
Abstract
Reducing sugars can react non-enzymatically with the amino groups of proteins, lipids, and nucleic acids to initiate a complex series of rearrangements and dehydrations, and then to produce a class of irreversibly cross-linked heterogeneous fluorescent moieties, termed advanced glycation end products (AGEs). The formation and accumulation of AGEs in numerous tissues have been known to progress in a normal aging process and at an accelerated rate under hyperglycemic and/or oxidative stress conditions. There is a growing body of evidence that interaction of AGEs with a cell-surface receptor termed receptor for AGEs (RAGE) elicits oxidative stress generation and subsequently evokes proliferative, angiogenic, and inflammatory reactions, thereby being involved in the development and progression of various types of cancers. These observations suggest that accumulation of AGEs and resultant activation of the RAGE signaling pathway could partly explain the increased risk of a variety of cancers in patients with diabetes or in elderly subjects. This article summarizes the pathological role of RAGE activation by AGEs and other ligands in tumor growth and metastasis and its therapeutic interventions for the life-threatening disorders.
Introduction
E
A non-enzymatic reaction between reducing sugars and the amino groups of proteins, lipids and nucleic acids, which is also known as the Maillard reaction, has been known to progress in a normal aging process and at an accelerated rate under hyperglycemic and/or oxidative stress conditions, thus leading to the formation and accumulation of advanced glycation end products (AGEs). 6 –13 There is an accumulating body of evidence that interaction of AGEs with a cell-surface receptor termed receptor of AGEs (RAGE) elicits oxidative stress generation and subsequently evokes proliferative, angiogenic, and inflammatory reactions, thereby being involved in the development and progression of various age- or diabetes-related disorders, including cardiovascular disease, Alzheimer's disease, osteoporosis, cancer growth, and metastasis. 14 –31 Moreover, oxidative stress generation promotes the formation of AGEs and as a resultant increases RAGE expression, thereby forming a positive feedback loop between AGEs accumulation and the RAGE downstream pathway. 32
These observations suggest that blockade of the crosstalk between the AGEs–RAGE system and oxidative stress generation might be a therapeutic target for preventing and slowing the development and progression of various types of cancers, especially in patients with diabetes or elderly subjects. However, methylglyoxal, one of the most powerful glycating agents of proteins and precursors of AGEs, could retain its efficacy as an anti-cancer drug, and overexpression of glyoxalase I, an enzyme that mainly detoxifies methylglyoxal, was associated with multidrug resistance in cancer chemotherapy. 33 –35 Therefore, it should be noted that there is some controversy about the role of methylglyoxal-derived AGEs in cancer development and progression.
This article summarizes the pathological role of RAGE activation by AGEs, carboxymethyllysine (CML), and other ligands in tumor growth and metastasis and therapeutic interventions for life-threatening disorders. In this review, literature searches were undertaken in Medline by the PubMed interface. Non-English language articles were excluded. Key words (RAGE, glycation, cancer) have been used to select the articles.
Molecular Mechanisms for AGEs–RAGE-Evoked Tumor Growth and Metastasis
Previously, we have shown that AGEs–RAGE interaction-mediated, nicotinamide adenine dinucleotide phosphate (NADPH) oxidase–induced reactive oxygen species (ROS) generation increases vascular permeability and stimulates proliferation and tube formation of microvascular endothelial cells (ECs), the key steps of tumor angiogenesis, through vascular endothelial growth factor (VEGF) expression via transcriptional activation of nuclear factor-κB (NF-κB) and activator protein-1 (AP-1). 36 –40 Furthermore, activation of the AGEs–RAGE axis also evokes inflammatory reactions in ECs via ROS generation. 41 –45 Indeed, AGEs have been reported to increase monocyte chemoattractant protein-1 (MCP-1) and intercellular cell adhesion molecule-1 (ICAM-1) expression in microvascular ECs through intracellular ROS generation through the interaction with RAGE, thereby inducing leukocyte adhesion to ECs. Several lines of evidence implicate VEGF as the key factor involved in tumor growth and metastasis. 46 VEGF expression levels are associated with angiogenesis and macrophage infiltration, the extent of which is correlated with various tumor prognoses. 47 –51 Therefore, the AGEs–RAGE interaction could stimulate tumor-associated angiogenesis and macrophage infiltration by inducing VEGF expression and its related inflammatory reactions, thereby promoting tumor growth and metastasis.
