
Abstract
The objective of this meta-analysis was to determine the association of the apolipoprotein E (ApoE) gene with exceptional longevity (EL) (i.e., reaching 100+ years) by identifying possible unequal distribution of alleles/genotypes in the common variants ε2, ε3, and ε4 among centenarians and younger population. The association of ApoE with EL was analyzed in a total of 2776 centenarians (cases) and 11,941 younger controls (from 13 case–control studies) using the chi-squared test with the Yates correction. We conducted combined and separate analyses for all ethnic groups studied in the literature (Caucasian and Asian). The main result for all ethnic groups combined was that the likelihood of reaching EL was negatively associated with ε4 allele carriage [pooled odds ratio (OR)=0.43; 95% confidence interval (CI) 0.36, 0.50; p<0.001] and with ε4/ε4 (OR=0.18; 95% CI 0.08, 0.39; p<0.001), ε3/ε4 (OR=0.44; 95% CI 0.37, 0.53; p<0.001) and ε2/ε4 genotypes (OR=0.48; 95% CI 0.31, 0.74; p<0.001). In contrast, the ε2/ε3 genotype was positively associated with EL (OR=1.35; 95% CI 1.06, 1.72; p=0.017). When compared with the ε3 allele, the ε2 allele was not associated with increased odds of EL (OR=1.08; 95% CI 0.77, 1.50, p=0.660). The present meta-analysis confirms that, besides its previously documented influence on Alzheimer's and cardiovascular disease risk, the ApoE gene is associated with the likelihood of reaching EL.
Introduction
T
Whether ApoE is associated with exceptional longevity (EL), i.e., reaching 100+ years of age, however, is not clearly established. To identify those genetic variables that are associated with EL is of interest because centenarians, more so than nonagenarians, are the survival tail of the population and also a model of healthy aging. They have avoided or postponed age-related diseases and their fatal consequences 8 and usually delayed the onset of disability until their mid-90s. 9,10
Thus, a possible approach for identifying the gene variants that are associated with EL is to compare the genotypes of centenarians and ethnically matched younger controls. 11 This approach relies on the hypothesis that centenarians have genetic variants that influence human life span compared with the general population, which rarely reaches this age. 12 A lower frequency of the “risk” ApoE ε4 allele was reported among centenarians compared with their ethnically matched younger controls in Spanish, 13 French, 14,15 Finnish, 16 southern Italian, 17 northern Italian, 13 Chinese, 18 or Japanese cohorts. 13 Yet others found no such association in Japanese, 19 Finnish, 20 or southern Italian cohorts. 21 In turn, the ApoE ε2 allele was more frequent in some cohorts of centenarians compared to their younger controls, 14 –16,22 but this finding was not replicated by others. 17 –20 A previous sub-meta-analysis of studies published until 2003 reported a point estimate when comparing centenarians versus younger controls of 1.50 [95% confidence interval (CI) 1.27, 1.78] for the ε2 allele and of 0.49 (95% CI 0.41, 0.58) for the ε4 allele frequency. Here, we conducted a meta-analysis of all the available case–control studies (as of February 7, 2014) reporting the potential association between the ApoE gene and human EL.
Methods
Literature search
A search of electronic databases was conducted to identify all available publications on ApoE and human EL (as of February 7, 2014). The search terms “ApoE” or “apolipoprotein E,” “longevity” or “centenarian,” and “polymorphism” or “variation” or “genotype” were used, or combinations of one or more of these terms, with restrictions to English-language articles, in Medline, EMBASE, and Web of Science databases. We also extended the search spectrum to the “related articles” and the references of all retrieved studies. The authors of published papers were also contacted directly if crucial data were not reported in original papers. Once potentially relevant articles were identified, their references and abstracts were retrieved. A total of 126 papers were identified, and from their abstract or title it was determined that 113 were not relevant to the analysis or were excluded with reasons (Fig. 1).

Literature search flow.
Inclusion and exclusion criteria
The following criteria were used to include studies in the meta-analysis: (1) providing information on the evaluation of the common ApoE alleles ε2, ε3, and ε4 and on the “EL” phenotype; (2) using a case (centenarians)–control (younger adults) design; and (3) providing sufficient genotype data for calculating the odds ratio (OR) with 95% CI. Reasons for exclusion of studies were: (1) Mean age of the cases' group <100 years; (2) lack of control group; (3) overlapping study populations; (4) reported data not being usable; and (5) not written in English. A total of 126 studies were considered to be relevant, but 108 were excluded because the mean age of cases was not ≥100 years (Fig. 1). From all remaining eligible papers (n=16), only 13 met all inclusion criteria.
