
Abstract
6-Hydroxydopamine (6-OHDA)- and 1-methyl-4-phenylpyridinium (MPP+)-induced hemi-parkinsonism was investigated in relation to the severity of the disorder in terms of behavioral disability and nigral neuronal loss and recovery regarding the number of stem cell–derived neurons transplanted in the striatum. Intra-median forebrain bundle infusion of the parkinsonian neurotoxins and intra-striatal transplantation of differentiated embryonic stem cells (ESCs) were carried out by rat brain stereotaxic surgery. The severity of the disease was determined using the number of amphetamine- or apomorphine-induced rotations, striatal dopamine levels as estimated by high-performance liquid chromatography (HPLC)-electrochemistry, and the number of surviving tyrosine hydroxylase immunoreactive dopaminergic neurons in the substantia nigra pars compacta. Rats that received unilateral infusion of 6-OHDA or MPP+ responded with dose-dependent, unilateral bias in turning behavior when amphetamine or apomorphine was administered. Rotational asymmetry in both models correlated significantly well with the loss in the number of nigral dopaminergic neurons and striatal dopamine depletion. Transplantation of 2×105 differentiated murine ESCs revealed remarkably similar kinds of recovery in both animal models. The survival of the grafted dopaminergic cells in the striatum was better in animals with low-severity parkinsonism, but poor in the animals with severe parkinsonism. Amphetamine-induced rotational recovery correlated positively with an increasing number of cells transplanted in animals with uniform nigral neuronal lesion. These results suggest that disease severity is an important factor for determining the number of cells to be transplanted in parkinsonian rats for desirable recovery, which may be true in clinical conditions too.
Introduction
T
Clinical trials in patients have indicated an important role of severity of the disease for transplantation, because patients with lesions restricted to the dorsal striatum are more likely to experience better recovery. 11 Graft-induced dyskinesias, a known problem of transplantation therapy, are caused by transplantation of an inappropriate number of cells that result in large numbers of surviving cells in the graft. 12 The transplant cell number also has an effect on the survival of cells; grafting of more cells than required may limit the survival of the desired cell type and also reduce the efficacy of behavioral improvement. 13 Patients suffering from PD experience various movement and behavioral abnormalities like bradykinesia or slowness in movement, resting tremor, rigidity or a resistance to passive movements, and postural abnormalities, 14 a measure of which is quantified by the Unified Parkinson's Disease Rating Scale. 15 Similarly, increasing doses of parkinsonian neurotoxins can cause a proportional reduction in striatal DA levels, thereby modeling PD with increased severity. 14,16 –18 There are several behavioral tests for animal models of PD that display the onset and progression of the disease. 19,20 Some of the well-established behavioral tests in unilaterally lesioned animal models of PD include amphetamine- or apomorphine-induced unilateral rotations, cylinder test, elevated body swing test, skilled-forelimb use test, beam balancing test, etc. 21 –23 Amphetamine- or apomorphine-induced rotatory response is used extensively by others and us for validating the development of the disease in hemi-parkinsonian models. It is also one of the consistent tests for evaluation of transplantation recovery in PD. 24 –27 A good correlation between the drug-induced rotations and the severity of the disease was established in intra-nigral 6-hydroxydopamine (6-OHDA)-infused rats. 28
We investigated correlation, if any, between dopaminergic cell loss in the substantia nigra pars compacta (SNpc) and unilateral bias in turning to find if the rotational severity, nigral dopaminergic cell loss, and striatal DA content, singularly or together, could be set as a scale for the severity of or stage the disease in rats unilaterally lesioned with 6-OHDA or 1-methyl-4-phenyl pyrinidium (MPP+). A change from negative to positive correlation of nigral neuronal loss or striatal denervation with that of the performance in behavioral tests provides a scale for the assessment of functional recovery in cell transplantation studies. 29 We show here that transplantation recovery in PD is directly related to the number of cells transplanted, which in turn is associated with disease severity.
Materials and Methods
Animals
Adult male Sprague–Dawley rats (250–300 grams) were used. Experimental protocols met the national guidelines on proper care and use of animals in laboratory research (Committee for the Purpose of Control and Supervision of Experiments on Animals, Ministry of Environment and Forests, Government of India). The number of animals used in each experiment is provided in Table 1.
The study was approved by institutional animal ethics committee (IAEC) appointed by CPCSEA, Animal Welfare Division, Ministry of Environment & Forests, Government of India. The number of animals used in intra-MFB infusion of 6-OHDA/MPP+animals is provided one after the other, respectively. Contralateral sides that received vehicle served as a control, when measuring striatal DA content and SNpc neuronal counts.
6-OHDA, 6-hydroxydopamine; MPP+, 1-methyl-4-phenyl pyridinium; TH-IC, tyrosine hydroxylase immunohistochemistry; SNpc, substantia nigra pars compacta; DA, dopamine; 7d, mESCs differentiated for 7 days; mESCs, murine embryonic stem cells; MFB, median forebrain bundle.
Intra-median forebrain bundle infusion of 6-OHDA and MPP+
Rats anesthetized with chloral hydrate (325 mg/kg, intra-peritoneal [i.p.]) were held over a stereotaxic frame (Steolting, IL). Saline, MPP+, or 6-OHDA (8, 16, or 32 nmol in 1 μL) was infused (0.2 μL/min) into the right intra-median forebrain bundle (MFB) employing a Worker Bee and Syringe Pump (Bioanalytical Systems, West Lafayette, IN). The rat brain stereotaxic coordinates used were anterio-posterior (AP) −0.28 from the Bregma point, lateral (L) 0.15, and dorso-ventral (DV) −0.82. 30 The neurotoxin infusion day is assigned as day 0. On 14th and 16th days, rats were assayed for amphetamine- and apomorphine-induced rotations, respectively. Animals exhibiting three or more rotations per minute were included in the study. Rats were transplanted with given number of cells on 18th day.