AGEs inhibit prostacyclin production and induce plasminogen activator inhibitor-1 in microvascular ECs through an interaction with RAGE as well. 52 –54 These observations suggest that AGEs have the ability to cause platelet aggregation and fibrin stabilization, resulting in a predisposition to thrombogenesis and thereby contributing to capillary obliteration. 52 –54 The resultant ischemia and hypoxia within the tumors might trigger VEGF expression again, thus further augmenting tumor angiogenesis and expansion.
AGEs–RAGE interaction increases hypoxia-inducible factor-1α (HIF-α) transcriptional activity and its protein expression in ECs and renal cells. 55 –57 Conversely, hypoxia stimulates brisk generation of AGEs by ECs and subsequently activates the RAGE downstream pathway, which could trigger up-regulation of early growth response-1, thereby causing inflammatory and thrombotic reactions within the hypoxic areas of tumors. 58 In addition, hypoxic exposure stimulated neuronal RAGE gene expression via the transcriptional factor HIF-1, and hypoxia was associated with the increased expression of RAGE in human prostate and breast tumors. 59,60 Therefore, the positive feedback loop between the AGEs–RAGE axis and hypoxia might also potentiate tumor-associated angiogenesis and inflammation, being involved in tumor growth and expansion.
Recently, engagement of RAGE with AGEs has been shown to stimulate both expression and activation of carbohydrate-responsive element-binding protein (ChREBP), a key transcription factor that could promote glycolytic and anabolic reactions, thus enhancing proliferation of colorectal and liver cancer cells. 61 Interaction of the RAGE cytoplasmic domain with diaphanous-1 is required for ligand-stimulated cellular migration through activation of Rac1 and Cdc42. 62 Moreover, dedicator of cytokinesis protein 7 is a critical regulator of the RAGE–Cdc42 signaling axis that induces formation of dendritic pseudopodia in human cancer cells. 63 These findings suggest the active involvement of AGEs–RAGE axis in tumor migration and metastasis as well.
Role of the AGEs–RAGE Axis in Development and Progression of Various Cancers
Breast cancer
Recently, we have found that AGEs increase viable cell number and up-regulate mRNA levels of RAGE and VEGF in MCF-7 cells, human breast cancer cells. 64 Neutralizing RAGE antibodies blocked the AGEs-induced increase in viable cell number, whereas metformin completely suppressed the AGEs-induced proliferation as well as up-regulation of RAGE and VEGF mRNA levels in MCF-7 cells. Furthermore, an inhibitor of adenosine monophosphate (AMP)-activated protein kinase (AMPK), Compound C, significantly inhibited these beneficial effects of metformin on AGEs-exposed MCF-7 cells. Because RAGE mediates various biological actions of AGEs in numerous cell types, 65 –67 the observations suggest that metformin could inhibit the AGEs-induced growth and VEGF expression in MCF-7 breast cancer cells by suppressing RAGE gene expression via the AMPK pathway.
Tumors cannot grow beyond a volume of 1–2 mm3 without establishing a vascular supply. 51 Angiogenesis, a process by which new vascular networks are formed from pre-existing capillaries, is required for tumors to grow, invade, and metastasize. 51 VEGF, a specific mitogen to ECs and also known as a vascular permeability factor, is a crucial growth factor for tumor angiogenesis. 51,68,69 There is a growing body of evidence that the VEGF expression level is associated with breast cancer growth and metastasis. 68,69 Therefore, our study provides novel beneficial aspects of metformin on breast cancer expansion; one is inhibition of tumor cell proliferation, and the other is the suppression of VEGF expression, which would lead to inhibition of tumor angiogenesis.
In the largest cohort study (n=588,321), breast cancer mortality was modestly increased in type 2 diabetic women compared with non-diabetic females, and the positive relationship between breast cancer mortality and type 2 diabetes remained significant after adjusting for age, physical activity and BMI, well-known risk factors for breast cancers. 2,70,71 Moreover, metformin use, compared with placebo or other anti-diabetic drugs, has been shown to decrease the risk of cancer incidence and mortality, including breast cancers in type 2 diabetic patients. 72 –75 Therefore, the association of metformin use with the reduced risk for breast cancer incidence and mortality in type 2 diabetic patients 72 –75 may be partly ascribed to its inhibitory actions on the AGEs–RAGE axis in breast cancer cells. RAGE expression levels are correlated to the degree of severity of breast cancer, and small interfering RNAs (siRNAs) raised against RAGE induced G1 arrest and inhibited proliferation of breast cancer cells. 76 Furthermore, knockdown of RAGE has been reported to attenuate 17α-ethinyl-estradiol–dependent proliferation and survival of MCF-7 breast cancer cells. 77 These findings further support the active involvement of RAGE and RAGE ligands in breast cancer growth and expansion.