Data extraction
We followed the recommendations for Meta-analyses of Observational Studies in Epidemiology (MOOSE). 23 Two authors (A.S.L. and P.J.M.) independently viewed the titles and abstracts of all the studies identified in the electronic searches. The full copies of all possibly or definitely relevant studies were obtained and independently inspected by these two authors to determine whether they met the inclusion criteria. When a difference in opinion occurred, a third reviewer (N.G.) was consulted as an arbiter. The above-mentioned two authors independently summarized the following information from all eligible studies: The last name of the first author, publication year, ethnicity of the population studied, number of subjects in each category (cases and controls), baseline characteristics of the study population, and the counts of subjects with different genotypes in cases and controls. Information on Hardy–Weinberg equilibrium (HWE) and allele frequency was also tracked or calculated manually if not reported explicitly. Finally, any discrepancies were adjudicated by a discussion and a consensus was reached.
Statistical methods
Statistical analysis was performed using the chi-squared test with the Yates correction. Pooled OR (95% CI) was estimated using a weighted random-effect model. The OR can be used to determine whether a particular allele or genotype is associated with an outcome (e.g., EL), and to determine the magnitude of this association; that is, OR=1 indicates an allele or genotype does not affect odds of outcome; OR >1 indicates an allele or genotype is associated with higher odds of outcome; and OR <1 indicates a particular allele or genotype is associated with lower odds of outcome. 24 The significance limit was defined as p<0.05. The Egger test was used to evaluate possible publication bias in the meta-analysis with the number of included studies >2. 25 Heterogeneity, which partly determines the difficulty in drawing overall conclusions, was examined by Q and I2 statistics. The Q test only informs us about the presence or the absence of heterogeneity, but it does not report on the extent of such heterogeneity. The Q test was computed by summing the squared deviations of each study's effect estimate from the overall effect estimate, weighting the contribution of each study by its inverse variance. The I2 statistic describes the percentage of the total variability in a set of effect sizes due to true heterogeneity, that is, to between-studies variability. Thus, percentages of around 25%, 50%, and 75% would mean low, medium, and high heterogeneity among studies, respectively. 26 All statistical analyses were performed using MIX Pro software version 2.0. 27
We analyzed each individual genotype against the “reference” ε3/ε3 genotype. We also analyzed the association between the different alleles with EL, i.e., analyzing each of ε2 and ε4 versus the “reference” ε3 allele. The above-mentioned analyses were first performed for all subjects included in the retrieved studies (irrespective of their ethnic origin), and thereafter for Caucasian or Asian subjects only.
Results
Study characteristics
A flow diagram of the literature search is shown in Fig. 1. According to the above-defined inclusion criteria, we included 13 independent case–control studies in the meta-analysis, adding a total of 2776 cases and 11,941 controls. 13 –17,19,21,28 –33 The ethnicities included were Caucasian (n=8 studies) and Asian (n=4 studies); two cohorts of Caucasians (from northern Italy and Spain) and one of Asian people were included in one study. 13 The main characteristics for all eligible studies are listed in Table 1.
Data from Aalto-Setala et al. 54
SD, standard deviation; HWE, Hardy–Weinberg equilibrium; NR, not reported; NA, not available.
Main meta results: Genotypes
ε3/ε3 versus ε2/ε2
The association of the ε2/ε2 genotype with EL was investigated in eight studies. 13 –15,28,29,31 –33 The frequency of the ε2/ε2 genotype did not differ between cases and controls (OR=1.65; 95% CI 0.75, 3.64; p=0.216), with no evidence of publication bias (p=0.206) or heterogeneity among studies (I2=0.81%, Q=8.065, p=0.427). This lack of association was confirmed when analyzing Caucasians only, 13 –15,28,29,32 i.e., OR=1.13 (95% CI 0.36, 3.56; p=0.835), with no evidence of publication bias (p=0.092) or heterogeneity (I2=31.50%, Q=7.298, p=0.199) (Fig. 2).
