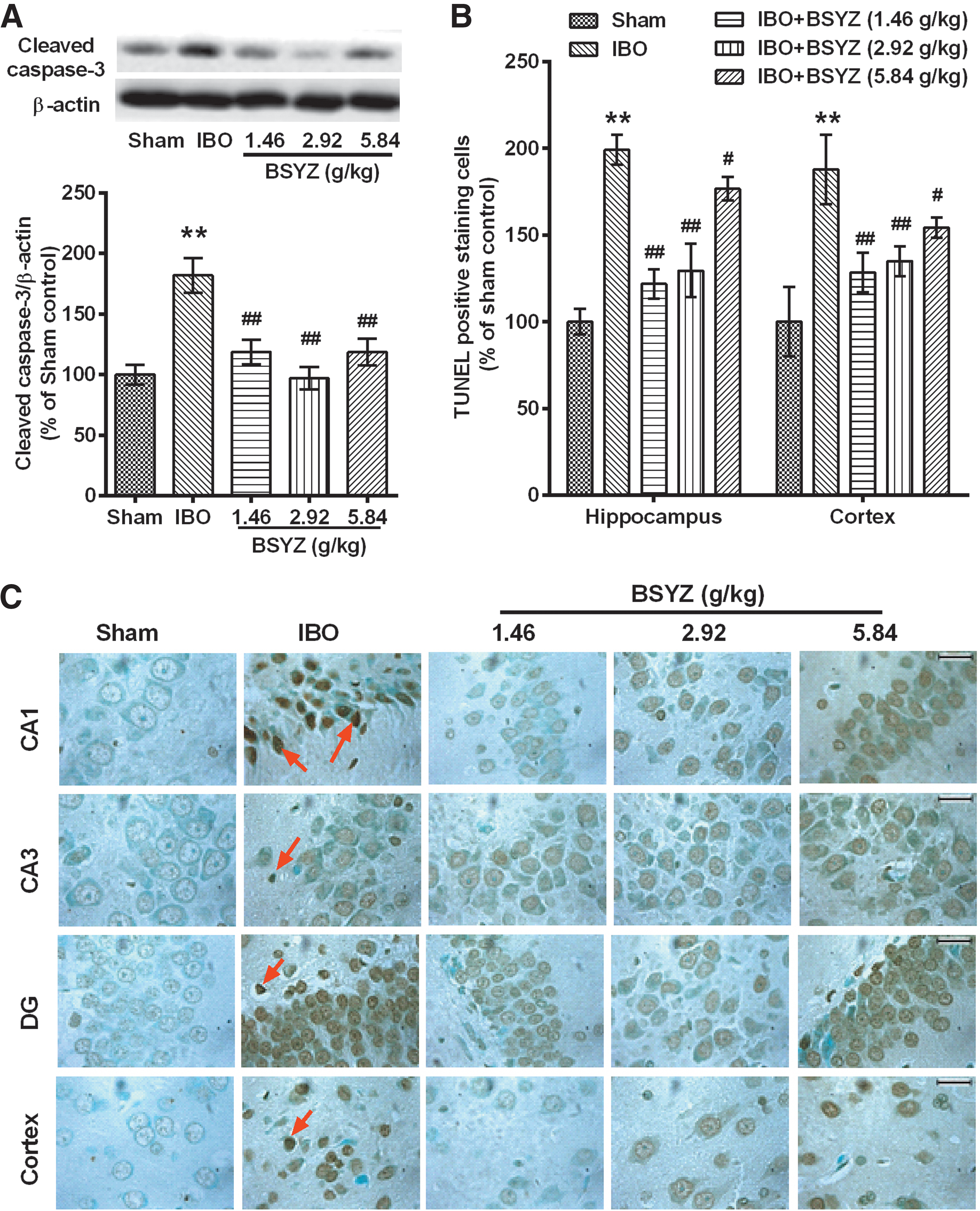
Effects of Bushen-Yizhi Formula (BSYZ) on neuronal apoptosis. (
The histological morphology examination showed irregular arrangements of neurons in the hippocampus, and unhealthy neurons (appeared as either indistinct, lacking a clear cell boundary, with a small darkened nucleus, or darkened, shrunken) (Fig. 8A), as well as chromatolysis and decrease of the Nissl body with the shrinkage of cell body (Fig. 8B) in IBO rats. In the CA1, CA3, and cortex regions, unhealthy neurons were significantly increased in IBO group (all p<0.01 compared to the sham group) (Fig. 9A), whereas Nissl-positive neurons were significantly less than those in the sham group (all p<0.01) (Fig. 9B). However, these abnormalities were markedly ameliorated after administration of BSYZ (Fig. 8), with a significant decrease of unhealthy neurons (CA1, p<0.01 for all dose groups; CA3, p<0.05 for all dose groups; cortex, p<0.01, p<0.01, and p<0.05 for low-, middle-, and high-dose groups, respectively) (Fig. 9A) and increase of Nissl-positive neurons compared with the IBO group (CA1, p<0.01 for all dose groups; CA3, p<0.05, p<0.01, and p<0.05 for low-, middle-, and high-dose groups, respectively; cortex, p<0.05 for both low- and middle-dose groups) (Fig. 9B).