Amphetamine- or apomorphine-induced rotations
Rats were placed in Perspex, transparent cylindrical cages (45 cm diameter, 40 cm high). Rotations were counted by persons who were trained in the behavioral assessments, but were blinded to the treatments administered. The number of circling was counted for all the animals following
Intra-striatal transplantation of differentiated cells
Mouse embryonic stem cells (mESCs) were differentiated in serum-free medium for 7 days, as detailed by Tripathy et al. 32 In short, D3 mESCs (American Type Culture Collection [ATCC] no. CRL11632; Manassas, VA) maintained in Dulbecco's modified Eagle Medium (DMEM; Gibco–Invitrogen) supplemented with 15% fetal bovine serum (FBS; Hyclone, Logan, UT), 2 mM glutamine (Gibco–Invitrogen), and 1000 Units of non-essential amino acids, 100 units of penicillin and 100 mg streptomycin (Gibco–Invitrogen), 0.7% β-mercaptoethanol (MP Biomedicals, Solon, OH), and 1000 units of leukemia inhibitory factor (LIF; Millipore, Billerica, MA). For differentiation (day 0), a specific number of cells was plated in culture flasks in DMEM (Gibco–Invitrogen) supplemented with 10% FBS, other constituents being the same as in routine culture. After 48 hr (day 2), medium was changed to knockout-DMEM supplemented with 10% knockout serum replacement (Gibco–Invitrogen) along with all the other components. On the day 7, the cells were scraped into the same medium and centrifuged at 3000 rpm for 5 min. The cell pellet was suspended in 100 μL of medium, dispersed evenly, and counted; different concentrations were aliquoted in Hanks' balanced salt solution (HBSS; Gibco–Invitrogen). MPP+- or 6-OHDA-infused rats showing three or more amphetamine-induced rotations/min were transplanted with 2×105, 4×105, or 6×105 cells in 2 μL of HBSS into the right striatum [AP+0.02 from the Bregma point, L −0.26, and DV −0.50] of the animals on the 18th day following the neurotoxin infusion.
Tyrosine hydroxylase immunohistochemistry
Essentially, we followed the immunohistochemical procedure described by us earlier for tyrosine hydroxylase (TH) immunostaining of brain sections passing through the SNpc. 32,34 Animals were anesthetized on the 50th day after toxin infusion, perfused transcardially with 0.1 M phosphate-buffered saline (PBS; pH 7.4) initially, followed by 4% paraformaldehyde perfusion. The brain was dissected out, kept in the fixative overnight, and transferred to 30% sucrose solution. Sections (20 μm) passing through the SNpc and striata were cut on a cryostat and collected on slides. These were rinsed in cold PBS and incubated in 1% hydrogen peroxide (H2O2) for 5 min. This was followed by PBS wash, permeabilization for 30 min in 0.4% Triton X-100, PBS wash (three times, 5 min each), and blocking in 8% bovine serum albumin (BSA) with 0.1% Triton X-100 for 1 hr. Rabbit anti-TH antibody (dilution 1:500; Millipore) was added and kept overnight at 4°C. The sections were washed in PBS with 1% BSA and incubated in anti-rabbit horseradish peroxidase (HRP; 1:1000, Bangalore Genei, India) or anti-rabbit Alexa Fluor 564 (1:1000, Invitrogen) antibody for 1 hr at room temperature. Following the PBS wash, the HRP-tagged sections were transferred in the developing solution for 1–2 min (1 mg of diaminobenzidine [DAB] in 2 mL of PBS was mixed with 100 μL of 1% H2O2). The sections were mounted in DPX Mountant, following dehydration with graded alcohol and clearing in xylene. 32,34 The anti-rabbit IgG antibody-Alexa Flour 564-tagged sections were mounted on slides with Antifade (UltraCruz™, sc-359850, Santa Cruz/ProLong Gold antifade, Invitrogen) mounting medium and viewed in an epifluorescence inverted microscope (Karl Ziess, Germany).
Serial sections (20 μm each) of the whole nigra region were taken, and every tenth section was processed for TH immunohistochemistry (TH-IC). Similarly, sections (20 μm each) passing through the grafts in the striatal region were taken, and every third section was processed for TH-IC. TH-positive neurons in each section (both in the SNpc and in the striatal grafts) were counted employing AxioVision software by three individuals who were blinded to the study. The inter-rater variability in counts was found to be insignificant.
Striatal dopamine levels
Animals were sacrificed on the 50th day, and both sides of the striata (each weighing 10–13 mg) were dissected out independently. The tissues were suspended in cold perchloric acid (HClO4; 0.1 M) containing 0.01% EDTA, sonicated (volume added was about 10 times the weight of the tissue), and kept on ice for 25 min. The samples were then centrifuged at 18,000×g for 10 min at 4°C and an aliquot of 10 μl was injected into a high-performance liquid chromatography (HPLC) system maintained at a 200-nA range of the detection sensitivity and 0.7 mL/min flow rate. The electrochemical detection was performed at 0.74 V. 35
Statistical analysis
The Student t-test was conducted to determine the significance between two for comparing two means in parametric assays. For non-parametric analyses, such as behavioral data, one-way analysis of variance (ANOVA) was performed, followed by a Dunnett test to determine the significance between the groups, employing SigmaStat software package. Correlation analysis was done using Microsoft Excel software. Results are provided as mean±standard error of the mean (SEM). A p value ≤0.05 was considered significant.
Results
6-OHDA–induced rotational bias, neuronal loss, and striatal DA depletion in rats
A loss in the number of TH-positive SNpc neurons was observed on the 50th day after 6-OHDA infusion (Fig. 1A–D). The number of TH-positive neurons in the SNpc of 6-OHDA–infused animals was found to be reduced dose-dependently up to a 16 nmol concentration; thereafter, it reached a plateau (Fig.1E). The reduction in striatal DA content was 48%, 89%, and 99% for 8, 16, and 32 nmol doses, respectively (Fig. 1F). Amphetamine-induced ipsilateral turning bias in animals was enhanced with an increase in 6-OHDA doses, but did not show a clear dose-dependency (Fig. 1G). Apomorphine did not induce turning bias in the 8 nmol 6-OHDA group, whereas for the two higher doses, the bias was towards the contralateral side (Fig. 1H). The percentage of DA depletion in the ipsilateral striatum in animals infused with increasing doses of 6-OHDA was inversely correlated with the number of surviving TH-positive neurons in the SNpc of the same side with an r value of −0.8484 (Fig. 1I). The number of rotations elicited by the animals correlated inversely with the number of surviving TH positive neurons (r=−0.9191; Fig. 1J). Apomorphine-induced rotational bias correlated inversely with the number of surviving neurons (r=−0.8774; Fig. 1K).
