
Abstract
Regular exercise has multi-system anti-aging effects. Here we summarize how exercise impacts the major hallmarks of aging. We propose that, besides searching for novel pharmaceutical targets of the aging process, more research efforts should be devoted to gaining insights into the molecular mediators of the benefits of exercise and to implement effective exercise interventions for elderly people.
Introduction
T
Summary of the Impact of Aging on the Main Physical Fitness—Related Phenotypes
Aging and cardiorespiratory fitness
Maximal oxygen uptake (VO2max; sometimes referred to as maximal aerobic capacity or simply aerobic capacity or aerobic endurance) is a main indicator of cardiorespiratory fitness. There is variability among studies, 2 –4 but the average rate of VO2max decline in old people is ≥4–5 mL·kg−1·min−1/decade. 5 Mostly reduced maximal cardiac output 6 –10 but also a decline in maximal arteriovenous oxygen difference (a-vO2 diff) (minus ∼3%/decade) 7,11 contribute to age reductions in VO2max. 12,13 Aging skeletal muscles have a low capacity to use O2 due to several factors, such as decreased muscle mass (see below), 14,15 increased peripheral resistance, 16 reduced muscle capillary density, 17 endothelial dysfunction, 18 changes in skeletal muscle microcirculation, 19 and reduced muscle oxidative capacity. 20
Aging and muscle function
Muscle mass usually starts to decline after 25–30 years of age, 21,22 such that on average 40% of muscle mass is lost by 80 years. 22,23 In turn, a quantitative loss in muscle cross-sectional area is a major contributor to the decrease in muscle strength seen with advancing age, i.e., after 60–70 years of age. 24 The term “sarcopenia” was originally created to refer to age-related loss of muscle mass with consequent loss of strength. 25 There are now four international definitions of sarcopenia. 26 –29 In essence they all agree, requiring a measure of impaired walking capability (either low gait speed or a limited endurance [distance] in a 6-min walk), together with an appendicular lean mass of less than 2 standard deviations of a sex- and ethnically-corrected normal level for individuals 20–30 years old. Sarcopenia (see below for details on signaling pathways involved) occurs due to several age-related factors, such as gradual muscle denervation, 23,30 diminished satellite cells, 31 low muscle protein synthesis, 32 low anabolic hormone levels, 33 malnutrition, 34 increased pro-inflammatory cytokines, 35 oxidative stress, 36 mitochondrial dysfunction, 37 –40 and physical inactivity. 41 Although there are differences between studies, on average, 5%–13% and 11%–50% of people aged 60–70 years and ≥80 years, respectively, suffer sarcopenia, 42 –47 with a higher prevalence (68%) reported in nursing home residents ≥70 years. 48 Sarcopenia needs to be differentiated from “cachexia,” a combination of both muscle and fat loss that is usually attributable to an excess of catabolic cytokines associated with a disease process, e.g., cancer. 49 –51 Sarcopenia is linked with increased disability, falls, hospitalization, nursing home admission, and mortality. 48,52 –54
Frailty and disability
A consequence of the aforementioned effects of aging on cardiorespiratory and muscle fitness (especially sarcopenia), alone or in combination with co-morbidities such as neurodegeneration, is the “frailty syndrome.” 55 Although no single operational definition of the frailty syndrome has been agreed upon, two major definitions with proposed assessment tools have predominated over the past decade—the frailty phenotype, also known as Fried's definition, 56 and the frailty index. 57 The most widely cited is the frailty phenotype, which is operationalized as a syndrome meeting three or more of the following five phenotypic criteria: Weakness as measured by low grip strength, slowness (low walking speed), low level of physical activity, low energy or self-reported exhaustion, and unintentional weight loss. 56 A prefrail stage, in which one or two criteria are present, identifies a subset at high risk of progressing to frailty. Older individuals with none of the above five criteria are classified as nonfrail. The frailty index was developed on the basis of a comprehensive geriatric assessment by counting the number of deficits accumulated, including diseases, physical and cognitive impairments, psychosocial risk factors, and common geriatric syndromes other than frailty. 57,58 Whereas the frailty index may have clinical utility in risk assessment and stratification, it is less clear that it adds significant value to comprehensive geriatric assessment. 59
Frailty is an important and growing problem in aging western populations, because it potentially affects 20%–30% of adults older than 75 years. 60 For instance, the prevalence of prefrail and frail individuals is high in the Spanish population (41.8% and 8.4%, respectively) and increases with age, according to a recent a population-based study conducted on 2488 individuals aged 65 years and older in a central area of the country. 61 On the other hand, frailty could eventually result in disability, 62 , i.e., “difficulty or dependency in carrying out activities necessary for independent living, including roles, tasks needed for self-care and household chores, and other activities important for a person's quality of life.” 63
For the above-mentioned reasons, it is of paramount medical importance to attenuate age-related declines in physical fitness. Yet no single drug can reverse the age-related loss of physical fitness because none benefits all of the systems involved, whereas regular exercise has multi-system anti-aging effects (see below and Fig. 1, left column for a summary).

Summary of the main anti-aging effects of regular exercise vs. aging effects at the multi-systemic (left; see Chodzko-Zajko et al.
64
for an in-depth review) and cellular level (right; see text). AKT, protein kinase B; AMPK, AMP-activated protein kinase; ASC, apoptosis-associated speck-like protein caspase; AUF1, AU-binding factor 1; BDNF, brain-derived neurotrophic factor; FoxO3a, human protein encoded by the FOXO3 gene; Glut 4, glucose transporter type 4; HATs, histone acetyltransferases; HDACs, histone deacetylases; HRV, heart rate variability; IGF-1, insulin-like growth factor 1; IL, interleukin; jmjC, jumonji C proteins; LSD, lysine-specific histone demethylase; miR, micro-RNA; mtDNA, mitochondrial DNA; mTOR, mammalian target of rapamycin; NK cell, natural killer cell; NLRP3, NOD-like receptor protein 3; PBMCs, peripheral blood mononucleated cells; PDK4, pyruvate dehydrogenase kinase isoenzyme 4; PGC-1, peroxisome proliferator-activated receptor gamma coactivator 1; PPAR-δ, peroxisome proliferator-activated receptor δ; PTMs, post-translational modifications; Qmax, maximal cardiac output; ROM, range of motion; SIRT, sirtuin; TERT, human telomerase reverse transcriptase. Color images available online at
Exercise Attenuates Fitness and Multisystem Age-Related Declines
Benefits of exercise (particularly aerobic exercise) in cardiorespiratory fitness/cardiovascular disease
Regular exercise, particularly dynamic exercise of moderate intensity (≤70% of VO2max or ≤80% of maximum heart rate) involving mostly the aerobic energy pathway and large muscle mass (e.g., brisk walking, bicycling) attenuates age declines in cardiorespiratory fitness (see Chodzko-Zajko et al. 64 for an in-depth review and experts' recommendations). This type of exercise, commonly referred to as “aerobic exercise” (or “endurance exercise”), has a restoring effect on an important risk factor of cardiovascular disease (CVD), i.e., endothelial dysfunction. 65 –67 It also increases endothelial nitric oxide (NO•) production and thus vascular tone regulation. Regular bouts of exercise-increased laminar flow activate (through phosphorylation via protein kinase B, Akt) endothelial NO• synthase while attenuating NO• degradation into reactive oxygen species (ROS) and reactive nitrogen species. 68 Together with increased angiogenesis (see further below), an additional benefit of aerobic exercise in endothelial health is stimulation of macrophage-mediated reverse cholesterol transport through activation of peroxisome proliferator-activated receptor gamma (PPARγ). 69,70 Yet aging autonomic dysfunction has a synergistic effect together with endothelial dysfunction in increasing CVD risk 71 and raises the risk of a leading cause of death in most industrialized countries, sudden death due to ventricular fibrillation. 72
During aging, the sympathetic nervous system (SNS) outflow to several peripheral tissues increases to stimulate thermogenesis and thus to prevent increasing adiposity. 73 Chronically activated SNS has deleterious effects on the cardiovascular system, i.e., reduced leg blood flow, increased arterial blood pressure, impaired baroreflex function and hypertrophy of large arteries; it can also increase insulin resistance, thereby raising the risk of metabolic syndrome. 74,75 Importantly, aerobic exercise training (e.g., brisk walking) has a beneficial, dose–response effect in attenuating aging autonomic system dysfunction, with trained elderly individuals showing similar baroreflex function compared with their moderately active younger peers. 76 Heart rate variability (HRV), a marker of autonomic function and a powerful predictor of CVD outcome (high HRV is associated with a better prognosis), increases with aerobic exercise training in old people. 77 Reduced angiotensin II, increased NO•, and mainly improved vagal modulation and decreased sympathetic tone are implicated in the beneficial exercise effects on HRV. 78
Benefits of exercise (particularly resistance exercise) in the aging muscle function
Training programs, especially if including resistance (strength) exercises (i.e., movements, such as weightlifting or exercises with resistance bands, performed against a specific external force that is regularly increased during training) are especially useful for improving muscle mass and/or strength in the elderly, 79 including in the oldest old 80 (see Table 1 for a summary of controlled exercise interventions [mostly based on resistance exercises] in the oldest old). In general, published resistance exercise interventions in old people had a duration ranging from 4 to 48 weeks and were in agreement with accepted or “traditional” exercise recommendations for older people, 81 with the usual weekly schedule including two to three nonconsecutive sessions with one to three sets of 10–15 repetitions of classic weightlifting exercises, such as leg presses.