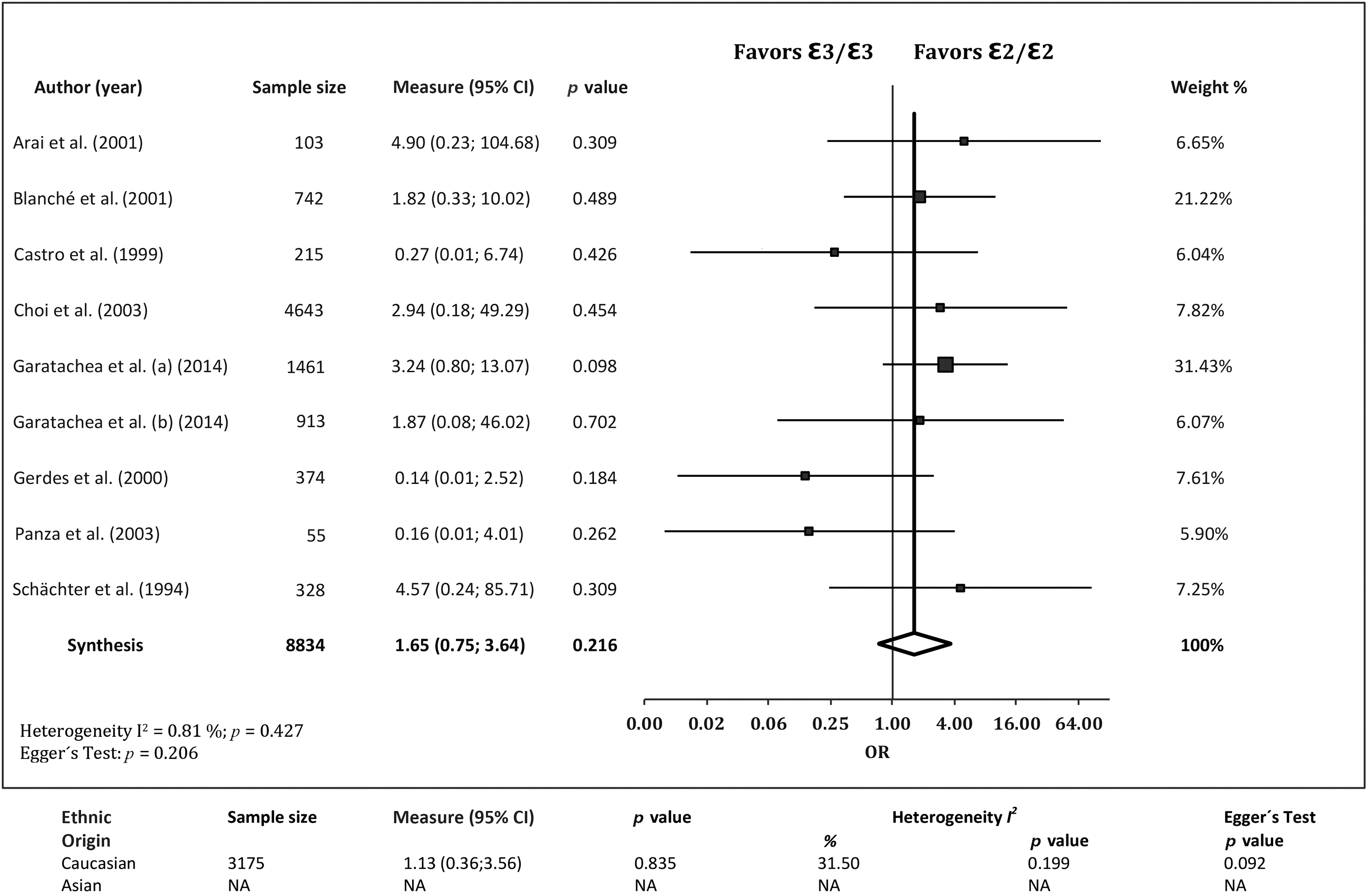
Associations between the ApoE genotype ε2/ε2 and exceptional longevity from the literature. ε3/ε3 was set as the reference group. CI, confidence interval; NA, not available; OR, odds ratio.
ε3/ε3 versus ε2/ε3
The association of the ε2/ε3 genotype with EL was investigated in nine studies. 13 –15,17,28,29,31 –33 The ε2/ε3 genotype was more frequent in cases than in controls with a pooled OR of 1.35 (95% CI 1.06, 1.72; p=0.017) and no evidence of publication bias (p=0.163) or heterogeneity among studies (I2=42.46%, Q=15.669, p=0.074). However, when confining the analysis to Caucasian cohorts, 13 –15,17,28,29,32 the ε2/ε3 genotype was not different between cases and controls, with a pooled OR of 1.31 (95% CI 0.95, 1.82; p=0.102) and no evidence of publication bias (p=0.322), but with heterogeneity among studies (I2=58.21%, Q=14.356, p=0.025) (Fig. 3).

Associations between the ApoE genotype ε2/ε3 and exceptional longevity from the literature. ε3/ε3 was set as reference group. CI, confidence interval; NA, not available; OR, odds ratio.
ε3/ε3 versus ε2/ε4
The association of the ε2/ε4 genotype with EL was investigated in seven studies. 13 –15,28,29,32,33 The ε2/ε4 genotype was less frequent in cases than in controls, with a pooled OR of 0.48 (95% CI 0.31, 0.74; p=0.001) and no evidence of publication bias (p=0.077) or heterogeneity among studies (I2=0.00%, Q=5.609, p=0.585). When this association was investigated for Caucasians only, 13 –15,28,29,32 it was corroborated that the ε2/ε4 genotype was less frequent in cases than in controls, i.e., OR=0.41 (95% CI 0.23, 0.74, p=0.003) and there was evidence of publication bias (p=0.014) but no heterogeneity among studies (I2=4.92%, Q=5.258, p=0.385) (Fig. 4).

