
Abstract
The lack of evolutionary established mechanisms linking genes to age-related traits makes the problem of genetic susceptibility to health span inherently complex. One complicating factor is genetic trade-off. Here we focused on long-living participants of the Long Life Family Study (LLFS), their offspring, and spouses to: (1) Elucidate whether trade-offs in the effect of the apolipoprotein E e4 allele documented in the Framingham Heart Study (FHS) are a more general phenomenon, and (2) explore potential mechanisms generating age- and gender-specific trade-offs in the effect of the e4 allele on cancer, diseases of the heart, and neurodegenerative disorders assessed retrospectively in the LLFS populations. The e4 allele can diminish risks of cancer and diseases of the heart and confer risks of diseases of the heart in a sex-, age-, and LLFS-population-specific manner. A protective effect against cancer is seen in older long-living men and, potentially, their sons (>75 years, relative risk [RR]>75=0.48, p=0.086), which resembles our findings in the FHS. The protective effect against diseases of the heart is limited to long-living older men (RR>76=0.50, p=0.016), as well. A detrimental effect against diseases of the heart is characteristic for a normal LLFS population of male spouses and is specific for myocardial infarction (RR=3.07, p=2.1×10−3). These trade-offs are likely associated with two inherently different mechanisms, including disease-specific (detrimental; characteristic for a normal male population) and systemic, aging-related (protective; characteristic for older long-living men) mechanisms. The e4 allele confers risks of neurological disorders in men and women (RR=1.98, p=0.046). The results highlight the complex role of the e4 allele in genetic susceptibility to health span.
Introduction
R
One fundamental mechanism, which is the most studied to date, is associated with the biochemical genetic basis of a specific trait. 5 Another mechanism, which is substantially less studied, is associated with the systemic decline in the functioning of an organism with aging. 5,6 A substantial basis of the systemic mechanism comes from observations of changes in the expressions of various phenotypes, regardless of their specifics with age (e.g., levels of physiological markers, 7 –9 bone mineral density, 10 or risks of aging-related diseases 11 ). Accordingly, this systemic mechanism can be associated with aging and, thus, explain the risks of not just one, but perhaps a major portion of traits in late life. 5,6 An exciting idea is that the discovery of genes associated with this systemic mechanism could be a major breakthrough in the problem of the genetic regulation of health span. 1,12,13
Landmark properties of the systemic, aging-related biogenetic mechanism of health span are its broad, inherently pleiotropic nature 14,15 and its sensitivity to age. Pleiotropy in the effects of genes on traits in late life is becoming recognized increasingly. 14 Studies also provide examples of its complex forms, such as genetic trade-offs 16 –22 and antagonistic pleiotropy (see ref. 23; examples are in refs. 24 –29). Trade-off is a broader concept than antagonistic pleiotropy because it refers to antagonistic effects of the same allele on different phenotypes that may not necessarily include fitness traits. Studies also provide evidence of age-sensitive genetic effects, i.e., that the same alleles could confer different risks of the same traits at different ages. 24,30 –36
In this study, we used information on 4659 genotyped participants of the Long Life Family Study (LLFS) to address two questions. First, we investigated whether recently documented trade-offs in the effects of the apolipoprotein E (APOE) common polymorphism on major human diseases in the Framingham Heart Study (FHS) 18,34 are a more general phenomenon extendable to other populations. Second, we explore mechanisms that can generate age- and gender-specific trade-offs in the effect of the APOE e4 allele on major human diseases, including cancer, diseases of the heart, and neurodegenerative disorders.
Methods
Data
The LLFS collected data at four field centers (three in the United States and one in Denmark) on families showing exceptional familial longevity. The study eligibility criteria are described in detail elsewhere. 37 –39 Briefly, in the United States, the families eligible for the LLFS must have two living siblings aged 80+ years, two living offspring of one or more of the siblings, and a living spouse of one of the offspring; the offspring represent a normal population. In addition, the family must demonstrate exceptional longevity on the basis of a Family Longevity Selection Score, which is a summary-measure based on the survival experience of the oldest living generation of siblings relative to what would be expected based on birth cohort life tables. 37
In Denmark, individuals who would be aged 90+ years during the study recruitment period were first identified in the Danish National Register of Persons. 38 Then, using information on the place of birth and the names, parish registers available in regional archives were searched to locate the parents of the elderly individuals to identify sibships. The identified subjects were contacted to further assess the family's eligibility for participation in the LLFS using criteria parallel to that used in the United States.