Of note, the mean age of the subjects in all the studies was ≥ 80 years.
Data provided by the total number of participants.
BMI, Body mass index; CHO, carbohydrate; CSA, cross-sectional area; FAC, functional ambulation classification; NA, not available; PA, physical activity; RPE, rating of perceived exertion (Borg's conventional scale, 6–20 points); 1RM, one repetition maximum; SD, standard deviation; SO, social activities; TG, training group; UCG, usual care group; WBV, whole-body vibration.
Other interventions feasible in old people include “high-velocity resistance training,” i.e., focusing on speed of movement, 82,83 or even explosive-type heavy-resistance training, e.g., weightlifting exercises with a load equivalent to 75%–80% of one repetition maximum (1RM) and performed with maximal intentional acceleration of the training load during the concentric movement phase. 84 In general, besides being feasible and well tolerated even by the oldest old, 84 this alternative type of intervention would elicit similar 85 or even higher improvements in functional performance and disability compared with more traditional, lower-velocity resistance training. 86 Another approach is the use of a weighted vest, which has proved effective to improve perceived health in old people, 87 as well as lateral stability, lower-body muscular strength, muscular power, and leg lean mass (and thus likely to reduce risk fall) in postmenopausal women aged 50–75 years. 88 Preliminary findings also support the potential effectiveness of including a weighted stair climbing exercise in the training programs of old people. 89 Others, however, found no significant benefits of a home-based intervention using a weighted vest. 90
Benefits of exercise in the frail elderly
Although numerous studies have shown the benefits of resistance training in old people in general, less research has focused specifically on the frail elderly. This is an important issue because frailty status might influence the potential applicability and effectiveness of the different training programs. For instance, Faber et al. found that fall-preventive, moderate-intensity, group exercise programs (mostly based on walking and balance exercises) had positive effects on fall risk and physical performance in pre-frail but not in frail elderly (mean age 85 years). 91
Fiatarone et al. reported that physically frail, oldest old men and women improved their leg muscle strength outcomes by 220% after a 10-week strength training program (three sets of eight repetitions at 80% of 1RM, three times a week). 80 Serra-Serra-Rexach et al. found an increment of 17% in the leg press strength of nonagenarians after 8 weeks of progressive strength training with lower loads (two to three sets of eight to 10 repetitions at 30% of 1RM in the initial phase progressing to 70% of 1RM at the end of the intervention). 92 Lustosa et al. found significant improvements in the knee extensor muscle power of pre-frail elderly women after 12 weeks of strength exercises of the lower extremities at 70% of 1RM, three times a week. 93 Although more research is needed, higher-intensity exercises seem to elicit higher gains. Thus, Sullivan et al. 94 found greater increases in the muscle strength of frail elderly after a 12-week resistance training program of progressive intensity (starting at 20% and ending at 80% of 1RM) compared with a continuous, lower intensity protocol (20% of 1RM during the entire 12-week period).
The American College of Sports Medicine recommends a multi-component (strength, endurance, flexibility, and balance) exercise program to maintain physical fitness in old adults. 95 Villareal et al. studied the effect of such a multi-component exercise program on physical fitness in frail, obese older adults during 3 months, finding beneficial effects on muscle mass and physical function. 96 Lord et al. found significant improvements in choice stepping reaction time test, 6-min walking distance, and simple reaction time requiring a hand press, after training (aerobic exercises, specific strengthening exercises, and activities for balance, hand-eye and foot-eye coordination, and flexibility) during 12 months in frail older people living in retirement villages. 97 Eshani et al. 98 assessed the effect of a 6-month aerobic training program on frail elderly (consisting of walking at 70%–75% of maximal heart rate during 20 min at the initial phase and progressing to 60 min at the end of the program) and found a 14% of increase in VO2max.
Evidence on the benefits of exercise interventions in elderly with frailty, co-morbidities, and subsequent physical disability comes from a recent meta-analysis by deVries et al. 99 This study included data on 18 randomized controlled trials (2,580 participants in total) of physical training interventions in community-dwelling adults aged 60–85 years who were physically frail and/or had physical disability and/or multi-morbidity. Half of the included studies used multi-component training programs, and intervention duration ranged from 5 weeks to 18 months. There were statistically significant benefits with physical exercise therapy compared to no exercise in mobility and physical functioning. There were no differences in effectiveness with regard to the duration of the program (short vs. longer interventions), whereas high-intensity programs elicited greater gains compared to low-intensity ones. The interventions that showed the largest effect sizes were those using resistance training components.
Benefits of exercise (particularly, resistance exercise) in the aging muscle tissue—main signaling pathways involved in exercise adaptations
The molecular mechanisms involved in the activation of signal transduction cascades regulating the adaptations to exercise are dependent on specific signaling pathways activated or repressed by numerous stimuli such as alterations in metabolites concentrations, adenosine triphosphate (ATP)-to-adenosine diphosphate (ADP) ratio, calcium flux, intracellular pH, redox balance, ROS production, and intracellular oxygen pressure. 100 –103 Post-exercise changes in gene transcription involve immediate early genes, myogenic regulators, genes that regulate carbohydrate metabolism and lipid mobilization, transport and oxidation, mitochondrial metabolism, and oxidative phosphorylation, as well as transcriptional regulators of gene expression and mitochondrial biogenesis, 104 –107 or alterations in the DNA-binding activity of a variety of transcription factors, such as myocyte enhancer factor 2 (MEF2), 107 histone deacetylases (HDACs), 108 and nuclear respiratory factors (NRFs). 109,110
The most relevant signaling pathways modulated by exercise include calcium/calmodulin-dependent protein kinase (CaMKs) signaling, mitogen-activated protein kinases (MAPKs) signaling, ATP turnover and adenosine monophosphate (AMP)-activated protein kinase (AMPK) signaling, oxidized nicotinamide adenine dinucleotide (NAD+)/reduced nicotinamide adenine dinucleotide (NADH) ratio and sirtuins (SIRTs), activation of mammalian target of rapamycin (mTOR), oxygen sensing and the hypoxia-inducible factor 1 (HIF-1), mitochondrial biogenesis pathway, and the PPARγ co-activator-1α (PGC-1α) and −1β(PGC-1β). 111 Accordingly, several important adaptations in skeletal muscle, such as mitochondrial biogenesis, anti-oxidant defense, hypertrophy, cytoprotection, and fiber transformation, are regulated primarily by these pathways. Below we summarize the main biological mechanisms and pathways through which exercise may attenuate sarcopenia (see also Fig. 2).