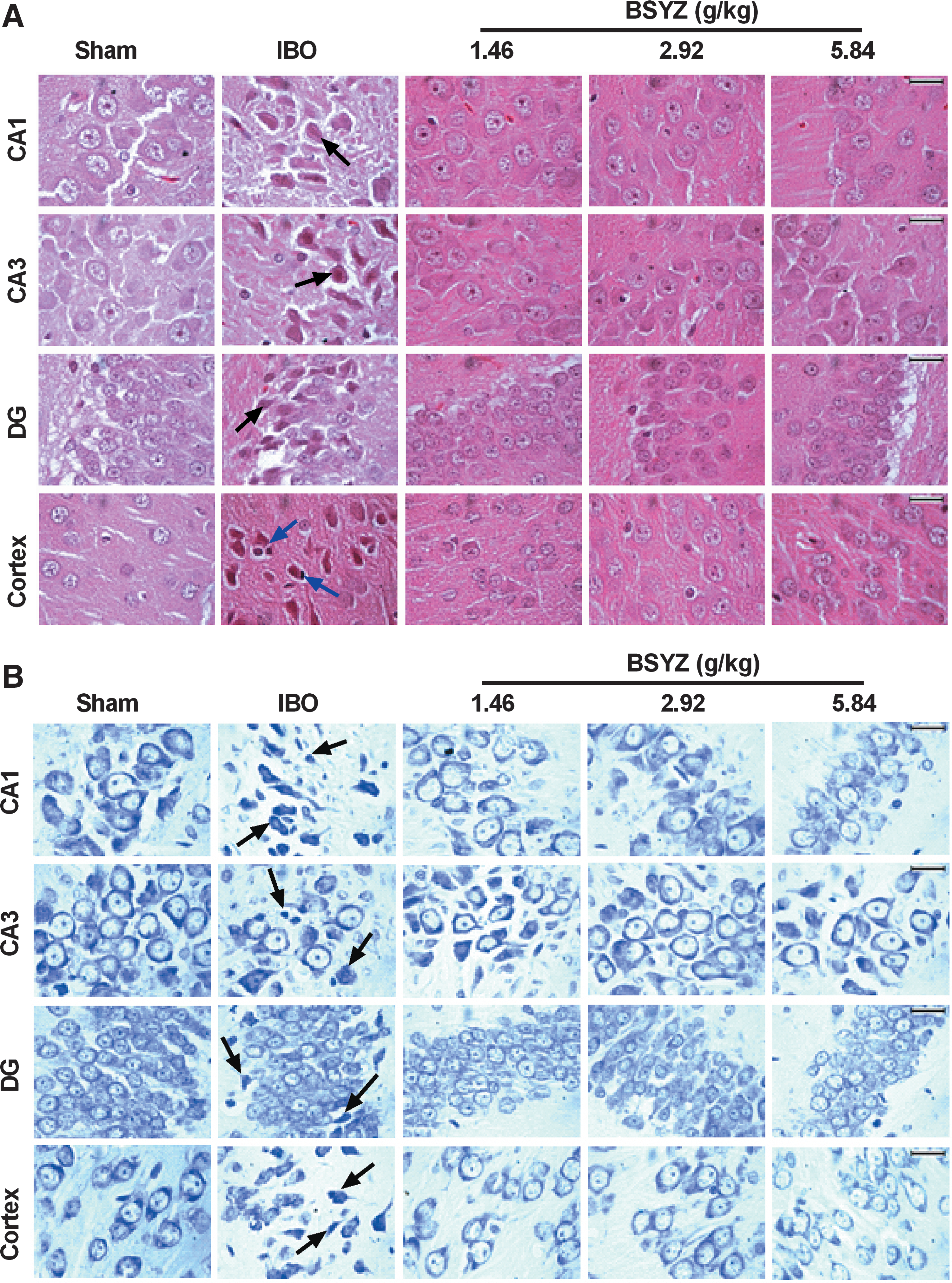
Effects of Bushen-Yizhi Formula (BSYZ) on the histological morphology in brain. (

Number of unhealthy neurons and Nissl-positive staining cells in brain. (
Discussion
The present study demonstrated that the BSYZ formula ameliorated learning and memory impairments, reversed cholinergic dysfunction, enhanced NGF expression, and inhibited neuronal apoptosis in the IBO-induced rats. Memory loss and cholinergic dysfunction are considered as two important hallmarks of many neurodegenerative and neuropsychiatric diseases. Many reports have shown cholinergic system atrophy in patients with AD or MCI 23 and a reduction of two key cholinergic markers, ChAT and AChE, in postmortem brain tissue from AD patients and MCI patients. 24,25 Thus, intracerebral injection of IBO served as a modeling method of AD, on the basis of its effects on cognitive and cholinergic changes. 16,26,27
In the present study, when determining the hippocampal-dependent spatial memory with the Morris water maze test, there was a significant difference between sham rats and IBO rats, indicating that spatial memory was impaired in IBO rats. With regard to object memory, which is both hippocampus and cortex dependent, 28 –30 there was also a significant difference between sham rats and IBO rats, suggesting that object memory was impaired in IBO rats. However, after oral administration of BSYZ for 4 weeks, escape latencies in the Morris water maze test were gradually decreased and significantly different from the IBO rats, and the number of crossing the platform was also increased. In addition, BSYZ-treated rats showed a significant preference for the novel object in the novel object recognition test. These results suggested that BSYZ could reverse the learning and memory impairments induced by IBO.
Aging-relative memory deficits could be reversed by several members of the neurotrophin family, 31 including NGF, 18 as well as by activation of multiple neurotrophin receptors (such as TrkA and p75) associated with cholinergic neurons in the brain. 31 It was shown that ChAT and TrkA expression were markedly reduced, whereas p75 was relatively high and independent of the NGF treatment in the basal forebrain cholinergic neurons of aged rats. 32 Defects in the production and/or utilization of the TrkA receptor might be a key event mediating degeneration of NGF-responsive cholinergic basal forebrain neurons in AD. 33 An in vitro study showed that NGF increased ACh transmission via p75-mediated mechanisms. 