Information from the 4954 US and Danish LLFS participants was collected using similar questionnaires and in-home physical examinations at baseline between 2006 and 2009. Information regarding onset of diseases was assessed retrospectively at baseline from self-reports. The LLFS participants have been followed longitudinally. Because prospective information on ages at onset of diseases is very limited (available currently through April, 2013), only retrospective information was used in this study.
Biospecimens were collected at baseline. Genotyping of the APOE polymorphism was conducted using procedures detailed elsewhere. 40 The data include information on the APOE e2/3/4 polymorphism for the 4659 LLFS participants, consisting of long-living individuals (n=1384, probands and siblings), their offspring (n=2321), and 177 spouses of long-living individuals and 777 spouses of offspring. Due to the small number of spouses of the long-living individuals, they were pooled together with spouses of offspring (n=954).
Analysis
Outcomes considered in this paper include ages at onset of diseases of the heart, cancer, and neurodegenerative disorders (ND). Diseases of the heart assessed in the LLFS included myocardial infarction (MI), congestive heart failure (CHF), coronary angioplasty (CA), and atrial fibrillation (AF). Cancer included all sites except skin. Neurodegenerative disorders included dementia and/or Alzheimer's disease.
We considered differences in the risks of the outcomes for carriers and non-carriers of the APOE e4 allele, defined as the e2/4, e3/4, and e4/4 genotypes for the carriers and the e2/2, e2/3, and e3/3 genotypes for the non-carriers.
Associations of the e4 allele with risks of the selected diseases were characterized by the Kaplan–Meier estimator and the Cox proportional hazard regression model in the samples of survivors selected for the LLFS and examined at baseline. Accordingly, only retrospective information on ages at onset of diseases was retained for such analyses. The time variable in the analyses was the age at onset of a disease or the age at interview at baseline to represent right censoring. All models were adjusted for field center and for birth cohorts measured by age at baseline; other adjustments were explicitly stated when applicable. The proportionality of the hazards was inspected by visual inspection of the survival curves. The cut offs in age were chosen based on the number of people in the groups. The analyses were conducted in men and women separately. We used a robust sandwich estimator of variances in the Cox model to account for potential clustering (e.g., familial). Statistical analyses were conducted using SAS (release 9.3, Cary, NC).
Table S1 (Supplementary Data are available at
Results
Probability of remaining free of cancer
Figure 1A shows that long-living men carrying the e4 allele can potentially be protected against cancer. This protective effect appears to be limited to onsets of cancers at ages older than 75 years, which resembles a similar finding of protective effect of this allele in two independent populations of older males from the FHS original and Offspring cohorts. 34 The e4 allele may also be protective against cancer in sons of the long-living parents at older ages in the LLFS (Fig. S1B). Male spouses carrying the e4 allele contract cancer at younger ages compared to non-carriers of this allele, but the effect of this allele can change at older ages (Fig. S1C). Empirical analyses also suggest trends for a detrimental effect of the e4 allele in long-living women and for a protective effect in female offspring and spouses (Fig. S1D–F).