Associations between the ApoE genotype ε2/ε4 and exceptional longevity from the literature. ε3/ε3 was set as the reference group. CI, confidence interval; NA, not available; OR, odds ratio.
ε3/ε3 versus ε3/ε4
The association of the ε3/ε4 genotype with EL was investigated in nine studies. 13 –15,17,28,29,31 –33 The ε3/ε4 genotype was less frequent in cases than in controls, with a pooled OR of 0.44 (95% CI 0.37, 0.53; p<0.001) and no evidence of publication bias (p=0.726) or heterogeneity (I2=0.00%, Q=6.386, p=0.700). When studying this association in Caucasian cohorts only, 13 –15,17,28,29,32 the ε3/ε4 genotype was also found to be less frequent in cases than in controls, with a pooled OR of 0.46 (95% CI 0.38, 0.57; p<0.001) and no evidence of publication bias (p=0.557) or heterogeneity among studies (I2=0.00%, Q=3.221, p=0.780) (Fig. 5).
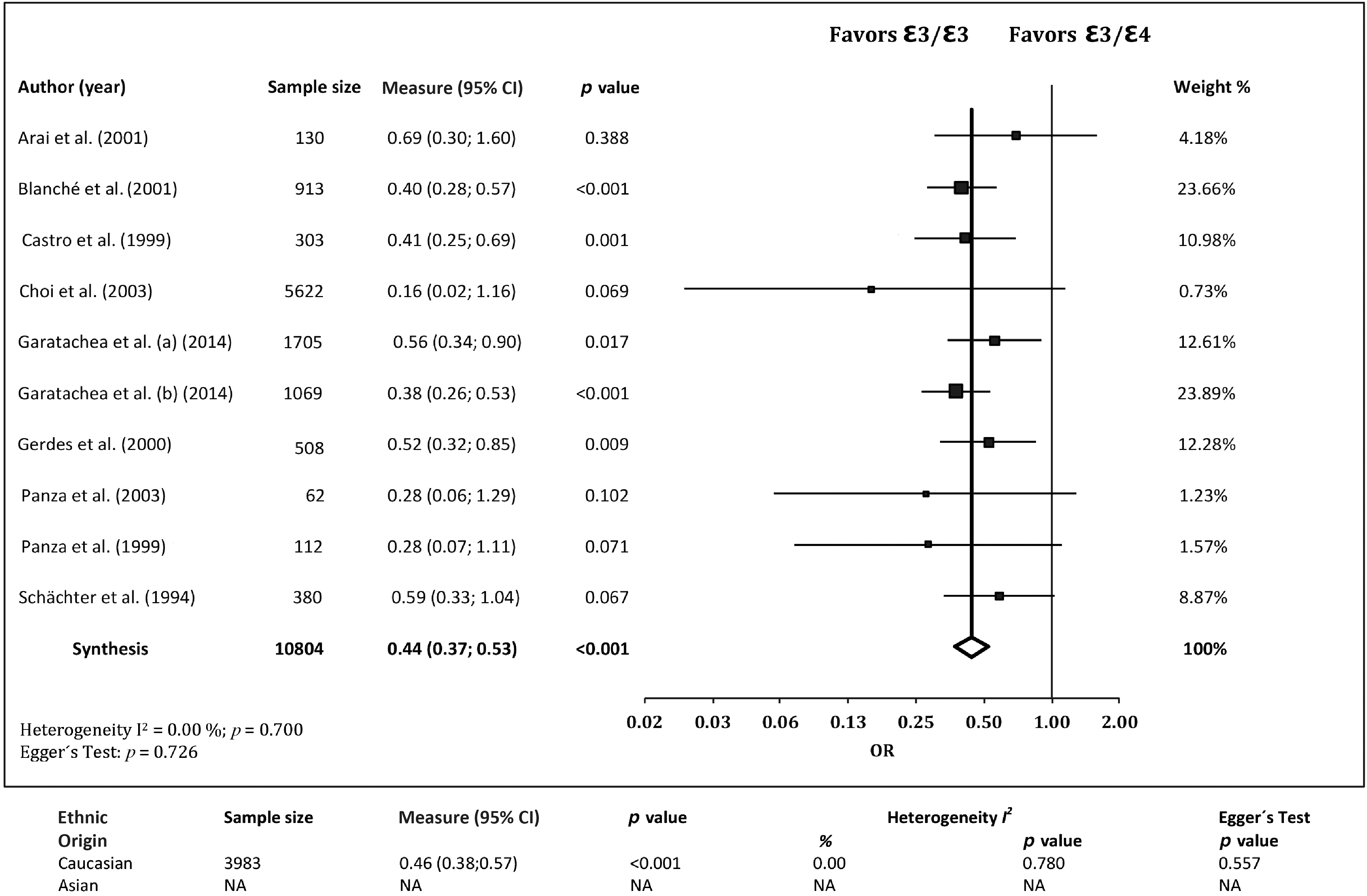
Associations between the ApoE genotype ε3/ε4 and exceptional longevity from the literature. ε3/ε3 was set as the reference group. CI, confidence interval; NA, not available; OR, odds ratio.