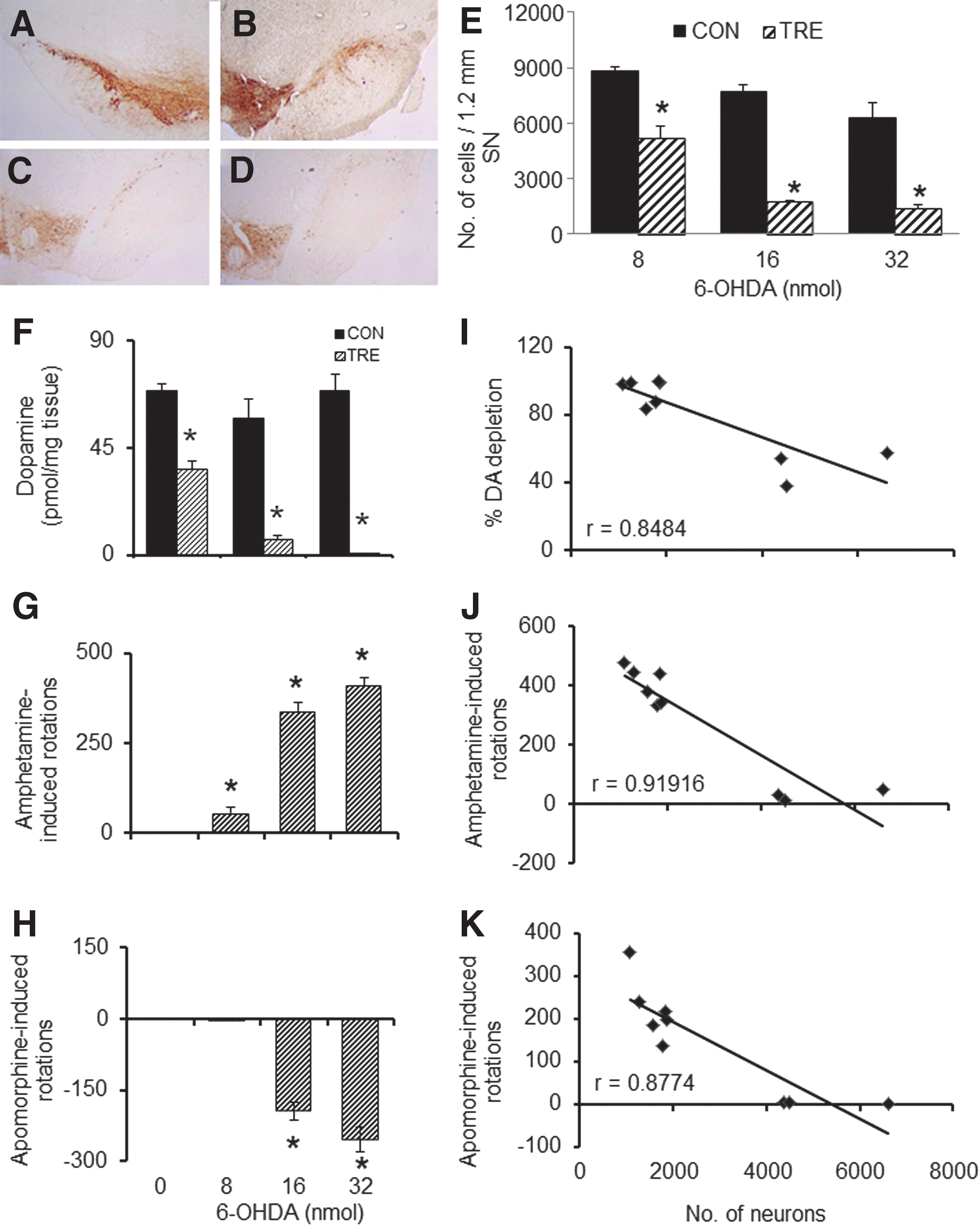
Cellular, neurochemical, and behavioral effects of unilateral intra-median forebrain bundle (MFB) infusion of different doses of 6-hydroxydopamine (6-OHDA). A representative image of tyrosine hydroxylase (TH)-positive cells in substantia nigra (SN) of the un-lesioned side (
Unilateral infusion of MPP+ into MFB in rats leads to progression of PD symptoms and pathology
Intra-MFB administration of MPP+ led to a dose-dependent loss of DA neurons in the ipsilateral SNpc of rats treated with 8 (Fig. 2B), 16 (Fig. 2C), and 32 nmol (Fig. 2D) neurotoxin, compared to contralateral SNpc (Fig. 2A). A stereological analysis of TH-positive DA neurons revealed a dose-dependent loss of 33%, 61%, and 82%, respectively, for 8, 16, and 32 nmol doses of MPP+ (Fig. 2E). A corresponding pattern of loss was seen for striatal DA levels with 59%, 89%, and 96% depletion for 8, 16, and 32 nmol of MPP+-treated rats (Fig. 2F). This was portrayed in the drug-induced rotational bias too. A significant increase of 35% between 8 and 16 nmol doses and an insignificant increase of 12% between 16 and 32 nmol doses of MPP+ was observed for amphetamine-induced ipsilateral turns (Fig. 2G). Apomorphine administration in animals that received the lowest dose of MPP+ (8 nmol) exhibited ipsilateral rotations, whereas the two groups with higher doses of MPP+ displayed contralateral rotations. Between groups with 8 and 16 nmol and 16 and 32 nmol doses of MPP+, there was a significant increase of 140% and 60% rotations, respectively (Fig. 2H). The number of surviving TH-positive cells in the SNpc of rats showed significant inverse correlation between the straital DA depletion (r value=−0.898; Fig. 2I), amphetamine-induced rotations (Fig. 2J; r=−0.679), and apomorphine-induced rotations (Fig. 2K; r=−0.956).