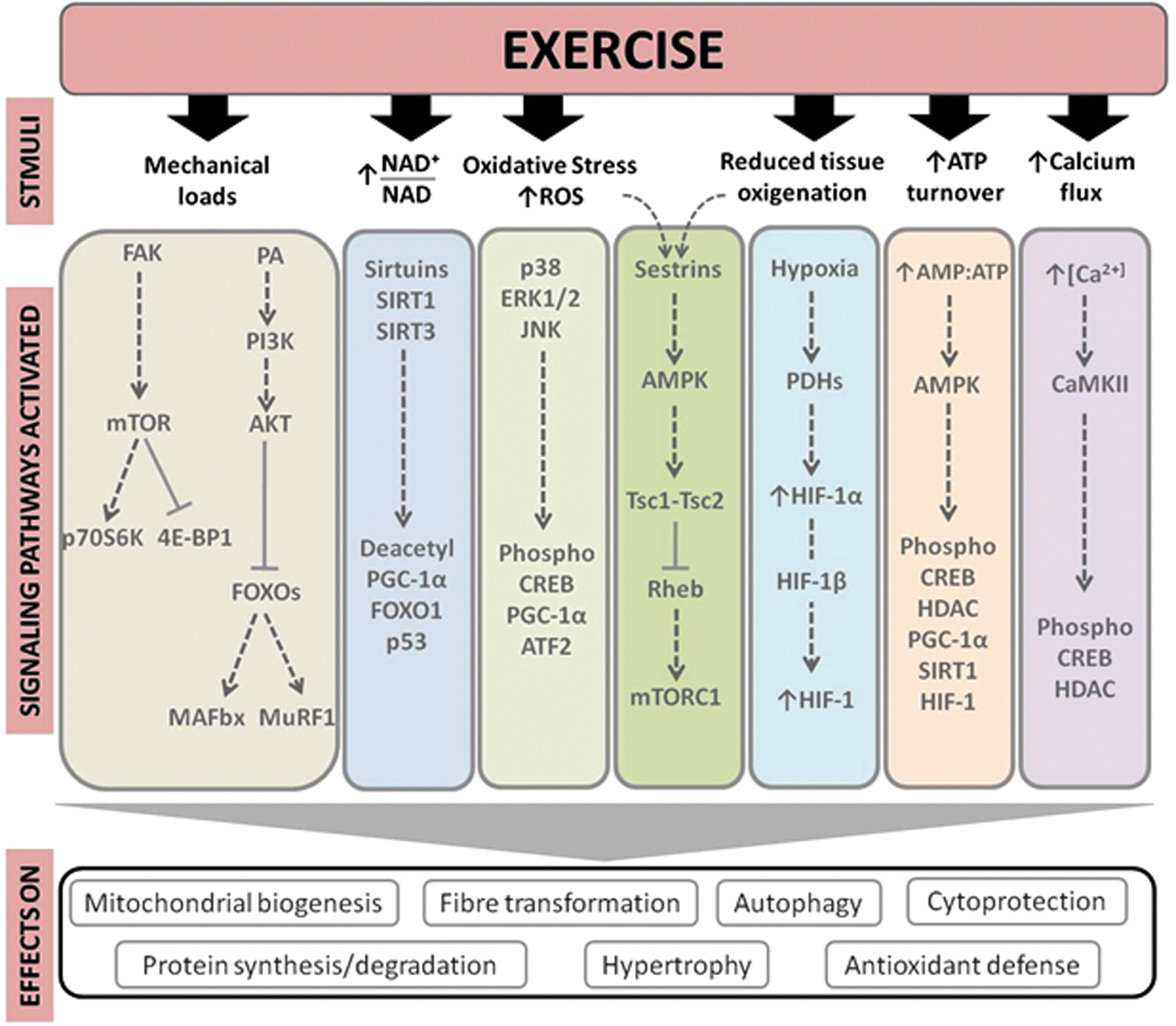
Main signaling pathways involved in the exercise effects in the skeletal muscle tissue. 4E-BP1, eukaryotic translation initiation factor 4E (eIF4E) binding protein; AKT, protein kinase B; AMP, adenosine monophosphate; AMPK, AMP activated protein kinase; ATF2, activating transcription factor 2; ATP, adenosine triphosphate; CaMKII, calmodulin-dependent protein kinase II; CREB, cAMP response-element-binding protein; ERK1/2, extracellular signal-regulated kinase 1 and 2; FAK, focal adhesion kinase; FoxO1, human protein encoded by the FOXO gene; FOXOs, Forkhead box-O transcription factors; HDACs, histone deacetylases; HIF, hypoxia-inducible factor; JNK, c-Jun NH2-terminal kinase; MAFbx, or Atrogin-1; MuRF-1, muscle RING-finger protein-1; mTOR, mammalian target of rapamycin; mTORC1, mTOR complex 1; NAD, nicotinamide adenine dinucleotide; PA, phosphatidic acid; p70S6K, ribosomal protein S6K; PDH, prolyl hydroxylase; PGC-1α, peroxisome proliferator-activated receptor-γ coactivator-1α; PI13K, phosphatidylinositol 3-kinase; Rheb, Ras homolog enriched in brain gene; ROS, reactive oxygen species; SIRT, sirtuin; Tsc1, tuberous sclerosis complex 1; Tsc2, tuberous sclerosis complex 2. Color images available online at
Calcium is implicated in the regulation of numerous intracellular proteins, such as protein kinase C, calcineurin, and CaMKs that mediate cellular signal transduction. 112 Exercise increases CaMKII phosphorylation in an intensity-dependent manner. CaMKs and calcium signaling affect glucose transport, 113 lipid uptake and oxidation, 114 and skeletal muscle plasticity. 115 In addition, the transcription factors cyclic AMP response element-binding protein (CREB), MEF2, and HDACs are CaMK targets involved in the regulation of skeletal muscle gene expression. Exercise also stimulates MAPK-related pathways, including extracellular signal-regulated kinase 1 and 2 (ERK1/2), 116 p38, 117 and c-Jun NH2-terminal kinase (JNK). 116 For instance, p38 MAPK can stimulate upstream transcription factors of the PGC-1α gene through skeletal muscle contraction. 118 Moreover, MAPKs regulate a wide range of physiological processes, such as differentiation, hypertrophy, inflammation, and gene expression. 119
The role of ROS in the exercise-induced adaptations of skeletal muscles has been studied extensively, particularly with regard to aerobic exercise. 120,121 Contracting skeletal muscles produce ROS, activating MAPK signaling and transcription factor nuclear factor-kappa B (NF-κB), thereby linking signal transduction to transcriptional processes. 122 Acute exercise activates JNK signaling in a ROS-dependent manner, as evidenced by attenuated JNK signaling during exercise with infusion of the anti-oxidant N-acetylcysteine. 123 Contraction-induced increases in interleukin-6 (IL-6) secretion, an exercise-associated cytokine with potent multi-organ metabolic effects 124 (see further below), is JNK dependent, 125 which attests to the likely importance of JNK signaling in mediating metabolic adaptations to exercise. AMPK is a serine/threonine kinase that modulates cellular metabolism acutely through phosphorylation of metabolic enzymes 126 and, over time, via transcriptional regulation. 127,128 Given the rate of ATP turnover during muscle contraction, AMPK acts as a signal transducer for metabolic adaptations by responding to an altered cellular energy status. Overall, AMPK activation preserves ATP by inhibiting both biosynthetic and anabolic pathways, while simultaneously stimulating catabolic pathways to re-establish cellular energy stores. 129 Chronic AMPK activation modifies metabolic gene expression and stimulates mitochondrial biogenesis, 127 partly via AMPK-induced modulation of the DNA-binding activity of transcription factors, including NRF-1, MEF2, and HDACs. 127,130
Sestrins are a recently discovered hallmark of aging sarcopenia. Mammalian cells express sestrins in response to stress including DNA damage, oxidative stress, and hypoxia. Sestrins can inhibit the activity of the mTOR complex 1 (mTORC1) through activation of AMPK. 131 Sestrins prevent sarcopenia, insulin resistance, diabetes, and obesity. They also extend life span and health span through activation of AMPK, suppression of mTORC1, and stimulation of autophagic signaling. 131 Recently, we proposed a possible role of the AMPK-modulating functions of sestrins in the benefits produced by exercise in older subjects. 132