ε3/ε3 versus ε4/ε4
The association of the ε4/ε4genotype with EL was investigated in eight studies. 13 –15,17,28,29,32,33 The ε4/ε4 genotype was less frequent in cases than in controls, with a pooled OR of 0.18 (95% CI 0.08, 0.39; p<0.001). No evidence of publication bias (p=0.782) or heterogeneity was found (I2=0.00%, Q=4.206, p=0.838). This association was confirmed when confining the analysis to Caucasian cohorts, 13 –15,17,28,29,32 where the ε4/ε4 genotype was also found to be less frequent in cases than in controls, i.e., OR=0.12 (95% CI 0.04, 0.34; p<0.001) and no evidence of publication bias (p=0.985) or heterogeneity among studies (I2=0.00%, Q=1.330, p=0.969) (Fig. 6).
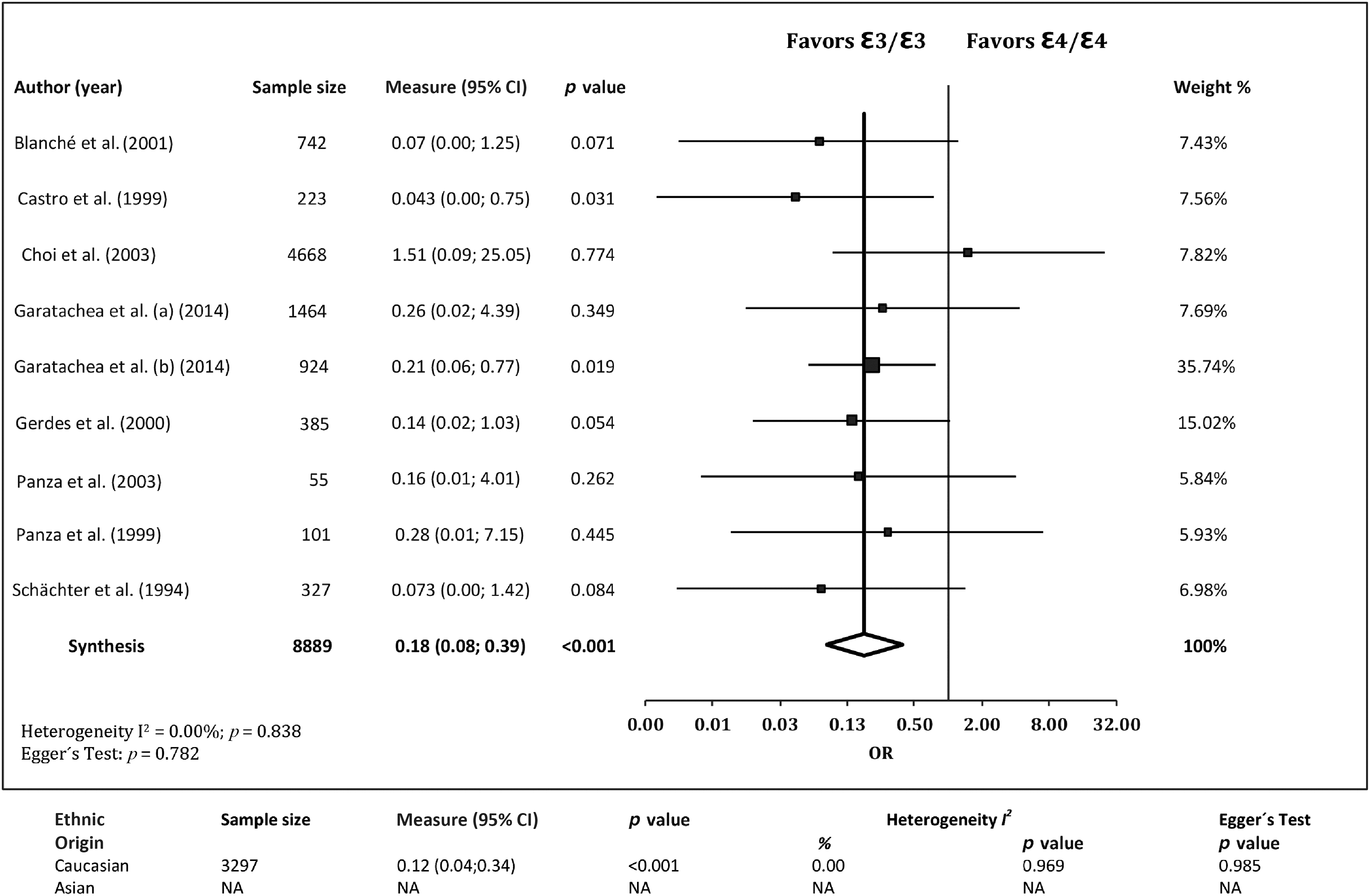
Associations between the ApoE genotype ε4/ε4 and exceptional longevity from the literature. ε3/ε3 was set as the reference group. CI, confidence interval; NA, not available; OR, odds ratio.