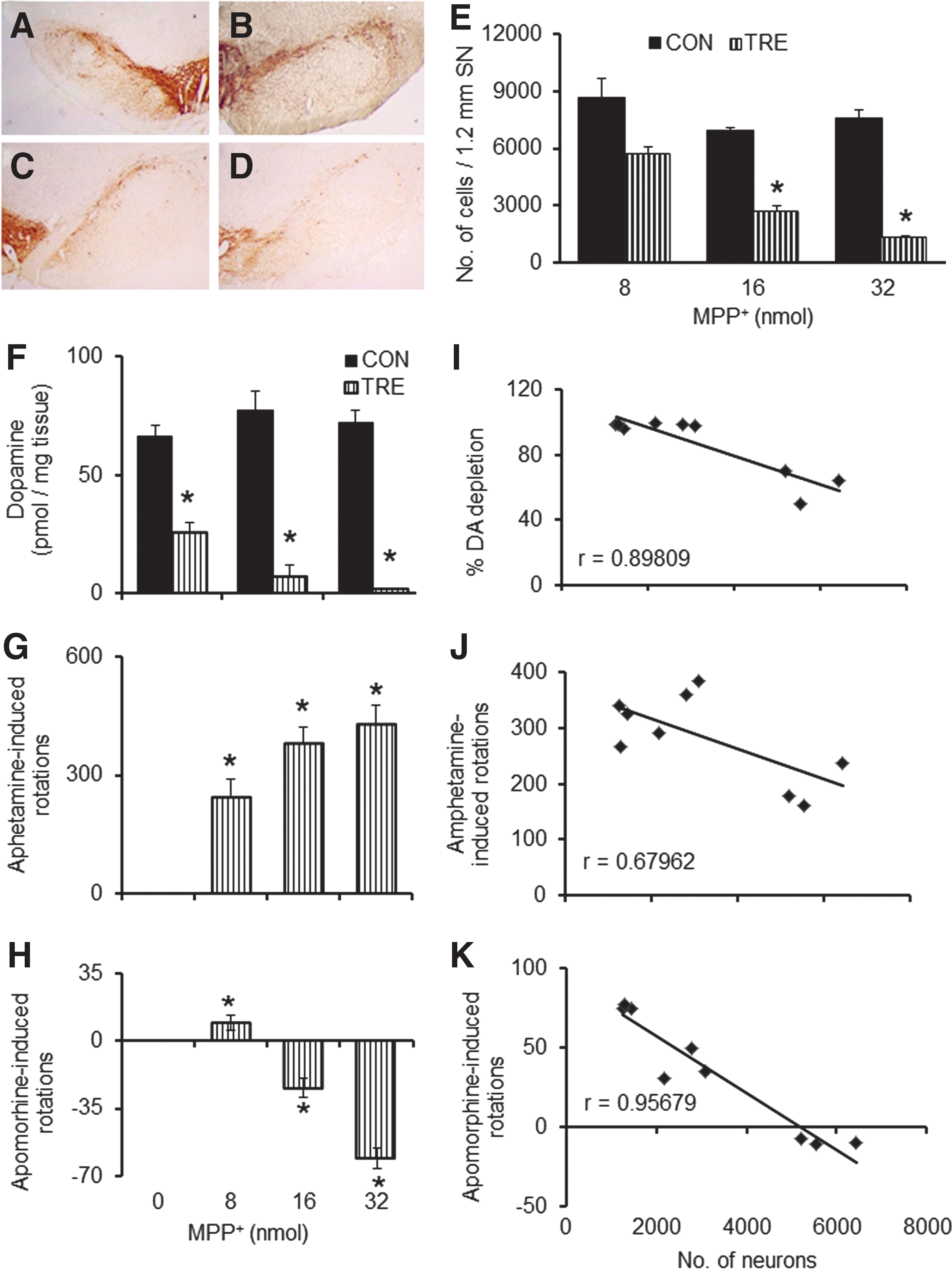
Effects of unilateral intra-median forebrain bundle (MFB) infusion of different doses of 1-methyl-4-phenylpyridinium (MPP+) on substantia nigra pars compacta (SNpc) neurons, striatal dopamine, and behaviors in rats. Immunohistochemistry (
Differentiated mouse ESC transplantation in the striatum of 6-OHDA/MPP+-lesioned animals attenuates rotational bias
We sought to determine the difference in recovery pattern post-transplantation of an equal number of cells in two different doses of neurotoxin-lesioned animals. Amphetamine-induced rotations were monitored on the second week of the neurotoxin infusion and after 2 and 4 weeks of transplantation. When analyzed on the second week following infusion of 16 nmol (Fig. 3A) or 32 nmol (Fig. 3C) of 6-OHDA, these rats exhibited significant ipsilateral rotations. Transplantation surgery was done (2×105 cells per animal) on the 18th day. By 2nd (32nd day) and 4th weeks (46th day) post-transplantation, the number of rotations elicited by these animals declined significantly as compared with the lesioned animals without grafts. When challenged with apomorphine, on the 16th day of neurotoxin infusion the animals in both the groups that received 16 (Fig. 3B) or 32 nmol (Fig. 3D) of 6-OHDA showed significant contralateral turning. Two (34th day) and 4 weeks (48th day) after transplantation, the number of rotations elicited by these animals declined significantly, as compared to non-grafted animals. On the 14th day following 16 nmol (Fig. 3E) or 32 nmol (Fig. 3G) intra-MFB MPP+ infusion, the rats showed significant ipsilateral rotations. Two (32nd day) and 4 weeks (46th day) after transplantation, the number of rotations elicited by these animals was reduced significantly. Apomorphine-induced rotations were monitored on 16th, 34th, and 48th days following MPP+ infusion. On the 16th day of the neurotoxin infusion, the animals that received 16 nmol (Fig. 3F) or 32 nmol (Fig. 3H) of MPP+ showed significant contralateral rotations. Two (34th day) and 4 weeks (48th day) after transplantation, the number of rotations elicited by these animals was significantly reduced as compared to the animals without graft.