However, there is still some controversy about the pathological role of AGEs in breast cancers. 78 –80 Triple-negative breast malignant tumors characterized by high proliferation rate, high histologic grade, tumor necrosis, and poor prognosis, exhibited low accumulation of methyglyoxal-derived AGEs compared with other breast cancer subtypes. 78 Inhibition of glyoxalase I, a cellular defense enzyme involved in the detoxification of methylglyoxal, induced apoptotic cell death of irradiated MCF-7 breast cancer cells. 79 Moreover, tamoxifen-resistant MCF-7 breast cancer cells were more sensitive toward toxicity of dicarbonyls, such as methylglyoxal. 80 These observations suggest that methylglyoxal-derived AGEs might protect against the progression of breast cancers by inducing apoptosis.
Melanoma
Among various cancers, malignant melanoma is one of the most highly invasive and metastatic tumors, with an incidence and mortality that have been rapidly increasing in recent years. 81 –84 Although therapeutic advances in the treatment of malignant melanoma have been made with anti-cancer drugs and immunotherapy and that the survival rate of patients with melanoma has improved substantially, these therapeutic options may be limited by considerable side effects. 81 –84 So, to develop a novel therapeutic strategy that could block the growth and metastasis of melanoma is one important clue to improving the prognosis of patients.
Previously, we have shown that AGEs stimulate the growth and migration of human melanoma cells. 85 Moreover, tumor formation of melanoma cell xenografts in athymic mice was prevented by neutralizing antibodies raised against RAGE. 85 In tumor-bearing mice, survival rates were prolonged, and treatment with RAGE antibodies inhibited spontaneous pulmonary metastases of melanoma. In addition, AGEs existed in the beds of human melanoma tumors, whereas they were barely detected in normal skin. These findings demonstrate that AGEs could play a role in growth and invasion of melanoma through interaction with RAGE.
Aptamers are short, single-stranded DNA or RNA molecules that can bind with high affinity and specificity to a wide range of target proteins. 86 Numerous aptamers have been developed and used in the clinical fields as a tool for modulating the function of various proteins. 87,88 Pegaptanib, an RNA aptamer directed against the VEGF165 isoform, has been shown to be effective in treating choroidal neovascularization in patients with age-related macular degeneration. 87 Recently, we have found that a high-affinity DNA aptamer directed against AGEs (AGEs-aptamer) blocks the progression of diabetic nephropathy in an animal model of obese and type 2 diabetes. 89 In addition, AGEs-aptamer significantly inhibited the in vivo tumor growth of G361 melanoma cells. 90 Immunohistochemical and western blot analyses of G361 melanoma cells revealed that AGEs-aptamer decreased expression levels of proliferating nuclear antigen, the number of tumor-associated vessels, and CD31 and Mac-3, markers of endothelial cells and macrophages, respectively. 90 RAGE and VEGF expression levels were also reduced in AGEs-aptamer–treated G361 melanoma. 90 Furthermore, AGEs-aptamer inhibited the AGEs-induced proliferation and tube formation of ECs as well as growth of G361 cells in vitro. These findings suggest that AGEs-aptamer could inhibit the AGE–RAGE axis in G361 melanoma and suppress tumor growth in nude mice by blocking tumor-associated angiogenesis and macrophage infiltration. Blockade of the AGEs–RAGE axis by AGEs-aptamer might represent a promising therapeutic target for malignant melanoma.
RAGE is a multi-ligand receptor, and among its ligands are the calcium-binding S100 proteins and high mobility group box 1 (HMGB1) protein. 91 –96 Compared to normal skin tissue, S100B protein was over-expressed in human melanoma tumor samples. 91 Furthermore, S100P and RAGE expression levels were increased in melanomas than in nevus pigmentosus, especially in metastatic melanomas. 92 There was a positive correlation between expression levels of S100P and RAGE in melanoma tissues. 92 In addition, HMGB1 protein levels were also elevated in human melanomas and co-localized with RAGE. 93 Given that the interaction of S100 proteins and HMGB1 with RAGE plays a role in tumor angiogenesis, growth, invasion, and metastasis, 94 –96 the observations suggest that S100 proteins and HMGB1 protein levels might be a reliable prognostic biomarker in patients with malignant melanoma.