Main meta results: ε4 carriage versus no ε4 carriage
The association of ε4 carriage with EL was investigated in nine studies. 13 –15,17,28,29,31 –33 The frequency of ε4 carriers was lower in cases than in controls, with a pooled OR of 0.43 (95% CI 0.36, 0.50; p<0.001) and no evidence of publication bias (p=0.488) or heterogeneity (I2=0.00%, Q=5.465, p=0.738). In Caucasian cohorts, 13 –15,17,28,29,32 the frequency of ε4 carriers was also lower in cases than in controls, with a pooled OR of 0.41 (95% CI 0.34, 0.50; p<0.001) and no evidence of publication bias (p=0.556) or heterogeneity among studies (I2=0.00%, Q=4.382, p=0.694) (Fig. 7).
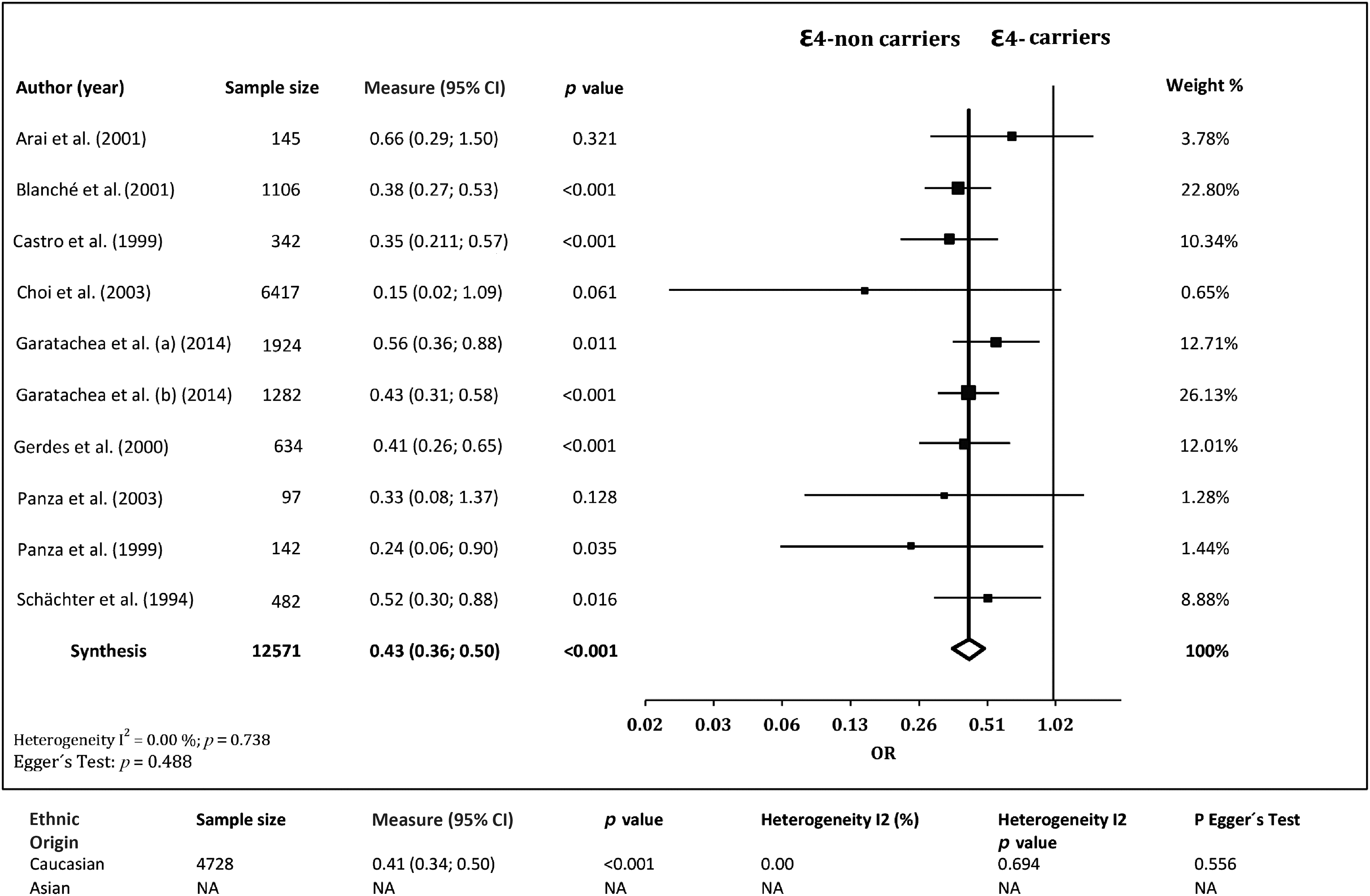
Associations between ApoE ε4 allele carriage and exceptional longevity from the literature. CI, confidence interval; NA, not available; OR, odds ratio.