The regulation of the SIRT family of protein deacetylases is NAD+ dependent. 133 Both deacetylases SIRT1 and SIRT3 respond to elevations in [NAD+] and the NAD+/NADH ratio. Increased SIRT activity is associated with positive adaptations in skeletal muscle metabolism, including improved mitochondrial function and exercise performance. 134,135 Likewise, the adaptive muscle growth consequent to mechanical loads induced by resistance exercise is largely determined by the enhanced skeletal muscle protein synthesis due to the activation of mTOR, ribosomal protein S6K (p70S6K), and downstream targets. 136 p70S6K is a major regulator of muscle protein synthesis through pathways of protein translation and ribosome biogenesis involving eukaryotic translation initiation factor 4E (eIF4E), 4E binding protein 1 (4E-BP1), and elongation factor 2 (eEF2). Phosphorylation of 4E-BP1 by mTOR suppresses binding and inhibition of eIF4E by 4E-BP1. Phosphorylation of S6K leads to the phosphorylation of the 40S ribosomal protein S6 (rpS6) and eukaryotic translation initiation factor 4B (eIF4B). Collectively these events lead to the formation of the translation initiation complex and activate protein synthesis inducing cellular hypertrophy. 137 Mechanosensory regulation of muscle protein synthesis also involves other signaling proteins, such as focal adhesion kinases (FAK), a class of transmembrane receptors that act as protein tyrosine kinases. FAK proteins are pivotal points for the transmission of contractile force throughout the skeletal muscle structure and a central component of integrin signaling. The grade of expression and activity of FAK in skeletal muscle is loading dependent, 138,139 and contraction results in conformational modifications and activation of FAK phosphotransferase, 138,140 which can trigger muscle protein synthesis through mTOR-dependent or -independent mechanisms. 141
Oxygen sensing is also involved in the adaptations of skeletal muscle fibers to exercise, particularly aerobic exercise. Hypoxia-inducible factor (HIF), a heterodimeric transcription factor composed of two subunits (HIF-1α and HIF-1β), regulates the major signal transduction pathway sensitive to the intracellular partial pressure of oxygen. Activation of HIF-1 by many stimuli, including aerobic exercise, induces transcription of target genes involved in erythropoiesis, angiogenesis, glycolysis, and energy metabolism. 105,142,143
Exercise benefits in neurodegeneration—main signaling pathways involved
Physical exercise produces important benefits in several neurodegenerative diseases. 144,145 Here we focus on the effects of exercise in Alzheimer's disease, which comprises per se 50%–56% of all causes of aging dementia (an additional 13%–17% is caused by Alzheimer's disease combined with vascular diseases). 146 Exercise, especially aerobic exercise, is beneficial for patients with Alzheimer's disease, 147 not only because it attenuates patients' physical and psychosocial dependence 148 but it also because it improves several features of the pathophysiology of this disorder, 149,150 including oxidative stress regulation, autophagy systems, neurotrophic signaling, mitochondrial biogenesis, angiogenesis, neurogenesis, and the modulation of specific amyloid-β (Aβ)-degrading enzymes (see Table 2 for a summary of intervention studies in humans and Fig. 3 for a summary of the putative molecular pathways involved). 149,150
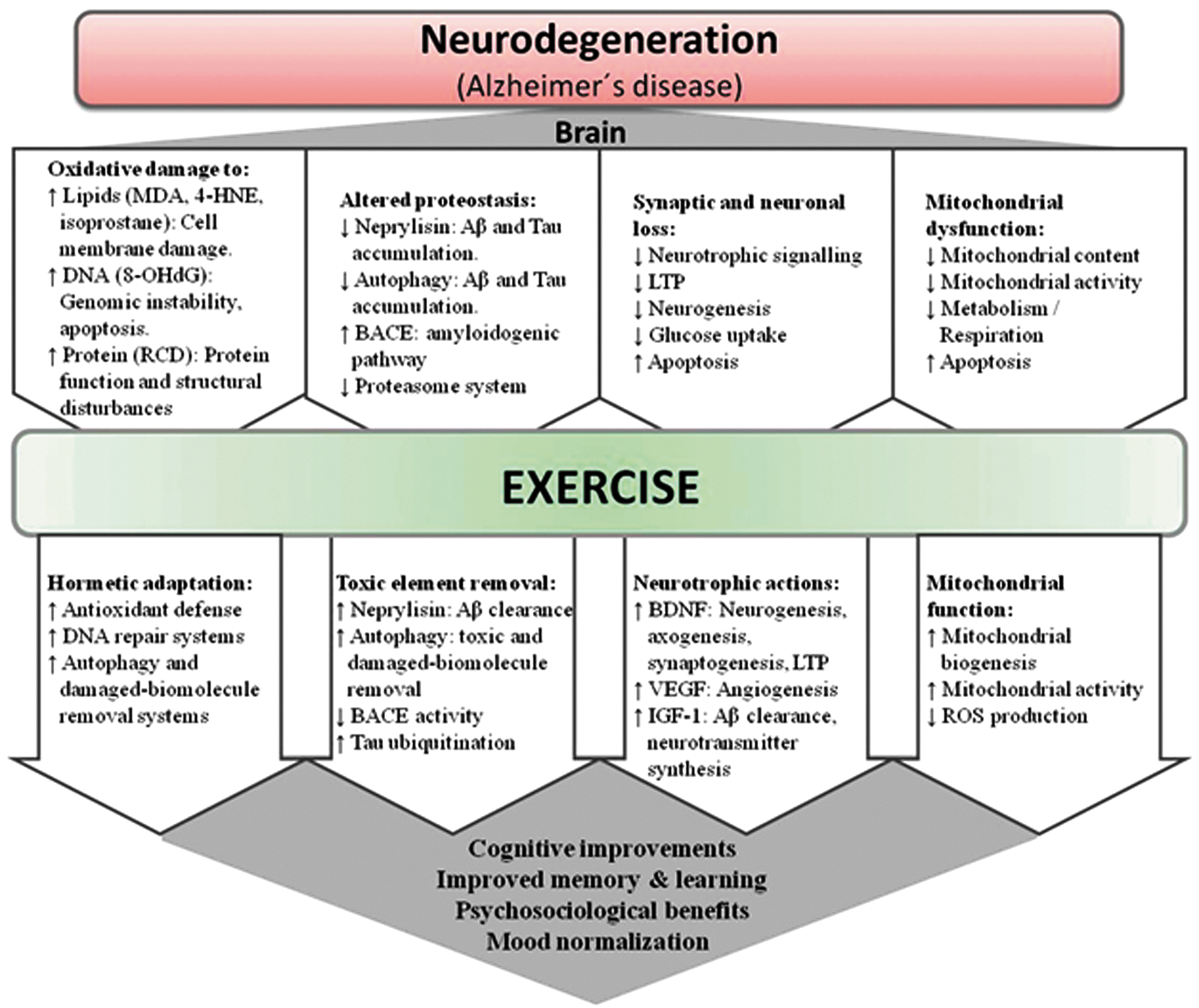
Main signaling pathways involved in the exercise effects in neurodegeneration, especially with regard to Alzheimer's disease. 4-HNE, 4-hydroxynonenal; 8-OHdG, 8-hydroxy-2′-deoxyguanosine; Aβ, amyloid-pβ; BACE, β-secretase; BDNF, brain-derived-neurotrophic factor; IGF-1, insulin-like growth factor 1; LTP, long-term potentiation; MDA, malondialdehyde; RCD, reactive carbonyl derivative; ROS, reactive oxygen species; VEGF, vascular endothelial growth factor. Color images available online at
ACE, angiotensin-converting enzyme; APP, amyloid precursor protein; DMA, asymmetric dimethylarginine; BDNF, brain-derived neurotrophic factor; EGF, epidermal growth factor; EPC, endothelial progenitor cells; ET, endurance training; GDNF, glial-derived neurotrophic factor; IGF-1, insulin like growth factor 1; NGF, nerve growth factor; SDF-1, stromal cell-derived factor-1; ST, strength training; TNFα, tumor necrosis factor-α; VEGF, vascular endothelial growth factor.