RAGE clustered at membrane ruffles and leading edges and at sites of cell-to-cell contact in primary melanoma cells. 97 RAGE overexpression conferred a metastatic phenotype to the WM115 human primary melanoma cell line, 98 further suggesting the involvement of RAGE in metastasis of melanoma.
Pancreatic cancer
Previously, we have shown that AGEs increase growth of and up-regulate platelet-derived growth factor-B (PDGF-B) mRNA levels in Mia PaCa-2 cells, a human pancreatic cancer cell line. 99 Neutralizing antibodies raised against PDGF-B inhibited the AGEs-induced increase in DNA synthesis of Mia Paca-2 cells. Furthermore, antisense DNA complementary to mRNA encoding RAGE were found to reverse the AGEs-induced PDGF-B gene induction and cell proliferation of pancreatic cancer cells. 99 These observations indicate that AGEs could stimulate proliferation of pancreatic cancer through the interaction with RAGE via autocrine induction of PDGF-B.
Knockdown of RAGE led to increased apoptosis, diminished autophagy, and decreased survival of pancreatic cancer cells. 100 In contrast, over-expression of RAGE was associated with enhanced autophagy, diminished apoptosis, and greater tumor cell viability. RAGE limited apoptosis of pancreatic cancer cells through a p53-dependent mitochondrial pathway. Furthermore, most forms of tumor cell death resulted in release of the RAGE ligand HMGB1. 100 RAGE was required for the activation of interleukin-6–mediated mitochondrial signal transducers and activators of transcription 3 signaling in pancreatic carcinogenesis. 101 In addition, RAGE-elicited activation of autophagy, a programmed cell survival system, has been shown to promote pancreatic cancer growth as well. 102 These findings suggest that a RAGE and HMGB1 interaction could be a crucial inflammatory mediator that modulates the crosstalk between pro-survival pathways and autophagy, and thereby is involved in the development and progression of pancreatic cancer.
Engagement of RAGE with S100 proteins has been shown to stimulate degradation of the extracellular matrix and metastasis, and thus is involved in pancreatic cancer progression and metastasis. 103 S100A11 and S100P levels were increased in pancreatic cancers and could predict poor prognosis of patients with pancreatic cancer. 103
Jiao et al. reported that CML levels were not associated with pancreatic cancer in their prospective study. 104 Moreover, a case–control study within the prospective European Prospective Investigation into Cancer and Nutrition (EPIC) cohort also revealed that there was no association of higher CML levels with risk of pancreatic cancer. 105
Hepatocellular carcinoma
RAGE was expressed in the human hepatocellular carcinoma (HCC) cell lines, Hep3B and HepG2 cells, whereas AGEs increased VEGF expression in these cell types. 106 Furthermore, AGEs-treated conditioned medium significantly increased proliferation, migration, and tube formation of ECs, thus suggesting that AGEs–RAGE signaling could enhance the angiogenic potential of HCC cells by up-regulating VEGF expression. 106 AGEs have been shown to increase the growth of HuH7 cells, another type of HCC cell line, as well. 107 MK615, an extract from Japanese apricot, was reported to inhibit the AGEs-induced proliferation of HuH7 by suppressing RAGE expression. 107
The HCC cell line resistant to hypoxia was found to have higher levels of RAGE expression, and RAGE overexpression showed significantly prolonged survival under hypoxia. 108 Furthermore, cytoplasmic expression of RAGE was correlated with poorly differentiated HCC, and RAGE was an independent predictor for both overall survival and disease-free survival in patients with HCC after hepatectomy. 109 HMGB1 was released under hypoxic stress, which could induce an inflammatory response to promote invasion and metastasis in HCC. 110 Moreover, we have very recently found that serum levels of AGEs are significantly higher in non-B or non-C HCC (NBNC-HCC) patients compared with non-alcoholic steatohepatitis (NASH) and control subjects (9.1±2.7, 5.2±1.7, and 3.5±1.2 [U/mL], respectively, p<0.05). 111 By the use of multiple stepwise regression analysis, age, γ-glutamyl transpeptidase, and high-density lipoprotein cholesterol (inversely) remained significant and were independently related to AGEs levels (R 2 =0.406). These observations suggest that AGEs might be involved in the pathogenesis of NBNC-HCC, thereby being a biomarker that could discriminate NBNC-HCC from NASH. However, Moy et al. have reported that CML levels were inversely associated with liver câncer. 112