Main meta results: Alleles
ε3 versus ε2 allele
The association of the ε2 allele (vs. the ε3 allele) with EL was investigated in 13 studies. 13 –17,19,21,28 –33 The frequency of the ε2 allele did not differ between cases and controls (OR=1.08, 95% CI 0.77, 1.50; p=0.660) and there was no evidence of publication bias (p=0.396) although there was heterogeneity among studies (I2=57.33%, Q=28.289, p=0.004). This lack of association was confirmed when analyzing: (1) Only Caucasians, 13 –17,21,28,29,32 i.e., OR=1.11 (95% CI 0.74, 1.68; p=0.586), with no evidence of publication bias (p=0.310) but with heterogeneity existing among studies (I2=70.24%, Q=27.230, p=0.001); or (2) only Asian cohorts, 13,19,30,31,33 i.e., OR=0.83 (95% CI 0.50, 1.37; p=0.527), with no evidence of publication bias (p=0.370) or heterogeneity (I2=0.00%, Q=1.909, p=0.770).
ε3 versus ε4 allele
The association of the ε3 versus ε4 allele with EL was investigated in 13 studies. 13 –17,19,21,28 –33 The frequency of ε4 allele did not differ between cases and controls (OR=0.64; 95% CI 0.40, 1.02; p=0.059) and there was no evidence of publication bias (p=0.441), although there was heterogeneity among studies (I2=77.78%, Q=53.675, p<0.001). This lack of association was confirmed when the analyses were confined to: (1) Caucasian cohorts, 13 –17,21,28,29,32 i.e., OR=0.61 (95% CI 0.33, 1.13; p=0.118), with no evidence of publication bias (p=0.786) but with heterogeneity existing among studies (I2=77.58%, Q=35.685, p<0.001); or (2) the Asian cohorts, 13,19,30,31,33 i.e., OR=0.67 (95% CI 0.26, 1.77; p=0.429), with no evidence of publication bias (p=0.387) but with heterogeneity (I2=71.73%, Q=14.147, p<0.001).
Discussion
The present meta-analysis shows a strong negative association of the likelihood of reaching EL with the homozygous ApoE ε4/ε4 genotype, the heterozygous ε3/ε4, and ε2/ε4 genotypes, and with ε4 allele carriage in general. Thus, the same allele, ε4, that increases AD and CVD risk, 4,34 could negatively affect EL. In contrast, except for the ε2/ε3 genotype, which was more frequent in centenarians, another important result was that overall carriage of the ε2 allele does not seem to be associated with EL. An additional novel finding of our study was that the ε3/ε3 genotype, that is, carriage of two copies of the most common ApoE allele in all populations, could positively affect EL in Caucasian populations. We believe our findings are of potential medical relevance because western societies are aging, with centenarians being the fastest growing demographic group of the world population (US National Institute on Aging 2007). This indicates that research should be conducted on those factors, including genetics that can be potentially associated with EL and healthy ageing.
Some debate exists in the field, ApoE being viewed by some authors as a “frailty” or “disease-risk” gene rather than as a longevity gene. 32,35,36 As such, the potential influence of this gene on death risk would theoretically decrease at very old ages. 35 This is in line with the “heterogeneity hypothesis,” that is, surviving carriers of a “frail” genotype might also possess other factors compensating for such a “frail” genotype at old ages. 35 However, our data seem more in agreement with an alternative viewpoint, the so-called “multifactorial threshold model” hypothesis, according to which it is the accumulation of many single factors, even with a relatively small individual effect (e.g., carriage of a frail genotype such as one or more ε4 alleles) that contributes to a condition (included death) to occur as we age. 37
There is a solid biological rationale in postulating that the gene encoding a major cholesterol carrier, ApoE, is a candidate to influence EL (see below for a summary of putative underlying biological mechanisms). The ε4 allele is a risk factor for age-related conditions, such as CVD and AD. A meta-analysis including 121 studies showed that the ε4 allele is associated with higher CVD risk and higher plasma cholesterol levels. 38 The well-documented impact of ApoE–ε4 on CVD risk is commonly attributed to its hypercholesterolemic effect, 39 and the impact of the different ApoE isoforms on blood concentrations of lipids and lipoproteins has been explained by isoform-related differences in several mechanisms, including lipoprotein receptor-binding affinity, dietary fat clearance, clearance of low-density lipoprotein (LDL) ApoB, and efficiency of intestinal cholesterol absorption. 40,41 With regard to AD, a recent meta-analysis of 21 studies showed that the ε4 allele is strongly associated with a higher risk of sporadic late-onset AD. 4 ApoE lipoproteins bind to several cell-surface receptors to deliver lipids and also to hydrophobic amyloid-beta (Aβ) peptide, with the latter initiating toxic events resulting in neurodegeneration in AD. 3 ApoE isoforms differentially regulate brain aggregation and clearance of Aβ and regulate brain lipid transport, glucose metabolism, neuronal signaling, neuroinflammation, and mitochondrial function. 3 Thus, the ε4 allele is associated with higher Aβ cellular uptake compared to ε3, thereby leading to more Aβ aggregation and higher AD risk. 3,42,43 Nevertheless, a causal relationship between the ε4 allele and higher AD risk does not necessarily exist.