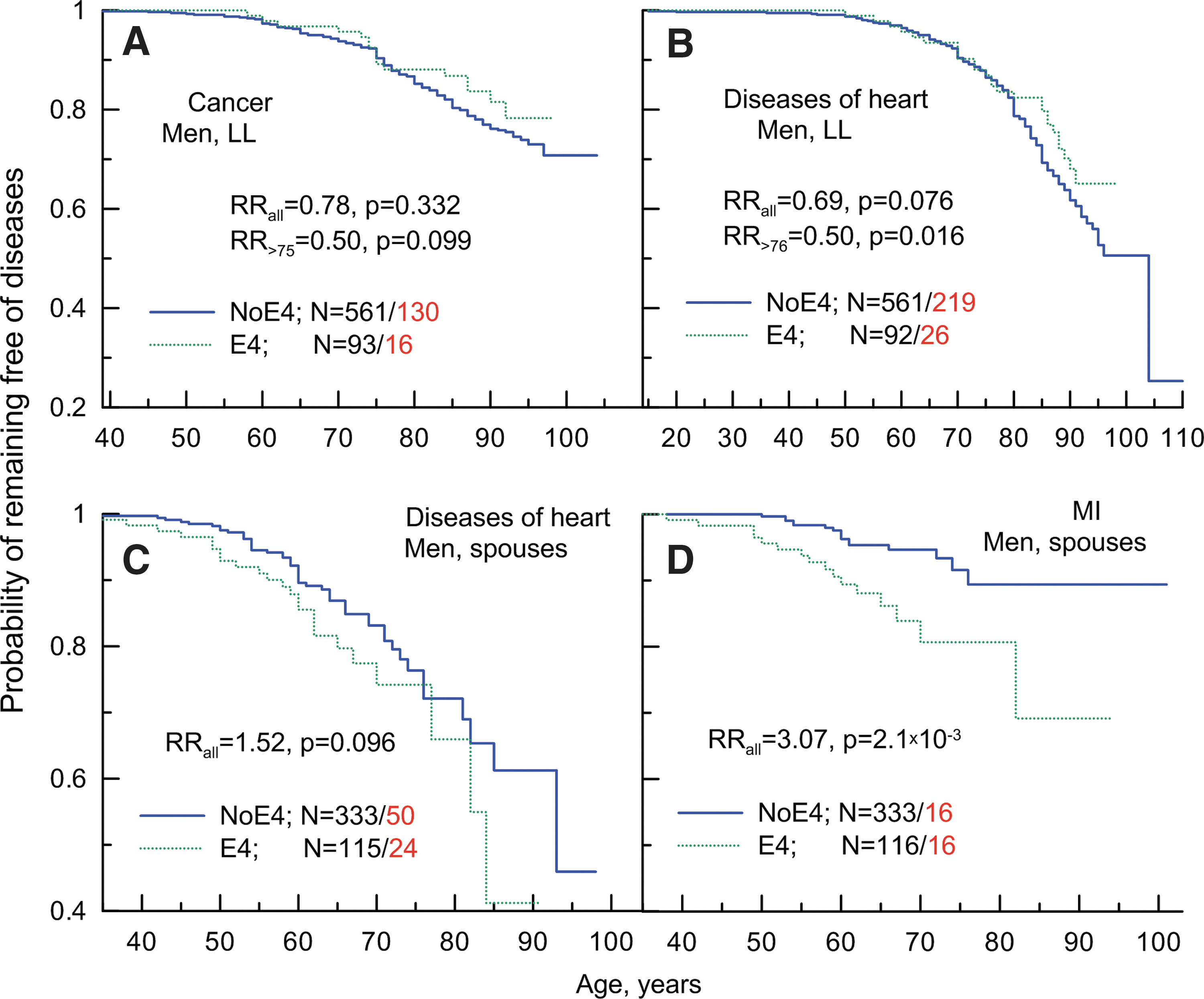
Empirical age patterns of probability of remaining free of cancer (
Analysis of the relative risks (RRs) of cancer using the Cox proportional hazards regression model (Table S2, All) underestimates the effects because it disregards age-related heterogeneity, which is seen as a disproportional hazard (Figs. 1A, and S1). Focusing on the more homogeneous group of individuals who had onset of cancer or were right censored at ages older than 75 years, the estimates of the RRs for men become substantially improved (Fig. 1A, see RRs in the inset and Tables 1 and S2). Given the potentially protective role of the e4 allele in sons of the long-living parents (Fig. S1B), these samples can be pooled together. However, because only a few sons were aged older than 75 years (Fig. S1B), the improvement in the estimates of the RRs for the pooled older sample was minor (Table 1; RR=0.48, p=0.086). The estimates of the RRs in women with potential disproportionality in risks (Fig. S1F) were not refined because of insufficient number of cancer cases at older ages for such analyses.
All models were adjusted for birth cohorts measured by age at baseline and the field centers. Models for pooled samples of parents and offspring (LLFS_P+O), and offspring and spouses (LLFS_O+S) are additionally adjusted for differences between long-living individuals, spouses of long-living individuals, offspring, and spouses of offspring, as applicable. These adjustments do not substantially modify the estimates implying that these factors play at most minor mediating role.
Notations in the “Age group” column indicate the range of ages at onset of cancer or right censoring. “All” denotes the sample of all ages; “>75” implies a group of individuals who had onset of cancer at ages older than 75 years or was right censored at baseline (see Methods).
RR, relative risk; CI, confidence interval; Ntotal and Ncancer, the total number of genotyped individuals and the number of onsets of cancer among them, respectively; LLFS_P, long-living participants of the Long Life Family Study (LLFS); LLFS_O, offspring of the LLFS long-living individuals; LLFS_S, spouses of the LLFS long-living individuals and offspring.