34 Furthermore, p75 might modulate cell survival and death decisions. 35 Although the mechanisms of NGF signaling are various and complex, evidence has shown that NGF might be a promising target for the treatment of dementia. A recent phase 1 clinical trial of ex vivo NGF gene delivery in mild AD has shown improvement in the rate of cognitive decline. 36
To explore whether the BSYZ formula has an effect on NGF signaling, we detected ChAT, NGF, TrkA, and p75 in the rat brain. In the hippocampus, ChAT, NGF, and TrkA were significantly reduced after IBO injection, whereas significant differences were not observed for p75. BSYZ treatment significantly enhanced expression of ChAT and NGF. In the cortex, ChAT, NGF, TrkA, and p75 were significantly decreased after IBO injection, and reverse influences were observed in expression of ChAT, NGF, and p75 with administration of BSYZ. In the NBM region, ChAT, NGF, and TrkA were significantly reduced after IBO injection, whereas an opposite change for p75 was observed. BSYZ treatment significantly enhanced immunoreactivity of NGF and TrkA. In the thalamus, NGF and TrkA were significantly reduced after IBO injection, whereas BSYZ failed to exhibit significant alternations of TrkA expression. These results demonstrated different changes of cholinergic phenotype, loss of which might induce senile impairment of cholinergic neurons 32 in different brain regions. It should be noted that the effects of BSYZ on NGF expression were similar to that on ChAT expression in the hippocampus and cortex. These two brain regions were involved in the process of recognition and memorization and played distinct roles. The hippocampus might play a central role in spatial memory, whereas the cortex might have a hippocampal-independent role in memory. 37 Additionally, it has been reported that NGF is related to the normal cholinergic function in animal brain, 38,39 and endogenous NGF regulates expression of TrkA and ChAT. 40 This observation also suggests that BSYZ enhances endogenous NGF and ChAT in the hippocampus and cortex, which might contribute to improving memory.