Oxidative stress plays an important role in the etiology of Alzheimer's disease. 151 –154 The brain is especially sensitive to oxidative stress compared to other organs owing to its high metabolic rate (i.e., high O2 consumption) together with its relatively low anti-oxidant defense capacity and its high levels of polyunsaturated fatty acids and metals. 151,155 Elevated levels of brain oxidative stress are found with normal aging, 156,157 and this phenomenon is exacerbated in Alzheimer's disease due to additional sources of ROS such as Aβ accumulation and mitochondrial dysfunction. 151,153,154,158 Although there is some controversy, aerobic exercise enhances anti-oxidant defense and mitigates oxidative damage in the brain of rodent models. 150 Thus, exercise increases glutathione peroxidase (GPx) activity in whole brain, 159 as well as superoxide dismutase (SOD) and GPx activity in the brainstem and corpus striatum. 160 Using a triple transgenic mouse model of Alzheimer's disease, we recently showed that aerobic exercise training can increase hippocampal catalase mRNA levels. 161
Exercise attenuates aging neurodegeneration partly by up-regulating neurotrophic factors, such as the brain-derived neurotrophic factor (BDNF). 162,163 Circulating BDNF levels increase with aerobic exercise, especially when intensity is high. 164 –166 Exercise-produced BDNF can help maintain brain function and promote neuroplasticity 167,168 as well as repairing motor neurons. 169 Increased BDNF transcripts in exercised rodents' brains are well documented, which provides a biological explanation for the beneficial effect that exercise has in cognitive function, with tropomyosin receptor kinase (trkB), CREB, or synapsin I signaling been involved. 170 These pathways are involved in synaptogenesis 171 and long-term memory formation. 172 Furthermore, the activation of the transcription factor CREB leads to induction of several genes that regulate neurotrophic effects, including those encoding PGC-1α, 173 dynorphin, and BDNF. 174 In addition, when the exercise induction of the BDNF pathway is blocked, aerobic exercise is unable to activate CREB and stimulate cognition. 170,175 The hippocampal regulation of BDNF induced by exercise is mediated by neurotransmitters, 167,176 neuroendocrine mechanisms, 167 and insulin-like growth factor 1 (IGF-1) modulation. 177,178 Thus, hippocampal IGF-1 levels increase with exercise training 177 and have neurotrophic effects because IGF-1 activates BDNF signaling 177 and increases trkB levels. 179 Aerobic exercise induces other neurotrophic factors, such as vascular endothelial growth factor (VEGF), 180 nerve growth factor (NGF), 181 glial-derived neurotrophic factor (GDNF), 182 neurotrophin (NT)-3, 183 and NT-4/5, 184 all of which act synergistically to induce neurogenesis and neuroplasticity. 167,180
Although neurons are post-mitotic cells, neurogenesis can still occur in specific areas of the adult hippocampus 185 with some stimuli such as ischemia/reperfusion, aging, metabolic pathology, or physical exercise being able to change the rate of neurogenesis. 150 Van Praag and collaborators have extensively studied the effects of exercise, particularly of the aerobic type, on adult neurogenesis 186 and have demonstrated that the newly formed neurons are associated with the cognitive and synaptic effects induced by exercise. 187 This phenomenon is especially important in age-related neurodegeneration because attenuation of accelerated neuronal loss can prevent several age-related disorders, including Alzheimer's disease. Several signaling pathways induced by aerobic exercise have been suggested to mediate this process such as BDNF, 188,189 VEGF, 180 and IGF-1. 190
Both age and disease-induced neurodegeneration are partially produced by a dysregulation of protein homeostasis (see further below on the exercise effects in aging “proteostasis”). Aerobic exercise increases proteolytic degradation by proteasomes and neprilysin, a specific Aβ-degrading enzyme. 167,191 Activation of the brain proteasome is important in preventing Alzheimer's disease because proteasome inhibition produces Aβ accumulation, a hallmark of this disorder. 192 The neurofibrillary tangles produced by an accumulation of hyperphosphorylated tau proteins is also an important hallmark of Alzheimer's disease. Lysine residues of tau are susceptible to ubiquitination, indicating interaction of tau aggregation by oligomerization and ubiquitination-mediated degradation through the proteasome system. 193 The proteasome might also be involved in the learning process, because its inhibition in the hippocampus blocks long-term memory. 194–195
Exercise and the Cellular Hallmarks of Aging
In a recent state-of-the-art review, López-Otín et al. 196 nicely postulated nine hallmarks of aging that might be targeted in future pharmacological interventions—genomic instability, telomere attrition, epigenetic alterations, loss of protein homeostasis (proteostasis), deregulated nutrient sensing, mitochondrial dysfunction, cellular senescence, stem cell exhaustion, and altered intercellular communication. Although more research is needed, exercise, which is available at low cost and largely free of adverse effects, 111 can influence, at least partly, most of these hallmarks (see Fig. 1, right column, for a summary).