Colorectal cancer
RAGE expression was detected in the human colon adenocarcinoma cell line Caco-2, and food-derived casein-linked AGEs induced activation of p44/42 mitogen-activated protein kinases in this cell type, thus suggesting that dietary AGEs intake and subsequent RAGE activation could promote the growth of colorectal cancer. 113 Furthermore, rats treated with high-linoleic acid and high-glucose diets showed the most pronounced multiplicity of aberrant crypt foci and carcinoma associated with increased mucosal AGEs and RAGE expression, compared with control or each diet alone group. 114 In addition, RAGE signaling has been shown to play a critical role in sporadic development of intestinal adenomas and hepatic tumor growth of colorectal cancer as well. 115,116 Very recently, we have found that circulating AGEs levels show a strong positive association with the increased risk of rectal cancer (odds ratio=1.9, n=396) in the nested case–control EPIC study (unpublished data). However, Jiao et al. reported that the highest quintile of CML levels was not associated with an increased risk of colorectal cancer. 117
Lung cancers
CML was detected in tumor cells and tumor-surrounding stroma of lung squamous cell carcinomas and adenocarcinomas, whereas argpyrimidine was predominantly found in tumor cells of lung squamous cell carcinomas. 118 Heat shock protein 27 (Hsp27) was identified as a major argpyrimidine-modified protein, and anti-apoptotic activity of argpyrimidine-modified Hsp27 in squamous cell carcinomas was significantly higher than that of adenocarcinomas. 118 These findings suggest the involvement of high levels of argpyrimidine-modified Hsp27 in the progression of lung squamous cell carcinomas. Furthermore, we have found that AGEs enhance the migration capacity of lung cancer A549 cells by activating Rac1 via ROS generation and increase their invasion capacity as well. 119 However, there is some controversy regarding the pathological role of AGEs in lung cancer. Non–small cell lung carcinoma patients with high AGEs-related plasma fluorescence were characterized by a later reoccurrence of the tumor after curative surgery and a higher survival rate compared with patients with low plasma fluorescence. 120 Mice fed with an AGEs-rich diet developed smaller tumors than mice with a normal diet, thus suggesting that AGEs might be protective against the progression of lung cancer. 120
Other cancers
AGEs–RAGE interaction induced in vitro cell growth of renal cell carcinoma (RCC) and promoted the production of interleukin-6, an autocrine growth factor of RCC. 121
RAGE was associated with angiogenesis in human oral squamous cell carcinoma. 122 RAGE expression could predict recurrence in human oral squamous cell carcinoma. 123 RAGE antisense DNA treatment suppressed the invasive activity of RAGE-positive MKN28 cells, gastric cancer cells. 124 RAGE was expressed preferentially in poorly differentiated adenocarcinomas, and strong RAGE was also associated with depth of invasion and lymph node metastasis. 124 Knockdown of RAGE reduced proliferation and invasion of gastric cancer cells and induced cell apoptosis and cycle arrest. 125 Targeting RAGE expression has been reported to induce apoptosis and inhibit prostate tumor growth in nude mice. 126 Furthermore, expression of RAGE and HMGB1 and their co-expression were all associated with advanced tumor clinical stages of prostate cancer. 127 Blockade of RAGE–HMGB1 decreased growth and metastases of gliomas in mice. 128 These observations suggest that RAGE expression might be closely associated with invasion and metastasis in human oral squamous cell carcinoma, gastric cancer, and prostate cancer, suggesting the pathological role of RAGE and its ligands in growth and expansion of these tumors.
Conclusions
As discussed here, there is accumulating evidence that the AGEs–RAGE axis could play a role in the development and progression of various types of cancers (Fig. 1). AGEs directly stimulated proliferation of cancer cells and inhibited their apoptotic cell death. Furthermore, AGEs and other ligands induced tumor growth and metastasis partly by enhancing angiogenic and pro-inflammatory reactions. All of these deleterious effects were largely mediated by interaction with RAGEs. So, inhibition of the RAGE axis in tumor cells and tumor-surrounding stroma might be a therapeutic target for slowing cancer growth and metastasis. Further basic and clinical studies will provide us with clue for developing a novel therapeutic strategy for cancers.