Probability of remaining free of diseases of the heart
Figure 1B shows that long-living men carrying the e4 allele can be also protected against diseases of the heart. As in the case of cancer, this protective effect appears to be limited to onsets at older ages (about 77 years and older). No role of the e4 allele is seen in risks of diseases of the heart in older sons of the long-living parents (Fig. S2B). Male spouses carrying the e4 allele appear to be at higher risks compared to the non-e4 carriers (Fig. 1C). This detrimental effect is not sensitive to the ages at onset. Empirical estimates also suggest a potential detrimental effect of the e4 allele on risks of diseases of the heart in long-living women (Fig. S2D). No effect of the e4 allele is seen in female offspring (Fig. S2E) or spouses (Fig. S2F).
Unlike cancer, however, empirical estimates in the case of diseases of the heart in the long-living individuals, particularly in men, are affected by cohort specifics and by differences between field centers. Evaluating the RRs in long-living individuals, disregarding age-related heterogeneity and using unconditional and conditional models, shows that the estimates for long-living men improve, attaining marginal significance which strengthens the protective role of the e4 allele (Fig. 1B, Table 2).
Unconditional model. Other models were adjusted for birth cohorts measured by age at baseline and the field centers.
Ntotal and NHD, the total number of genotyped individuals and the number of onsets of diseases of heart (HD) among them, respectively; RR, relative risk; CI, confidence interval; LLFS_P, long-living participants of the Long Life Family Study (LLFS); LLFS_O, offspring of the LLFS long-living individuals; LLFS_S, spouses of the LLFS long-living individuals and offspring.
These analyses do not support either protective or detrimental role of the e4 allele in male offspring or in different samples of women (Table S3, All). Empirically manifested age-independent detrimental effect of the e4 allele in male spouses (Fig. 1C) attains suggestive significance (Fig. 1C and Table 2). The analyses show that the protective effect of the e4 allele becomes substantially improved in long-living men beginning at ages at onset of diseases of the heart older than 76 years (see RR>76 in Fig. 1B and Table 2).
Unlike cancer, we had a relatively large number of cases with specific heart pathologies in the studied samples. This allowed us to address the question of whether or not the observed associations of the e4 allele with diseases of the heart are explained by their specific forms (see Methods). The analyses show that protective effect at older ages is distributed uniformly across all heart pathologies assessed in the LLFS, a result that suggests that protective effect can unlikely be attributed to a specific form of heart disease (Table 3). In contrast, the detrimental effect of the e4 allele in male spouses is primarily due to MI (Table 3) and it is pronounced regardless of age at onset (Fig. 1D). The differential role of the e4 allele across these samples in different heart pathologies can unlikely be explained by the sample size differences (Table 3).
Note that the number of onsets of specific forms of diseases of heart (HD) do not sum to the number of onsets of HD because the same individual can have different forms of HD whereas only the youngest age at onset is considered for a given individual.
Ntotal and NHD, the total number of genotyped individuals and the number of onsets of diseases of heart (HD) among them, respectively; RR, relative risk; CI, confidence interval; MI, myocardial infarction; CHF, congestive heart failure; CA, coronary angioplasty; AF, atrial fibrillation; LLFS_P, long-living participants of the Long Life Family Study (LLFS); LLFS_O, offspring of the LLFS long-living individuals; LLFS_S, spouses of the LLFS long-living individuals and offspring.
Probability of remaining free of ND
Due to the small number of onsets of ND, the analyses were conducted in the pooled sample of all family members (virtually all onsets of ND were at ages of 78 years and older). Table 4 shows that both men and women carrying the e4 allele may be at risk of ND. This detrimental effect attains nominal significance in the pooled sample of men and women.
Ntotal and NND, the total number of genotyped individuals and the number of onsets of Alzheimer disease or dementia (ND) among them, respectively; RR, relative risk; CI, confidence interval.