Genomic instability
A 5-month aerobic exercise program prevented mitochondrial DNA (mtDNA) instability in multiple tissues of mtDNA mutator (progeroid) mice, thereby reducing multisystem pathology and preventing premature mortality. 197 Oxidative damage to DNA occurs during the aging process. 198 Resistance exercise decreases such damage in old people, as indicated by 8-hydroxy-2′-deoxyguanosine (8-OHdG) determination, through stimulation of endogenous anti-oxidant defense, 199 whereas in rodent models aerobic exercise improves DNA repair mechanisms (e.g., proteasome complex), 200 as well as NF-κB and PGC-1α signaling. 121,201
Telomere attrition and telomerase activity
Accelerated telomere shortening is linked with numerous age-related chronic diseases and risk factors. 202 –210 On the other hand, there is increasing evidence supporting an association between habitual physical exercise, particularly aerobic exercise, and longer leukocyte telomere length. 211 –217 Leukocyte telomere length is also positively associated with cardiorespiratory fitness (expressed as VO2max), 213,218,219 which, in turn, is associated with lower CVD and all-cause mortality. 220 Long-term aerobic exercise can modulate leukocyte telomere length as well as the network of proteins that interact with telomeres, through activation and induction of telomerase enzyme activity (mediated by human telomerase reverse transcriptase [TERT]) and the shelterin complex (or telosome). 217 In effect, TERT mRNA expression is up-regulated in leukocytes after exercise. 221 Exercise also regulates the microRNAs (miRNAs) that control the downstream expression of genes involved in telomere homeostasis. 221 The association between physical exercise and telomere length could also be due to lower oxidative stress and inflammation, exercise-induced regulation of telomeric genes, or a complex interplay between these processes. 221,222 The effects of exercise on skeletal muscle tissue telomeres have been less studied compared with leukocytes, and the results are less conclusive. 223,224
Epigenetic adaptations
The relationship between epigenetics regulation (e.g., DNA methylation) and aging is complex and controversial, i.e., hypomethylation or hypermethylation might be either beneficial or detrimental depending on the different cell types; and, whether manipulations of histone-modifying enzymes can influence aging through purely epigenetic mechanisms remains to be clarified. 196 While keeping in mind the above-mentioned controversy, exercise seems to induce epigenetic modifications that can help attenuate age-deregulations, 225 and several mechanisms, such as metabolic adaptations and transient hypoxia conditions, have been proposed recently. 226 Regular aerobic exercise can modify genome-wide DNA methylation in humans. 227 Animal 216,228,229 and human research 216 suggests that aerobic exercise induces, through epigenetic mechanisms, telomerase activity and the transcription of genes encoding telomere-stabilizing proteins. Both resistance and aerobic exercise can increase DNA methylation, cause histone modifications, and induce miRNAs in a wide range of tissues, including among others, muscle, brain, and cardiovascular system. 225,226 Transient DNA hypomethylation of gene-specific promoter regions precedes increases in mRNA expression in response to acute exercise. 230 In turn, these pulses of elevated mRNA during recovery from acute exercise facilitate protein synthesis and induce gradual structural remodeling and long-term functional adjustments. 231 In general, these adaptations are intrinsic to the working skeletal muscle and collectively contribute to maximize substrate delivery, mitochondrial respiratory capacity, and contractile function during exercise. The net effect is promotion of optimal performance during a future exercise challenge, resulting in a robust defense of homeostasis in the face of metabolic perturbation and, consequently, enhanced resistance to fatigue. 232,233 Several epigenetic mechanisms, including histone H4 deacetylation and loss of promoter methylation, have been implicated in the modified gene expression profile that occurs as an adaptation to aerobic exercise. 234
Epigenetic mechanisms are not restricted to early stages of human development but are broad dynamic controllers of genomic plasticity in response to environmental factors such as exercise. 235 For instance, in young adults, the class II HDACs 4 and 5 (transcriptional repressors) can translocate from the nucleus to the sarcoplasm of muscle fibers in response to aerobic exercise. 108 Over-expression of HDAC5 in transgenic mice blocks the effects of exercise training, further suggesting a contribution of histone modifications in the transcriptomic response to muscle contraction. 236 In human and mouse muscles, methylation of PGC-1α, mitochondrial transcription factor (TFAM), MEF2A, citrate synthase (CS), and pyruvate dehydrogenase kinase isozyme 4 (PDK4) gene promoters decreases after an acute bout of aerobic exercise. 230 The degree of DNA methylation of a large number of genes changes in response to exercise in both skeletal muscles and adipose tissue. 237,238 In addition, aerobic exercise-induced SIRT-1 regulates the tumor suppressor p53, PGC-1α, NF-κB as well as other transcription factors via its deacetylase activity. 225,239
Chronic moderate aerobic exercise increases the methylation levels of the pro-inflammatory apoptosis-associated speck-like protein caspase (ASC) gene, which modulates IL-1β and IL-18 in the leukocytes of old people, thereby contributing to attenuation of age-related increases in pro-inflammatory cytokines. 240 Aerobic exercise training also alters DNA methylation in a chronic manner. 241 Thus, 48 hr after a bout of aerobic exercise, the DNA methylation profile of genes involved in diverse metabolic pathways, as well as in calcium and insulin signaling, was recently found to be differentially methylated in skeletal muscle. 241 A majority of the detected genes in this study were chronically hypomethylated after exercise training. Both aerobic and resistance exercise also can help combating aging sarcopenia and frailty by modulating, through epigenetic mechanisms, several myogenic regulatory factors, e.g., myogenin, myoblast determination protein 1 (MyoD), or myogenic factors 5 (Myf5) and 6 (Myf6, also known as myogenic regulatory factor 4 [Mrf4] or herculin). 242 –244 Exercise also helps to attenuate the aging-related epigenetic deregulation of growth factors in neurodegenerative diseases, not only by up-regulating BDNF induction as mentioned above, but also by promoting remodeling of the chromatin containing the BDNF gene. 245
Overall, the study of exercise-induced epigenetic modifications is just in its infancy. Yet the studies available have already provided new insights into the potential tissue-specific alterations in DNA methylation induced by exercise and into some of the mechanisms explaining the beneficial effects of regular exercise.
Loss of proteostasis
Proteostasis is defined as the protein homeostasis that is responsible for refolding or degrading altered proteins by several mechanisms, such as autophagy, proteasomal degradation, or chaperone-mediated folding. A loss of function in these processes leads to an aggregation of damaged proteins and thereby proteotoxic effects that have been associated with aging 196,246 and age-related conditions such as Alzheimer's or Parkinson's disease. 247
The autophagy–lysosomal and the ubiquitin–proteasome systems, two important proteostatic mechanisms, are impaired by aging. 248,249 Exercise has a beneficial effect in autophagy. 250 In aging mouse models, aerobic exercise induces autophagy in: (1) The brain, supporting its potential to promote elimination of damaging proteins causing neurodegeneration 251 ; (2) heart 252 ; or (3) muscle (besides preventing apoptosis), by modulating IGF-1, Akt/mTOR, and Akt/Forkhead box O3A (FoxO3a) signaling, thereby preventing loss of muscle mass/strength. 253,254 Although data are still scarce in aging humans, autophagy muscle markers are up-regulated after combined exercise training (walking and moderate-intensity leg resistance exercises) in old women. 255
Aerobic exercise induces autophagy in mice through activation of the BCL-2–beclin-1 complex, 256 whereas beclin-1 disruption in transgenic mice reduces autophagy leading to neurodegeneration. 257 Moreover, the aging human brain shows a down-regulation of beclin-1, 258 whereas healthy centenarians have higher serum levels of beclin-1 compared with young controls, suggesting that elevated basal levels of autophagy may be related to healthy human exceptional longevity. 259 MacKenzie et al. showed that acute high-resistance exercise evoked increased muscle protein synthesis and decreased protein degradation in rats, through activation of the class 3 phosphatidylinositol 3OH-kinase (PI3K) Vps34 mVps34, 260 which is known to regulate autophagy by forming a complex with beclin-1. 261 Using an atrogin-1 (also known as MAFbx) knockout mouse model, Zaglia et al. demonstrated that autophagy dysfunction promotes cardiomyopathy and premature death. 262 Atrogin-1 is a muscle-specific ubiquitin ligase involved in muscle atrophy through FoxO signaling. 263 Similar to skeletal muscle, atrogin-1 up-regulation in the heart leads to atrophy. 264 Interestingly, aged atrogin-1 knockout mice have reduced tolerance to treadmill exercise and shortened life span compared with age-matched controls. 262 In this animal model, muscle age-related increases in oxidative damage and apoptosis are attenuated by regular aerobic exercise, whereas both mechanisms are negatively correlated with autophagy. 261
Deregulated nutrient sensing
Exercise exerts protective effects against age declines in the glucose-sensing somatotrophic axis, 265 and also activates at the muscle level the three main interconnected nutrient-sensing systems, i.e., the amino acid–sensing mTOR pathway 266,267 and the low energy–sensing AMPK and SIRT pathways, 132,268 thereby promoting a beneficial muscle anabolic state. On the other hand, exercise improves insulin sensitivity through increased production of the glucose transporter type 4 (Glut 4). 269
In addition, resistance exercise acutely increases the circulating levels of testosterone, growth hormone (GH), and IGF-1, with the magnitude of the effect usually increasing with higher exercise intensity 270 or duration, 271 shorter rest intervals, 272 and higher exercising muscle mass. 273,274 Thus, resistance exercise is a useful approach to prevent sarcopenia by virtue of its ability to increase protein synthesis 275 –277 and skeletal muscle mass. 278,279 However, the rate of muscle protein synthesis in response to exercise training is lower in the elderly than in younger people, 280,281 leading to a lower capacity to improve skeletal muscle strength and fiber size. 282 On the other hand, supplementation with essential amino acids together with resistance training increases muscle protein synthesis both in young and old individuals, although such an effect is also attenuated in the latter, owing, at least partly, to lower ERK1/2 and mTOR signaling. 280 (For a more extensive description about nutrient-sensing modulation by exercise, see above.)