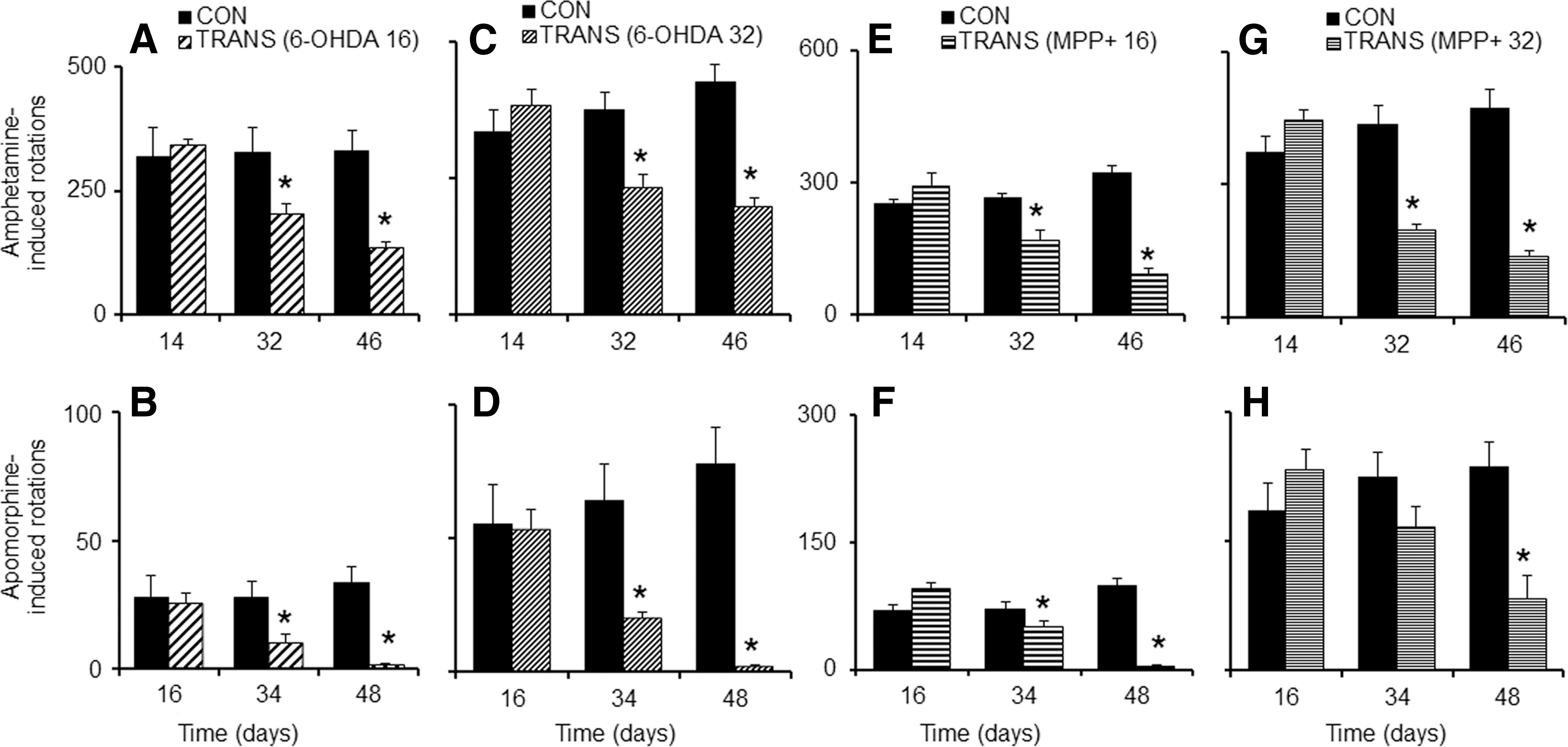
Intra-striatal transplantation alleviates rotational bias in neurotoxin-lesioned animals. Intra-striatal transplantation of 2×105 murine embryonic stem cells (mESCs)-derived neurons was effective in significantly reducing amphetamine-induced (
Survival of TH-positive cells in the graft and recovery in striatal DA levels
Transplantation of 2×105 cells in animals lesioned with different concentrations of the toxin, regardless of the type (6-OHDA or MPP+), showed variation in the graft size as seen by the TH immuno-labeling of the implanted cells. A dose of 32 nmol of 6-OHDA (Fig. 4E) or MPP+(Fig. 4K) showed reduced TH immunoreactivity compared to the 16 nmol-infused groups of either toxin (Fig. 4B, H), irrespective of the fact that an equal number of cells were transplanted in all the groups. The first column (Fig. 4A, D, G, J) shows the nuclei (stained with 4′,6-diamidino-2-phenylindole [DAPI]) for the same field, and the last column (Fig. 4C, F, I, L) shows merged images of both the fields. Animals were sacrificed and analyzed for their striatal DA levels on the 50th day of the neurotoxin infusion, i.e., 32 days post-transplantation. There was significant improvement in the percentage recovery (about 34%) of striatal DA in low-dose 6-OHDA groups (16 nmol) with transplantation (Fig. 5A), but in animals that received higher dose of 6-OHDA (32 nmol), the percentage of DA recovery was only about 15% (Fig. 5A). In MPP+-treated animals, the 16 nmol group showed 42% DA recovery following transplantation; however, no recovery was seen in the animals that received the higher dose of MPP+ (Fig. 5D).