Incidental findings often occur when genome sequencing data are screened for disease-causing variants, as Freudenberg-Hua et al. 44 recently noted. They reported one of 44 centenarians who did not have AD despite harboring the theoretically “highest-risk” genotype, i.e., ApoE ε4/ε4. On the other hand, except for the ε2/ε3 genotype, we found no evidence of a potentially favorable role of the ε2 allele on EL. In this regard, the ε2 allele might play a protective role against CVD, 32,38,45 with recent studies showing an association (thorough meta-analysis of 3189 nonagenarians cases and 5757 controls from Europe) between this allele and surviving to old age, 6 or that, among AD patients, levels of Aβ oligomers were lowest in ε2 allele carriers. 46 Yet our data are in agreement with those of previous large population-based longitudinal studies, i.e., the Rotterdam study 47 and the Danish 1905 birth cohort. 48 These two studies reported no favorable effect of the ε2 allele on mortality in the oldest old, despite protecting against aging cognitive decline. 48
Our data are in agreement with a previous sub-meta-analysis 49 showing lower ε4 allele frequency in centenarians compared with younger controls (OR=0.49; 95% CI, 0.41, 0.58). Yet a major novel aspect of our meta-analysis compared to the report by Lewis and Brunner 49 is that we analyzed not only allele but also genotype frequencies. Furthermore, given the large differences in relative ApoE allele frequencies that are known to exist according to ethnic origin, 49,50 we conducted not only pooled allele-frequency analyses, but also separate analyses for the two ethnic groups (Caucasian and Asian) that have been studied to date. (Separate analyses of genotype frequencies could not be performed in Asian cohorts owing to lack of available published data.) On the other hand, case–control studies comparing allele/genotype frequencies among centenarians and younger counterparts as the ones included in the present meta-analysis have some limitations that might explain controversy in the field.
First, candidate gene association studies using a case–control design as the ones analyzed are frequently underpowered due to small sample sizes (i.e., <100 subjects/group). 51 This is understandable owing to the difficulty of gathering centenarians, with the size of the cases' group being<100 subjects in seven of the 13 studies we analyzed (Table 1). Indeed, living 100 or more years is still a rare phenotype, i.e., less than one of every 10,000 people. 52 Thus, meta-analyses of multiple studies as the present one are needed to overcome the sample size limitation. The theoretically higher mortality risk conferred by a given genotype might be confounded by the interaction with environmental or lifestyle factors (which are seldom accounted for in candidate gene case–control studies). 49 With regard to the latter, EL is likely influenced by complex genotype–environment interactions. As such, differences in environmental (lifestyle, geographic) factors among cohorts might explain differences in the potential association between EL and a given gene (e.g., ApoE) among studies. On the other hand, it is likely that the combined effect of different genotypes influences EL beyond the potential individual effect of a single gene effect. In this regard, Sebastiani et al. recently reported a multivariate modeling of EL showing different allele combinations of 281 single-nucleotide polymorphisms (SNPs) that determine different probabilities of survival to≥100 years. 53 One of the 281 SNPs that reached irrefutable genome-wide significance was rs2075650 in the translocase of the outer mitochondrial membrane 40 homolog (TOMM40)/APOE, which occurs in an intronic TOMM40 variant but is also a strong proxy of the SNPs that define the ApoE alleles. Finally, interpretation of case–control studies comparing allele/genotype frequencies among centenarians and younger counterparts may be flawed by differences in date of birth (which it is unknown for the studies included in this meta-analysis), because the pattern of exposure to environmental/lifestyle risk factors can vary with year of birth. 49
Conclusions
In summary, our meta-analysis of results from the published literature provides evidence mainly for a negative association between ApoE ε4 allele carriage and human EL. Although the biological mechanisms explaining such findings remain to be clearly elucidated, this finding is of potentially relevance for predicting and identifying individual differences during the aging process.
Footnotes
Author Disclosure Statement
No competing financial interests exist.