Mitochondrial dysfunction
mtDNA mutations (typically deletions) accumulate with age in different tissues, 283,284 including mainly the nervous and skeletal muscle tissue. 37,285 Mutations in muscle mtDNA play a causal role in the physiological mechanisms implicated in sarcopenia, 37 –40 particularly in abnormalities of the electron transport system, muscle fiber atrophy, and breakage. 37,39,40 Despite classic studies demonstrating exercise-induced mitochondrial biogenesis in young but not in aged mice, 286 recent findings have shown that aerobic exercise training attenuates mitochondrial dysfunction and loss of mitochondrial content in the aging human skeletal muscle while increasing oxidative capacity and the activity of different electron transport chain protein complexes. 287
The accumulated damage to the mitochondria due to the ROS generated from the electron transport chain is the base of the mitochondrial theory of aging first proposed by Harman. 288 Oxidative damage to mtDNA increases with aging, affecting mtDNA replication and transcription, which, in turn, alters the functionality of mitochondrial proteins. 289,290 Lanza et al. demonstrated that age-related decline in mitochondrial oxidative capacity was absent in endurance-trained humans, who showed elevated expression of mitochondrial proteins, mtDNA, and mitochondrial transcription factors. 291 In mtDNA mutator mice, aerobic exercise promoted systemic mitochondrial biogenesis, prevented mtDNA depletion and mutations, increased mitochondrial oxidative capacity and respiratory chain assembly, restored mitochondrial morphology, and blunted pathological levels of apoptosis in multiple tissues. Thus, the authors concluded that a systemic “mitochondrial rejuvenation” occurred as a result of the training program. 197
The lower mitochondrial enzyme activity commonly shown in older compared with younger adults 17,292 is associated with a down-regulation of the mRNAs encoding mitochondrial proteins in skeletal muscle. 293 –295 Aged subjects have cytochrome c oxidase (COX)-deficient muscle fibers, 296 especially in sarcopenic muscles or in those focal regions with higher content of mtDNA mutations. 39,296 –299 Yet resistance exercise training can reverse the muscle transcriptional signature of aging back to that of younger levels for most genes involved in mitochondrial function. 299 Gomes et al. 300 recently provided novel insights into the mechanisms responsible for the age decline of mitochondrial homeostasis by elegantly showing a regulatory pathway that is SIRT1-mediated and independent of PGC-1α and -β, with aging declining NAD+ levels and thereby reducing SIRT1 activity and leading to impaired oxidative phosphorylation (OXPHOS). Short treatment (1 week, or ∼8 months, when translated to the human life span) of 22-month-old mice with nicotinamide mononucleotide (NMN) (a precursor to NAD+ increasing NAD+ levels in vivo) reversed several biochemical indicators of muscle mitochondrial senescence, with increased OXPHOS transcripts in the gastrocnemius muscle. It was proposed that NMN or other compounds able to increase NAD+ are candidates to be included in the human anti-aging armamentarium. And yet NMN, as opposed to exercise, was unable to reverse other age-dependent whole-organism effects, such as loss of muscle strength.
Besides the above-mentioned benefits of resistance exercise on muscle strength until end of life (which were summarized in Table 1), regular exercise has a profound beneficial effect on human mitochondrial function/biogenesis, 301 with this effect being both PGC-1 and SIRT mediated. 111 An active lifestyle attenuates aging mitochondrial dysfunction, promoting longevity through pathways common to the effects of caloric restriction. 291 In addition, some “myokines” (see further below for the definition of myokine) have a mitochondrial rejuvenating effect, e.g., visfatin, a NAD+ biosynthetic enzyme that stimulates the SIRT-1 pathway. 252
Cellular senescence
Cellular senescence is defined as a stable arrest of the cell cycle coupled with stereotyped phenotypic changes, and its regulation during aging is a complex process. Indeed, the same phenomenon, i.e., elimination of senescent cells, that is beneficial to delay age-related pathologies and thus to promote longevity, could also stimulate cancer development. 196 Besides inducing secretion of anti-tumorigenic myokines such as secreted protein acidic and rich in cysteine (SPARC, also known as basement membrane protein [BM]-40), calprotectin, or leukemia inhibitory factor, 302 exercise, mainly aerobic exercise, may decrease cancer incidence and help improve cancer prognosis through several mechanisms, including greater natural killer (NK) cell activity, enhanced antigen presentation, reduced inflammation, and prevention of functional senescent cells' accumulation. 303 Telomere-associated proteins regulate cellular senescence and, as described above, are up-regulated by exercise. Moreover, aerobic exercise increases the aortic expression of telomere repeat-binding factor 2 and Ku70 and reduces the expression of apoptosis regulators, such as cell-cycle-checkpoint kinase 2, p16INK4a, and p53 or survival regulators. 216
Telomere-associated proteins, as well as p16INK4a/Rb and p19ARF/p53 signaling, are considered main pathways in the control of human aging and age-associated pathologies. 196 p16INK4a and p21 are cell cycle inhibitors that are up-regulated in senescent cells. 304,305 p21 is a downstream target of p53 and telomere dysfunction, whereas p16INK4a appears to be up-regulated in a p53- and telomere-independent manner. 306 Sousa-Victor et al. recently highlighted the importance of p16INK4a in the modulation of cellular senescence. In a geriatric mouse model, muscle satellite cells lose their quiescent state owing to deregulation of p16INK4a, whereas repressing p16INK4a restores muscle regenerative capacity. 307 Thus, we suggested the importance of p16INK4a modulation as a new target for combating aging-related chronic diseases. 308 Lifestyle factors, including smoking and physical aerobic exercise practice, have been associated with p16INK4a mRNA levels in peripheral blood T ymphocytes, 309 a biomarker of human aging. 310 Thus, physical exercise is inversely correlated with p16INK4a mRNA levels, i.e., higher amounts of physical exercise leads to down-regulation of p16INK4a in blood cells, which might promote protective effects against age-dependent alterations. 309
As exposed above, cellular senescence plays a key role not only in cancer development
311,312
but also in aging.
313
In fact, cell senescence is one of the major paradigms of aging research through the acquisition of the senescence-associated secretory phenotype (SASP) or senescence-messaging secretome.
314
SASP is a DNA damage response, which, through production of inflammatory, growth-promoting, and remodeling factors can potentially explain how senescent cells alter tissue microenvironments.