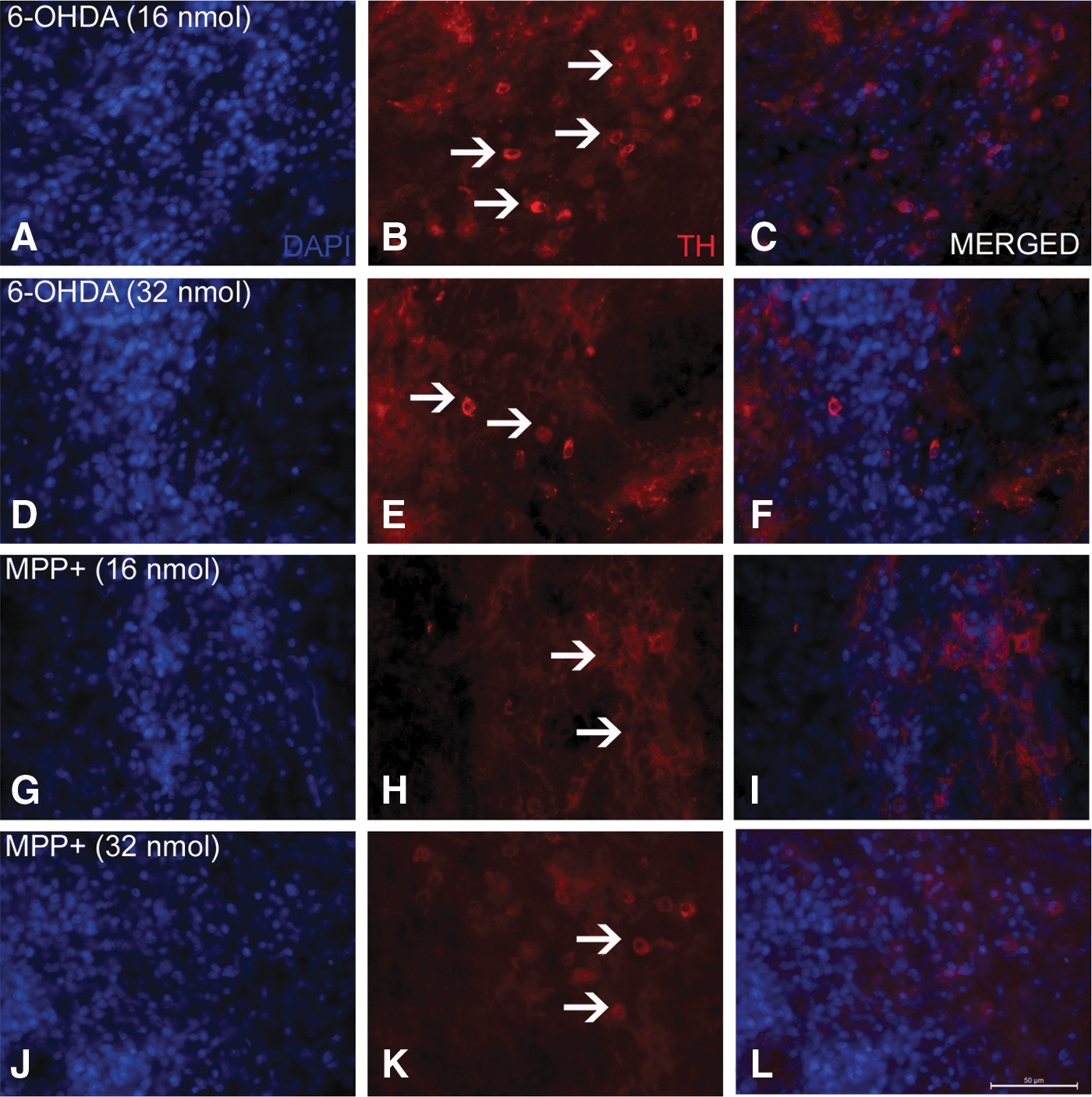
Representative pictures of tyrosine hydroxylase (TH) immunoreactivity in the striatal graft showing dopaminergic neurons in the graft. Images show striatal brain sections immunostained for TH taken from animals lesioned with 16 and 32 nmol 6-hydroxydopamine (6-OHDA) (
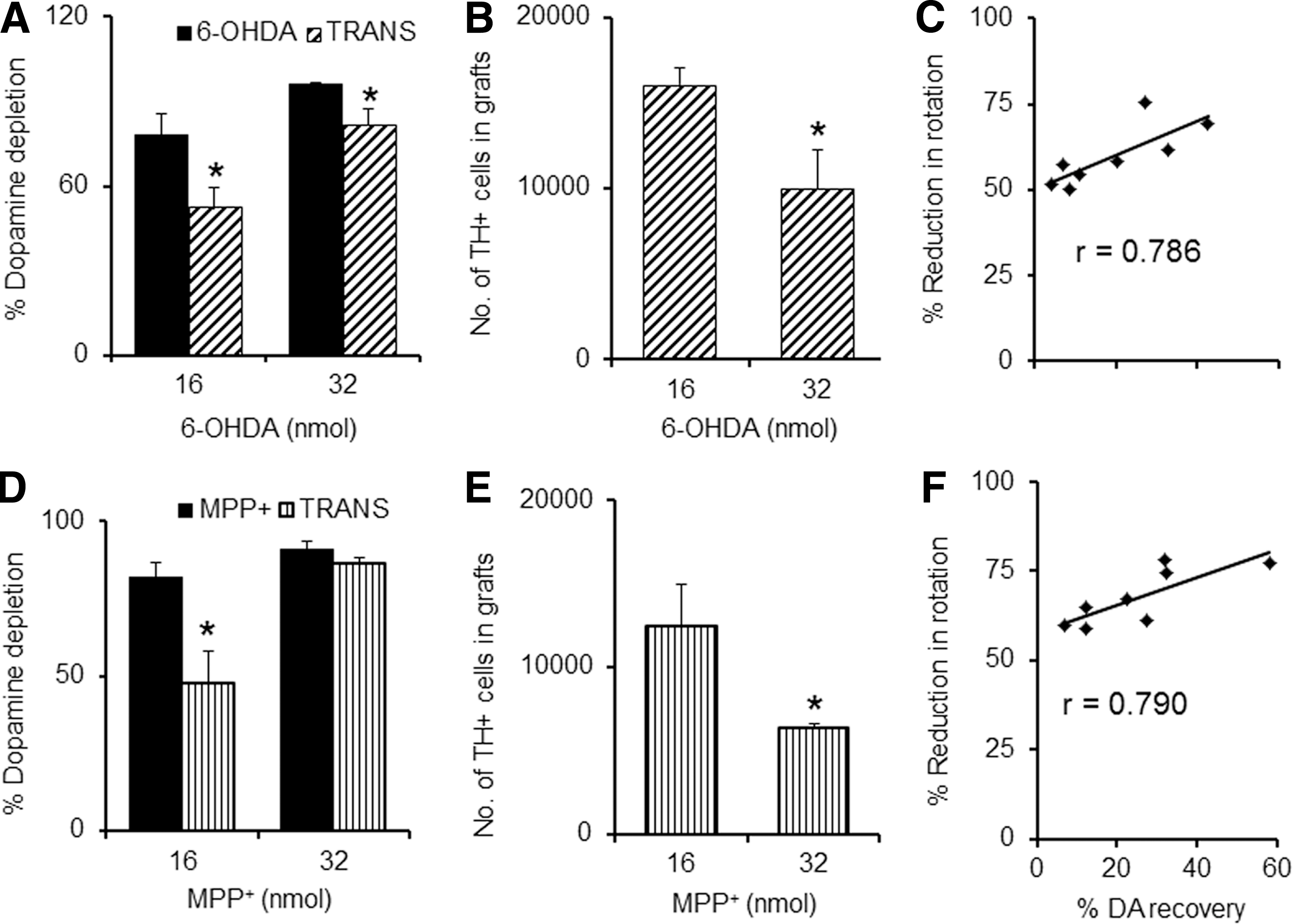
Post-transplantation striatal dopamine (DA) level recovery and the surviving tyrosine hydroxylase (TH)-positive cells in the graft. Reduction in DA depletion in the striatum, being more in the 16 nmol than in 32 nmol of 6-hydroxydopamine (6-OHDA)-infused animals (
The number of TH-positive neurons in the graft was higher in animals that received the lower dose of the toxin, but about 38% lower in the group that received the higher dose of 6-OHDA (Fig. 5B). MPP+-treated animals showed lesser surviving cells in the graft as compared to 6-OHDA. The lower dose of MPP+ (16 nmol) showed about 49% more surviving cells in the striatal grafts than the graft in animals that received the higher dose of MPP+ (Fig. 5E). The correlation between the percentage of reduction in amphetamine-induced rotations and the percentage of DA recovery on the 50th day of the treatment program was analyzed (Fig. 5C, F). A good correlation was observed, with a coefficient r of 0.786 and 0.790, respectively, for 6-OHDA (Fig. 5C) and MPP+ (Fig. 5F). No significant correlation was observed for the percentage of reduction in apomorphine-induced rotations with the percentage of DA recovery (data not provided).