Discussion and Conclusions
Genetic association studies, including those using genome-wide resources, often implicitly assume deterministic (unconditional) linkage of genes with age-related traits. A direct consequence of this hypothesis is an assumption of population genetic risks. However, experimental evidence and evolutionary constraints provide little support for this assumption. 41 As a result, one has to assume indirect (conditional) linkage of genes with age-related traits. 42 This change in the paradigm from unconditional to conditional risks requires a change in analytic strategies. That is, rather than attempting to evaluate unconditional population genetic risks, comprehensive analyses of the mechanisms that shape the conditional risks of genes on age-related traits in a given environment are required. 43 Guided by this paradigm, we extended our prior analyses of trade-offs in the effects of the APOE e4 allele on major human diseases in the FHS, 18,34 examining phenotypic mechanisms that can drive such trade-offs in a specific population of the LLFS families enriched for chances of exceptional longevity.
The e4 allele and risk of cancer
We found that long-living men carrying the e4 allele might be protected against cancer at ages older than 75 years (Table 1). A protective effect of the e4 allele against cancer was also documented in two independent samples of genotyped participants of the original and offspring cohorts in the FHS. 18,34 The potentially protective effect in the LLFS is concordant with findings in the FHS, not merely in the effect direction, but also in two additional aspects, i.e., it is seen at the same old ages and primarily in men. The role of the e4 allele in cancer in other groups of the LLFS participants is uncertain largely due to insufficient numbers of cancer cases in these groups of survivors.
The finding of the protective effect of the e4 allele against cancer in men at the same old ages in different populations has several implications. First, this finding suggests that aging-related processes can shape genetic effects on cancer. Second, this cancer-protective mechanism is sex specific and should be more pronounced in men than in women. Third, this finding contributes to thoughts on pure genetic (deterministic) and gene–environmental origins of age-related traits (see the beginning of Discussion and Conclusions section). Indeed, it suggests the existence of a cancer-protective genetic mechanism that should be weakly sensitive to the environment, i.e., such a mechanism may have relatively wide norm of genetic reaction to environmental exposures. This is evident from the observation of the associations of the e4 allele with cancer in different populations and generations of older men, i.e., the same generation of men in the FHS original cohort and in the cohort of the LLFS long-living men and offspring generation of men in the FHSO. Generations are, particularly, a proxy for environmental exposures in which individuals from these cohorts grew up. Age-sensitivity of the observed protective effects suggests that such a mechanism may be relevant to aging-related processes.
The e4 allele and risks of diseases of the heart
Long-living men appear to be also protected against diseases of the heart at virtually the same ages as in the case of cancer, i.e., at ages older than 76 years (Table 2). This effect is concordant with the protective effect of the e4 allele against cardiovascular diseases observed in a more homogeneous subsample of the FHS original cohort in terms of the effect direction and expression at the same old ages. 34 This result suggests that aging-related processes can shape the effect of the e4 allele not only on cancer but also on cardiovascular diseases. However, the age-specific protective effect of the e4 allele was seen in the LLFS men, whereas in the FHS it was seen in women. These results suggest that mechanisms mediating age-sensitive effects of the e4 allele on cardiovascular diseases are less conserved than in the case of cancer, and they can be modulated by sex. No reliable conclusions can be drawn on the role of the e4 allele in the LLFS women and male offspring, largely due to the limited number of disease cases.
The analyses show that protective effect of the e4 allele in the LLFS long-living men is not associated with specific heart pathology, but rather can be a superposition of protective effects against different forms of diseases of the heart (Table 3). This result suggests the existence of a systemic (disease non-specific) mechanism that can be involved in regulation of not just one trait, but perhaps a major subset of them. 5,6 Indeed, although atherosclerosis can be a common risk factor for MI, CHF, and CA, the other heart pathology, AF, is typically due to another mechanism associated with abnormal spreading of electrical signals in the heart. This result is strengthened by the protective effect of the e4 allele against such a pathology-distinct disease as cancer (see the section above). Sensitivity of such a systemic protective mechanism to age suggests its connection with aging-related processes.
A fundamentally different effect of the e4 allele is observed in male spouses. First, we observe a detrimental but not protective (as in long-living men) effect. Second, this detrimental effect is seen regardless of age at onset (Fig. 1C, D). Third, this effect is primarily attributed to specific heart pathology, i.e., MI (Table 3).
Qualitative differences in the observed effects of the e4 allele across the studied populations may reflect specifics of these populations. Indeed, unlike the LLFS long-living men, the LLFS male spouses were not selected for exceptional longevity and, thus, they represented an ordinary population as in other studies. Prior studies in various populations of individuals not selected for their exceptional longevity typically reported an adverse effect of the e4 allele on coronary heart disease 44,45 that was concordant with our observations.