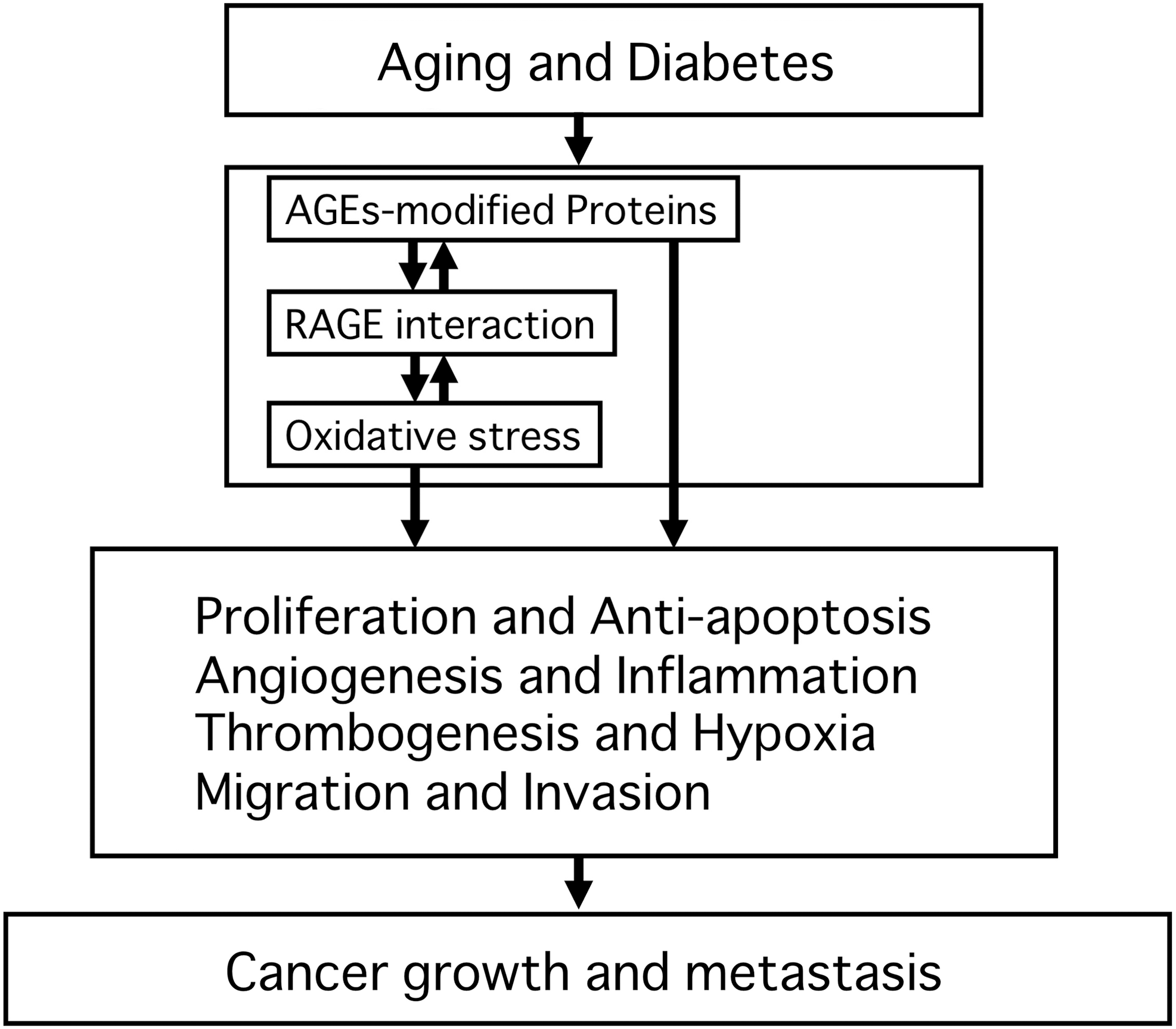
Role of advanced glycation end products and receptor for advanced glycation end products–receptor for advanced glycation end products (AGEs–RAGE) axis in cancer growth and metastasis.
Footnotes
Acknowledgments
This study was supported in part by Grants-in-Aid for Scientific Research (B) from the Ministry of Education, Culture, Sports, Science and Technology, Japan (S. Yamagishi).
Author Disclosure Statement
No competing financial interests exist.