315
Different animal model investigations have shown that exercise modulates senescence associated to aging. Thus, 12 weeks of aerobic (swimming) exercise training suppressed liver senescence markers and down-regulated inflammatory mediators by reducing gamma glutamyltranspeptidase activity and levels of p53, p21, and IL-6 in a
Stem cell exhaustion
The decline in the regenerative potential of tissues is a main characteristic of aging, whereas exercise is one of the most potent stimuli for the proliferation/migration of the different adult stem cell subsets from their home tissue (e.g., bone marrow) to target damaged tissues for subsequent engraftment and regeneration. 252 Thus, regular exercise attenuates age-associated reduction in the endothelium-reparative capacity of endothelial progenitor cells. 317 Exercise activates mesenchymal stem cells, which are pluripotent progenitors with a wide variety of therapeutic potential (e.g., as vehicles of anti-cancer genes) and promotes proliferation of neural stem cells, thereby contributing to improve brain regenerative capacity and cognitive ability. 252
Arguably the most affected stem cell type during aging is the myogenic one, known as satellite cells. 318 Although the human skeletal muscle tissue maintains myofiber replacement and repair potential throughout most of life, the efficiency of this process declines with aging, owing to satellite cell alterations. Age-reduced number or functionality of these myogenic cells prevents proper maintenance of muscle mass. 318 –321 Specifically, aging atrophy of type II muscle fibers is accompanied by a specific decline in the content of type II muscle fiber satellite cells. 318 Thus, since sarcopenia is associated with atrophy of type II muscle fibers, its pathophysiological mechanisms are closely related with the decline in satellite cell content with aging. 31 Both aging reductions in muscle mass and strength are positively correlated with muscle fiber type specific cross-sectional area, myonuclear content, and satellite cell content. 322
Animal studies have demonstrated that aerobic exercise increases myofibers that contain higher numbers of satellite cells in both young and old rats, 323 and also promotes expansion of the satellite cell pool in young and old mice. 324 The contribution of these stem cells to skeletal muscle regeneration has been well documented. 325,326 As stated by Hawke and Garry, 326 because adult myofibers are post-mitotic cells, the regulation of skeletal muscle is dependent on a small population of resident cells that are the satellite cells. The regulation of satellite cells involves several mechanisms, including immune response, neurotransmitters, neurotrophic and vascular factors (among other growth factors such as IGF-1 327 ), cytokines such as IL-6, 328 testosterone, or NO•, most of which are modulated by exercise. 111
Not only in young adults 329 but also during aging, resistance training is able to induce skeletal muscle satellite cell proliferation and differentiation, thereby resulting in hypertrophy of type II fibers. 330 The latter phenomenon, in turn, attenuates the pro-sarcopenic physiological events related to type II fiber atrophy associated with aging, 31,318, 322 On the other hand, although resistance training in the elderly of both sexes can counteract the loss of muscle mass and strength, 331 a recent study reported that satellite cell induction in response to a single bout of resistance exercise is delayed in old men. 332 McKay et al. 333 also showed that, compared to young adults, muscle levels of myostatin, a protein that inhibits muscle differentiation and growth in the myogenesis process, were two-fold higher in older individuals, who also had 35% fewer basal stem cells and a type II fiber-specific impairment in stem cell content. The authors concluded that the co-localization of myostatin with satellite cells explains the worsened myogenic capacity of the aged skeletal muscle. 333 In fact, an aging-blunted activation of type II muscle fiber satellite cells in response to an acute bout of resistance exercise was recently shown by Snijders et al. 332 In addition, the satellite cell response to resistance exercise is related to the extent of muscular hypertrophy induced by training. 334
Altered intercellular communication
Aging is associated with altered intercellular communication leading to inflammation or “inflammaging.” 196 Several mechanisms are responsible for this process, including accumulation of pro-inflammatory tissue damage, immune dysfunction, release of pro-inflammatory cytokines by senescent cells, higher activation of NF-κB, or impaired autophagy regulation. 196,335 These events activate the NOD-like receptor protein 3 (NLRP3) “inflammasome,” characterized by elevations in IL-1β, tumor necrosis factor-α (TNF-α), and interferons. 335,336 Interestingly, calorie restriction and exercise-mediated weight loss in obese individuals with type 2 diabetes lead to a reduction in adipose tissue expression of the NLRP3 inflammasome and IL-1β, and thus to reduced inflammation. 337
The decay factor AUF1 (AU-binding factor 1, also known as heterogeneous nuclear ribonucleoprotein D or hnRNP D) is implicated in the cessation of the inflammatory response (by mediating cytokine mRNA degradation) and also in the maintenance of telomere length by modulating TERT. 338 Down-regulation of AUF1 leads to accelerated cellular senescence and premature aging in mice, which is rescued by normalizing the expression of this factor. 196 Lai et al. found that chronic muscle contractile activity increased different AUF1 isoforms (p37, p40, and p45) in the muscle of healthy rats, resulting in improved muscle plasticity in response to subsequent contractile activity. 339 Senescent cells transmit their condition to other cells through multiple mechanisms, including ROS, growth factors, and interleukins. 340 As mentioned above, chronic physical exercise (mostly of the aerobic type) decreases ROS damage, and it does so by decreasing ROS production at the mitochondrial level while up-regulating endogenous anti-oxidant defense. 121
Importantly, skeletal muscle fibers produce hundreds of secreted factors or “myokines” (including the above-mentioned neurotrophins) with a potential drug-like effect at the local and systemic levels, i.e., proteins, growth factors, cytokines, or metallopeptidases, and this secretory capacity increases during and after exercise training (see Fiuza-Luces 252 for an in-depth review and Table 3 for some illustrative examples). Systemic low-level inflammation and related conditions such as CVD or cancer can be attenuated by the cumulative effect of regular exercise bouts, during which the muscle can release IL-6, arguably the myokine prototype. 341 This, in turn, creates a healthy milieu by inducing the production of the anti-inflammatory cytokines IL-1 receptor antagonist (IL-1Ra), IL-10, or TNF soluble receptors (sTNF-R) while inhibiting the pro-inflammatory cytokine TNF-α. 341 The release of IL-6 from working muscles increases with exercise intensity 342 and duration, 343 and endogenous NO• and the interaction between Ca2+/nuclear factor of activated T cell (NFAT) and glycogen/p38 MAPK are the proposed upstream signals leading to its secretion. 344 Other anti-inflammatory myokines include IL-4, IL-10, or IL-13. 345,346 Thus, life-long aerobic exercise training is associated with lower inflammation levels. 347 –349 Higher levels of aerobic exercise have also been associated with lower levels of C-reactive protein (CRP), IL-6, and TNF-α in people aged 70–79 years. 350 However, although exercise training is known to have beneficial anti-inflammatory effects across a broad spectrum of organs and systems, more research is needed in the elderly, particularly to determine if the molecular mechanisms and pathways involved are similar in old people compared with younger population segments. 351
The information provided in the table is based (and adapted from) a previous review paper by the authors. 111
AMPK AMP-activated protein kinase; NAD+, oxidized nicotinamide adenine dinucleotide.
Perspective
The benefits of regular exercise are such that a dose–response is usually observed in humans. Higher levels of moderate-to-vigorous exercise (≥450 min/week, clearly above the minimum international recommendations of 150 min/week) are associated with longer life expectancy. 352 Furthermore, elite athletes—those humans sustaining the highest possible exercise levels (e.g., former participants in the Tour de France cycling race or former Olympic marathoners)—usually live considerably longer than the general population. 353
Physical exercise has a profound effect on the expression of a substantial proportion of our genome, which has evolved to optimize aerobic metabolism in an environment of food scarcity. Thus, physical inactivity is becoming a major public health problem worldwide.
354
Exercise certainly cannot reverse the aging process, but it does attenuate many of its deleterious systemic and cellular effects. Most common age-associated chronic conditions are diseases of physiology and thus physiological interventions, of which physical exercise is arguably the best example, are largely the answer.
355
We propose that more research efforts should be devoted to gain insights into the molecular mediators of the exercise benefits. Besides putting more anti-aging drugs on the market, it would be wise to determine which are the most effective combinations, types, and dosages (frequency, duration, intensity) of exercise for older people and to implement efficient exercise interventions for this and younger population segments.
Footnotes
Acknowledgments
Research in the field of exercise and health by A. Lucia is funded by Fondo de Investigaciones Sanitarias (FIS, grant no. PI12/00914).
Author Disclosure Statement
No competing financial interests exist.
All co-authors fully reviewed the entire manuscript and participated in at least some of its parts, including literature search (all authors), figure design (F.S.-G., N.G., H.P.-G.), tables edition (A.S.-L., C.F.-L., N.G.), or writing and editing of a significant part of the manuscript (all authors).