These results support the existence of qualitatively different forms of actions of the e4 allele on diseases of the heart through disease-specific and systemic, likely aging-related, mechanisms.
The e4 allele and onset of ND
Concordant with numerous studies on the role of the e4 allele in ND, the LLFS participants carrying the e4 allele are at higher risks of ND than the non-e4 carriers. This result should be interpreted with caution, however, because participants' recollection of their ages at onset of ND is less accurate than that on CVD and cancer.
The role of the e4 allele in health span
Thus, the results of our analyses show a complex role of the APOE e4 allele in health span when the same e4 allele can diminish risks of cancer and diseases of the heart, and, depending on the LLFS population, confer risks of diseases of the heart and neurological disorders. This complexity highlights trade-offs in the effect of the e4 allele on risks of major human diseases. Accordingly, this result extends a finding of genetic trade-offs in the FHS 18,34 to surviving individuals who participated in the LLFS. Potential mechanisms of genetic trade-offs are discussed in references 18, 34, and 46.
Our results contribute to the discussion that the traditional concept regarding replication of association of the same allele, in the same direction with the same aging-related trait in different populations, has inherent limitations. 41,47 –51 Additionally, our results support the view that more insight into the genetic origin of traits in late life and analyses of mechanisms linking genes to such traits are needed.
Our analyses suggest that the e4 allele is associated with risks of the selected diseases through two different mechanisms. One mechanism is likely associated with biochemical genetic basis of specific traits (conferring risks of MI and ND), which is traditionally the most-studied mechanism. The analyses show that this disease-specific mechanism is not sensitive to ages at onset, particularly of MI, in this study.
A key feature of the other mechanism is its age-sensitivity when the protective effect of the e4 allele against both cancer and diseases of the heart becomes expressed at the same old ages of mid-70s and older. This mechanism is likely relevant to aging-related processes and potentially can explain the risks of subsets of diseases in the elderly, including various forms of diseases of the heart and cancer.
Both of these mechanisms, i.e., disease-specific and systemic (aging-related), are mostly expressed in LLFS men. Importantly, the protective aging-related mechanism is seen in long-living men, whereas a detrimental MI-specific mechanism is seen in the population of male spouses who have not been selected for their chances of exceptional survival. This finding suggests that individuals with exceptional longevity can have protective mechanisms assuring their survival to old ages. 52 Given that men have a shorter life span compared to women, men surviving to extreme ages should have male-specific protective mechanism(s). Exceptional survival of men in the LLFS may be assured by such mechanism(s) because these mechanism(s) can cluster in specially selected long-living LLFS families. Our results suggest that long-living women in the LLFS may not need to have the APOE-related protective mechanism to live long lives. Then, the findings of the potential protective role of the e4 allele against cancer and diseases of the heart in long-living men may highlight sexual dimorphism in longevity-assuring mechanism(s). Sexual dimorphism in the observed effects of the APOE e4 allele can be linked to differential hormonal and insulin regulation in men and women. 53
We should underline that the LLFS is a family-based cohort study examining the genetic and non-genetic factors associated with exceptional familial longevity (see Methods). Long-lived individuals, their siblings, and their offspring and spouses were recruited for an examination that characterized key intermediate phenotypes of longevity, including major chronic diseases, risk factors, and physical and cognitive function. Therefore, the LLFS cohort is a highly selected group of exceptional families and not representative of the overall general population. Thus, caution is needed when interpreting and extrapolating these results to other populations because they have limited generalizability under the hypothesis of “pure genetic” origin of complex traits. In fact, in our case of retrospective data, the observed associations are characteristic for individuals who survived until enrollment into the LLFS despite contracting diseases of the heart or cancer.
Footnotes
Acknowledgments
The Long Life Family Study is funded by U01AG023749, U01AG023744, and U01AG023712 from the National Institute on Aging (NIA). The research reported in this paper was supported by the National Institute on Aging (2 U01 AG023746-09).
Author Disclosure Statement
No competing financial interests exist.
A.M.K. contributed to the study conception, design, statistical analysis, interpretation of the results, and writing the manuscript. K.G.A. contributed to preparation of the data and statistical analyses. I.C. contributed to the study design and interpretation of the results. S.V.U. contributed to final version and discussion of mechanisms of genetic trade-offs. M.P. contributed to discussion of the manuscript. E.S. and A.I.Y. contributed to assessing the logic of the final version and discussion of the results.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.