Recovery in amphetamine-induced rotations is directly related to number of cells transplanted
Our next goal was to see if by varying the number of cells transplanted in a group of animals lesioned with the same dose of neurotoxin, the degree of recovery is affected. mESCs differentiated for 7 days in culture were intra-striatally transplanted in 6-OHDA- (32 nmol; Fig. 6, top panel) and MPP+- (32 nmol; Fig. 6, lower panel) lesioned animals on the 18th day at various cell concentrations (2×105, 4×105, 6×105 cells in 2 μL). Ipsilateral rotations after amphetamine injection on the 14th, 32nd, and 46th days of toxin infusion were monitored. On the 46th day, a significant reduction in the amphetamine-induced rotations was observed in the three groups of animals that received different counts of cells, which were proportional to the number of cells transplanted [for 2, 4, and 6×105 cells 64%, 76%, and 79% (6-OHDA, Fig. 6A, C, E) and 48%, 60%, and 69% (MPP+, Fig. 6B, D, F) reductions, respectively]. When correlation analysis was performed for the percentage of reduction in amphetamine-induced rotations and the number of cells transplanted, a moderate correlation between the two groups was observed in 6-OHDA- (r=0.58) (Fig. 6G) or MPP+-lesioned rats (r=0.56) (Fig. 6H).
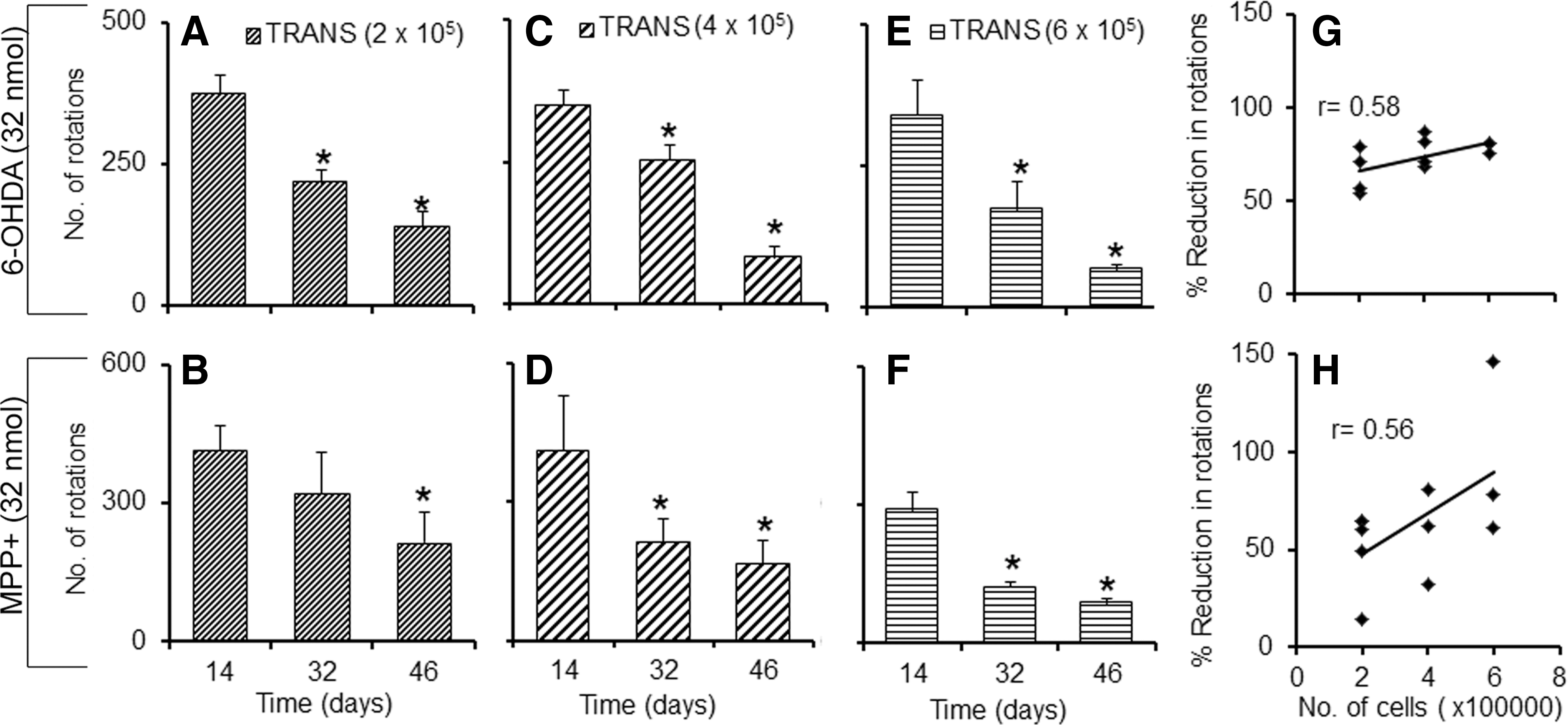
Effect of increasing graft cell number on amphetamine-induced rotations in hemi-parkinsonian rats. Unilateral intra-striatal transplantation of 2×105 cells (
Discussion
We have used two hemi-parkinsonian models of rats created following unilateral intra-MFB infusion of 6-OHDA or MPP+, which are known to denervate the nigrostriatal dopaminergic pathway and induce turning behavior following administration of dopaminergic drugs such as apomorphine and amphetamine. 24,27 The major findings are: (1) Amphetamine- or apomorphine-induced rotations correlated well with the severity of the disease in both 6-OHDA- and MPP+-induced animal models of PD in terms of striatal DA depletion and TH-positive neuronal loss in the SNpc. (2) Reduction in amphetamine-induced rotations after transplantation correlated well with the DA recovery following differentiated mESCs transplantation. (3) The reduction in amphetamine-induced rotation correlated significantly with the number of cells transplanted in the striatum. (4) Survival of the graft DA-ergic neurons was found to be better in animals with minor SNpc neuronal lesions than in animals with severe lesions.
These results are very similar and comparable in two animal models of hemi-parkinsonism, as assessed in the present study. Critical evaluation of the results, however, revealed that 16 and 32 nmol/animal doses of 6-OHDA have comparable neurotoxic potency in striatal DA depletion and loss of SNpc neurons, whereas MPP+ treatment at these doses caused more, but controlled, dopaminergic neurotoxicity. These subtle differences are reflected in the biased rotational behavioral outcome following amphetamine or apomorphine treatment in this model.