Because cholinergic deficits are a crucial characteristic of many neurodegenerative diseases, we explored the effects of BSYZ on cholinergic neurons and neuronal apoptosis in the hippocampus and cortex. Examination of cholinergic markers revealed a reduction of ACh level, ChAT activity, and ChE activity, as well as down-regulation of ChAT and AChE protein levels and mRNA expression in IBO-induced bilateral NBM lesions. Furthermore, detection of histological morphology and the apoptosis-relative protein cleaved caspase-3 showed not only neuronal loss and apoptosis, but also an increase of cleaved caspase-3 protein expression in IBO rats. However, administration of BSYZ significantly reversed these cholinergic dysfunctions as well as neuronal apoptosis. Additionally, the results of TUNEL staining revealed a dose-dependent decrease in anti-apoptosis. These results indicated that BSYZ had neuroprotective effects probably via cholinergic pathway.
In traditional Chinese medicine (TCM), the theory “the kidney nourishes marrow and brain is the sea of marrow,” which was recorded in the ancient book of The Medical Classic of the Yellow Emperor (Huang Di Nei Jing), has been instructing traditional Chinese medical practitioners in preventing and treating dementia in aged people for thousands of years. 41 Hence, a common therapy in treating dementia was “kidney-reinforcing” by using “kidney-reinforcing” herbs. Of the six herbs of BSYZ—She Chuang Zi, Zhi He Shou Wu, Nv Zhen Zi, and Gou Qi—all belong to the “kidney-reinforcing” herbs category. The combination of six herbs has effects of “kidney-reinforcing” and “brain nourishing.” On the other hand, modern pharmacological research has shown that six of the major constituents of BSYZ—ginsenoside Rg1, ginsenoside Rb1, tetrahydroxystilbene glucoside, osthole, paeoniflorin, and imperatorin—all had ameliorative effects on learning and memory deficits in animal models. 42 –47 Other studies in vitro and in vivo also exhibited neuroprotective effects of ginsenoside Rg1 48 and ginsenoside Rb1, 49 anti-apoptotic actions of osthole 50 and imperatorin, 51 and the ameliorating oxidative stress effect and regulating the NGF-mediated signaling effect of paeoniflorin. 52 Furthermore, research on the BSYZ formula has revealed effects of raising ChAT activity and neuroprotection in vitro. 14,15 The major constituents of BSYZ exerted multiple neuroprotective properties, which might underlie its efficacy in treating experimental cholinergic impairment in the present study. In addition, the synergistic effect of BSYZ compound prescription might influence the bioavailability of its constituents. As shown in a recent pharmacokinetic study, the bioavailability of osthole was dramatically enhanced after oral administration of BSYZ extract when compared with oral administration of pure osthole. 53
After BSYZ administration, the typical dose–response effect was not observed in the present study. Although significant improvements of object memory were observed in all BSYZ groups, it was noteworthy that compared to the other two groups, BSYZ at medium dose did not have a significant effect on ameliorating spatial memory and regulating NGF signaling in the hippocampus. This finding might be explained by the following. First, different brain regions might have distinct contributions to memory. It has been reported that the hippocampus might play a central role in spatial memory, whereas the cortex might have a hippocampal-independent role in memory. 37 Perirhinal cortex was considered as a critical region for object recognition memory. 54 The spatial memory impairment in the present study might be due partly to the disorder of NGF signaling in the hippocampus. It might be possible that BSYZ at the medium dose failed to regulate NGF signaling and thereby could not effectively improve the memory deficit. Second, the Chinese herbal formulas usually target multiple targets, pathways, and systems. The multiple effects of BSYZ were potentially implemented via multiple targets, which might be different from that of a single-target agent. Elucidation of the more detailed mechanisms merits further studies.
In conclusion, the present study indicated that the BSYZ formula was effective in ameliorating the spatial memory and object memory impairments in IBO-induced rats, and its neuroprotective effects might be mediated, at least in part, through cholinergic pathways, NGF signaling, and anti-apoptosis. Hence, our study suggests that application of the BSYZ formula may be a potential therapeutic approach in the treatment of AD and other neurodegenerative diseases that manifest cholinergic symptoms.
Footnotes
Acknowledgments
This work was supported by the National Natural Science Foundation of China (no. 81273817), Doctoral Fund of Education Ministry of China (no. 20124425120016 and no. 20134425110003), Guangdong Provincial Major Science and Technology for Special Program of China (no. 2012A080202017), Guangdong Provincial Natural Science Foundation of China (no. S2012040006514), the Scientific and Technical innovation Project of Guangdong Provincial Education Department of China (no. 2012KJCX0032), and the Characteristic Key Discipline Construction Fund of Chinese Internal Medicine of Guangzhou University of Chinese Medicine.
Author Disclosure Statement
No competing financial interests exist.