At a time when 8, 16, and 32 nmol/animal intra-MFB MPP+ infusion could be used for creating rats with low, moderate, and severe stages of PD, respectively, the study with the other neurotoxin reveals that a dose of 32 nmol of 6-OHDA is too high and doses less than 16 nmol should be used for a moderate stage of the disease in rats. Animals lesioned with 6-OHDA were more sensitive to apomorphine-induced rotations, with almost five-fold more counts as compared to MPP+-lesioned animals. This could be due to increased supersensitivity of the post-synaptic dopamine receptors in the 6-OHDA-lesioned animals owing to severe striatal DA depletion in these animals as compared with MPP+-lesioned animals. 36 The information generated from the present study in this regard is the first time it has been reported in the literature, because no other group has tried to compare these three dependent variables—dopaminergic neuronal loss, striatal DA loss, and their relationship to unilateral rotations to predict three independent stages of PD (low, moderate, and severe stages) in animal models. This information may not have direct implications to Unified Parkinson's Disease Rating Scale or Hoehn and Yahr scores in human PD, but could be a classical way to define the three useful, defined stages in unilateral parkinsonism in rats.
We have used mESCs differentiated to neuronal lineage for 7 days in culture by maintaining them in a serum-free media; these cells have been found to be more efficient for transplantation when compared to undifferentiated mESCs or cells that have been differentiated for 10 days. 32,37 When equal numbers (2×105 cells) of these cells differentiated for 7 days were transplanted in the striatum of animals with two different doses (16 and 32 nmol) of 6-OHDA- or MPP+-infused animals, we observed the extent of recovery in terms of amphetamine- and apomorphine-induced rotations. Striatal DA levels were better in animals that received a lower dose of the toxin, irrespective of the type of toxin used. Our study also supports a previous report by Robichon et al. 38 in which they have demonstrated graft survival is independent of the neurotoxin used and survival of the grafted cells was better in animals that received a lower dose (6%–8%) and not a higher dose (3%–4%) of the neurotoxin. In analogous situations, clinical trials conducted in younger patients showed better transplantation recovery than in trials with older patients. 39 In our hands, we have found that mESCs differentiated with a mixed population of cells provided better transplantation recovery than pure DA-ergic neurons sorted from the mixed population. 37
A study regarding details of certain transplantation procedures showed that fragmented cell suspensions as compared to single-cell suspensions, glass capillary as compared to metal cannula, and lesioned striatum as compared to intact striatum provide improved chances of cell survival. 40 In agreement with this study, Kirik et al. 41 illustrated that the survival of the grafts was better in the animals showing greater striatal denervation (>70%) as compared to those with partial denervation.
In our study, we have considered two doses of toxin where reduction in striatal DA levels (directly proportional to striatal denervation) is greater than 70%, suggesting that both the transplanted groups considered for our study have severe striatal DA neuron loss. So cell survival in our study is not affected by the host striatal terminals, unlike the study by Kirik and co-workers. 41 Rather we speculate it is influenced by the local hostile environment of the higher toxin dose group. It may be that the striatum retains some of the neurodegenerative signals that adversely affect the survival of neurons and, hitherto unknown, similar to inheritance of protein aggregation pathology within the transplanted fetal neurons in human PD striatum. 42,43 A higher dose of the toxins might have elicited inflammatory responses beyond physiological control caused by microglial activation, 44 which in turn can lead to cell death, and was also demonstrated in our previous reports. 32,37 Because hypoxia, inflammatory trauma, and excitotoxic events could be the striatal environment of the lesioned side, 32,34,39,45 we suppose that the higher the dose of neurotoxin, the more hostile will be the condition for survival of the transplanted cells. Some studies suggest that ectopic placement of the graft, i.e., the striatum instead of SN, may be an issue that affects the survival of the grafts, the reason being the lack of trophic factors required for survival and maintenance of dopaminergic neurons. 46,47 However, we have demonstrated recently 32 that there is more than a four-fold increase in the glial cell–derived neurotrophic factor in the grafted striatum, ipsilateral to the side of injury, following 16 nmol of MPP+, which is commensurate with astrocytosis surrounding the graft. 32,37
Politis and Lindvall 2 have mentioned that the survival of >100,000 cells in human striatum would ensure transplantation recovery. In our study, about 12–16×103 and 6–9×103 cells were surviving in the ipsilateral striatum of severely SNpc-lesioned animals after implantation of 2×105 cells. Transplantation of higher doses of cells (4 and 6×105) in both 6-OHDA- and MPP+-induced, severely SNpc-lesioned animals showed significant reductions in amphetamine-induced rotations, as compared to the scores prior to transplantation. The results show that reduction in the percentage of amphetamine-induced rotation correlates with the increase in the number of cells transplanted (r=0.56 and r=0.58 for MPP+ and 6-OHDA, respectively). This could be due to greater number of grafted cells surviving in the striatum, which has not been verified in the present study. Carman et al. 48 have reported that no correlation existed between surviving TH-positive cells and amphetamine-induced rotations in the 6-OHDA lesioned hemi-parkinsonian rat model, whereas apomorphine-induced rotations showed significant correlation. However, in our study, we show a correlation between the surviving TH-positive cells and amphetamine-induced rotational bias. The pathological hallmark of PD is a gradual loss of nigostriatal DA neurons, but neuronal degeneration also occurs in non-DA-ergic systems. 49 The more severe the disease, the more chance there is of other cell types being affected in the brain. A scale of severity of the disease as obtained from the present study will provide basis of neurons affected in SNpc, and hence would help to estimate the number and type of the cell population to be transplanted.
In conclusion, we would like to state that both MPP+ and 6-OHDA with carefully applied doses, can bring about controlled nigral neuronal damage. In addition, the extent of neuronal lesion could be associated with stages of the disease—low doses/less nigral cell loss, medium doses/moderate nigral cell loss, and high doses/extensive nigral cell loss, respectively, representing early, mid-stage, and advanced stage of the disease (or low, moderate, and severe stages of experimental PD), being evidenced by extent of behavioral dysfunction. The disease stages thus determined could help to predict the number of cells required to be transplanted in hemi-parkinsonian rats for a significant recovery in terms of amphetamine-induced rotary bias. In short, the more the loss of TH-positive neurons in SNpc, the higher is the number of cells required to be transplanted to bring recovery. The moot question is whether such a paradigm can work in human condition or not.
Footnotes
Acknowledgments
D.T. and R.H. received Council of Scientific and Industrial Research (CSIR) and University Grants Commission (UGC) fellowships, respectively. The study was funded by the project Neurodegenerative Disorders: Causes and Corrections (miND; BSC 0115) under CSIR's 12th 5-year plan program.
Author Disclosure Statement
No competing financial interests exist.