
Abstract
Parkinson's disease (PD) is the second most common neurodegenerative disease, and it is characterized by the loss of the neurotransmitter dopamine and neuronal degeneration in the substantia nigra pars compacta. Thus far, current therapeutic strategies have failed to address neuronal degeneration. It has been reported that overproduction of reactive oxygen species, resulting in oxidative stress, and neuroinflammation play an important role in neurodegenerative diseases through the induction of macromolecular oxidative damage and modulation of intracellular signaling pathways concurring to neuronal cell death. Indeed, anti-oxidant and anti-inflammatory drugs have been the subject of recommendation as a complementary therapy alongside an effective symptomatic treatment to hamper the progression of PD. Today, much attention is paid to polyphenols in light of their potent capacity to reduce oxidative stress and inflammation, while having much fewer side effects than most other drugs. Camellia sinensis L. is the most common ancient herbal tea prepared as a beverage worldwide and it possesses numerous beneficial effects on human health. Epigallocatechin-3-gallate is the best-known bioactive component of C. sinensis and is recognized to exert potent neuroprotective effects against oxidative stress, neuroinflammation, protein aggregation, autophagy, and neuronal cell death in vitro as well as in vivo. The present review appraises the available literature on the beneficial role of epigallocatechin-3-gallate pertaining to dopaminergic degeneration characteristic of PD with particular emphasis on its possible mechanisms of action.
Introduction
P
Edible medicinal herbs are regarded as promising candidates for the development of complementary therapeutic strategies in PD and other central nervous system (CNS) diseases in light of centuries of use in traditional medicine. 10,11 Camellia sinensis L. is one of the most important medicinal plants and is also known as the most commonly consumed tea worldwide. 12 Considering its vast, and ever-increasing, usage, its extensively proven safety, as well as the bioavailability of its bioactive compounds, numerous investigations have been performed on the pharmacological effects of C. sinensis and/or its bioactive molecules. 13 Epigallocatechin-3-gallate (EGCG) is recognized as the most abundant 14 and most anti-oxidative 15 molecule of C. sinensis, accounting for 50%–80% of total catechin content, which represents 200–300 mg in a cup of brewed green tea. EGCG's anti-oxidative, anti-inflammatory, and fibril-destabilizing potential may be responsible for the neuroprotective effects of green tea. 16,17
The three main avenues for drug development strategies in PD currently focus on: (1) Improving in DAergic therapies and prevention of the motor complications, (2) identifying non-DAergic drugs for symptomatic improvement, and (3) discovering disease-modifying or neuroprotective compounds. 18,19 Elucidation of the pharmacologically valuable role of EGCG in PD falls under the second and third strategic ambitions. The present paper aims to review the most salient literature on the beneficial effects of EGCG in in vitro and in vivo pre-clinical PD studies. Focus will be drawn to the potential mechanisms of action of EGCG.
Pathophysiology of Parkinson's Disease
Oxidative stress is currently known to play a prominent role in dopamine cell degeneration in the SNc. 20 DAergic neurons in this region of the CNS are particularly vulnerable to oxidative stress due to the high activity of monoamine oxidase, 21 the enzyme responsible for metabolizing dopamine (DA), and elevated levels of iron, 22 which both lead to increased generation of reactive oxygen species (ROS). High turnover rates of DA in the SNc ensured by monoamine oxidase contribute to the autotoxicity of catecholamines and their metabolites in the cells in which they are produced, as reported in PD. 23 Iron ions react with hydrogen peroxide, readily found in the SNc, to form the very deleterious hydroxyl radical through the Fenton reaction. The resulting oxidative stress damages proteins, lipids, and DNA and concurs with the production of oxidant by-products (e.g., 4-hydroxynonenal derived from lipid peroxidation) that, by reacting with proteins, may induce their aggregation and thereby impairing cellular viability. 24,25
Neuroinflammation is also recognized as a key factor in the initiation and progression of PD pathology 26,27 and mainly manifests itself as the excessive activation of microglia, the resident macrophages of the CNS. Indeed, postmortem studies have revealed the existence of high levels of major histocompatibility complex class II cell-surface receptors in activated microglia in brains of PD patients. 28 Interestingly, under neuroinflammatory conditions, glial cells readily express inducible nitric oxide synthase (iNOS) responsible for the production of nitric oxide (NO). Superoxide anion, a monoamine oxidase B by-product, reacts with NO to generate peroxynitrite, which is ultimately decomposed into the very reactive hydroxyl radical. In that sense, oxidative stress and neuroinflammatory actions intricately entwine in the pathophysiology of PD.
PD is the second most frequent protein misfolding disease and the most common synucleinopathy. Indeed, one of the principal hallmarks of this disease is the presence of intracellular Lewy body or Lewy neurite inclusions in DAergic neurons of the SNc, although some forms of PD exist without this fibrillization characteristic. Lewy bodies and neurites, respectively, found in the soma and processes of nerve cells, are composed of a complex mixture of proteins, primarily the presynaptic protein alpha-synuclein, and to a lesser extent ubiquitin, tau, and amyloid-beta, among several others. Research on the implication of alpha-synuclein in PD pathology exploded when a single point mutation (Ala53Thr) of SNCA, the gene encoding alpha-synuclein, was uncovered in a dominantly inherited form of early onset familial PD in a large Italian pedigree and in three unrelated Greek families. 29 As is the case for many other amyloidogenic proteins, experts have not identified the exact alpha-synuclein fibrillar structures responsible for cellular toxicity, although there is growing consensus that the process of fibrillization rather than the end product may be cytotoxic. 30 –33
Intracellular Lewy body inclusions also point toward impaired protein clearance and may play a role in mitochondrial dysfunction. The autophagy–lysosomal degradation pathway is indispensable for turnover of mitochondria and degradation of aggregated proteins. It has recently been linked with the development of neurodegenerative disorders such as PD. 34 –36 Mutations in risk genes associated with PD, such as parkin, DJ-1, phosphatase and tensin homolog (PTEN)-induced putative kinase 1 (PINK1), and leucine-rich repeat kinase 2 (LRRK2), provoke the accumulation of dysfunctional mitochondria and/or protein aggregates, leading to an augmented oxidative burden. 37 Moreover, these genes are also important in mitochondrial homeostasis and energy management thought to be impaired in PD. Hence, impaired autophagy-lysosomal degradation pathways and dysfunctional mitochondria may prompt increased oxidative stress and, over time, cell death and apoptosis.
Epigallocatechin-3-Gallate
Chemistry
EGCG is a polyphenol of the flavonoid family belonging to the chemical class of flavan-3-ols whose structure consists of four rings resulting from the esterification of epigallocatechin (EGC) with gallic acid (Fig. 1). 38 The A and C rings constitute the benzopyran ring which has a pyrogallol moiety at position 2, the B ring, and also a gallate moiety at position 3, the D ring. In particular, the galloyl ester is responsible for the exposure of EGCG to nucleophilic attacks. 39
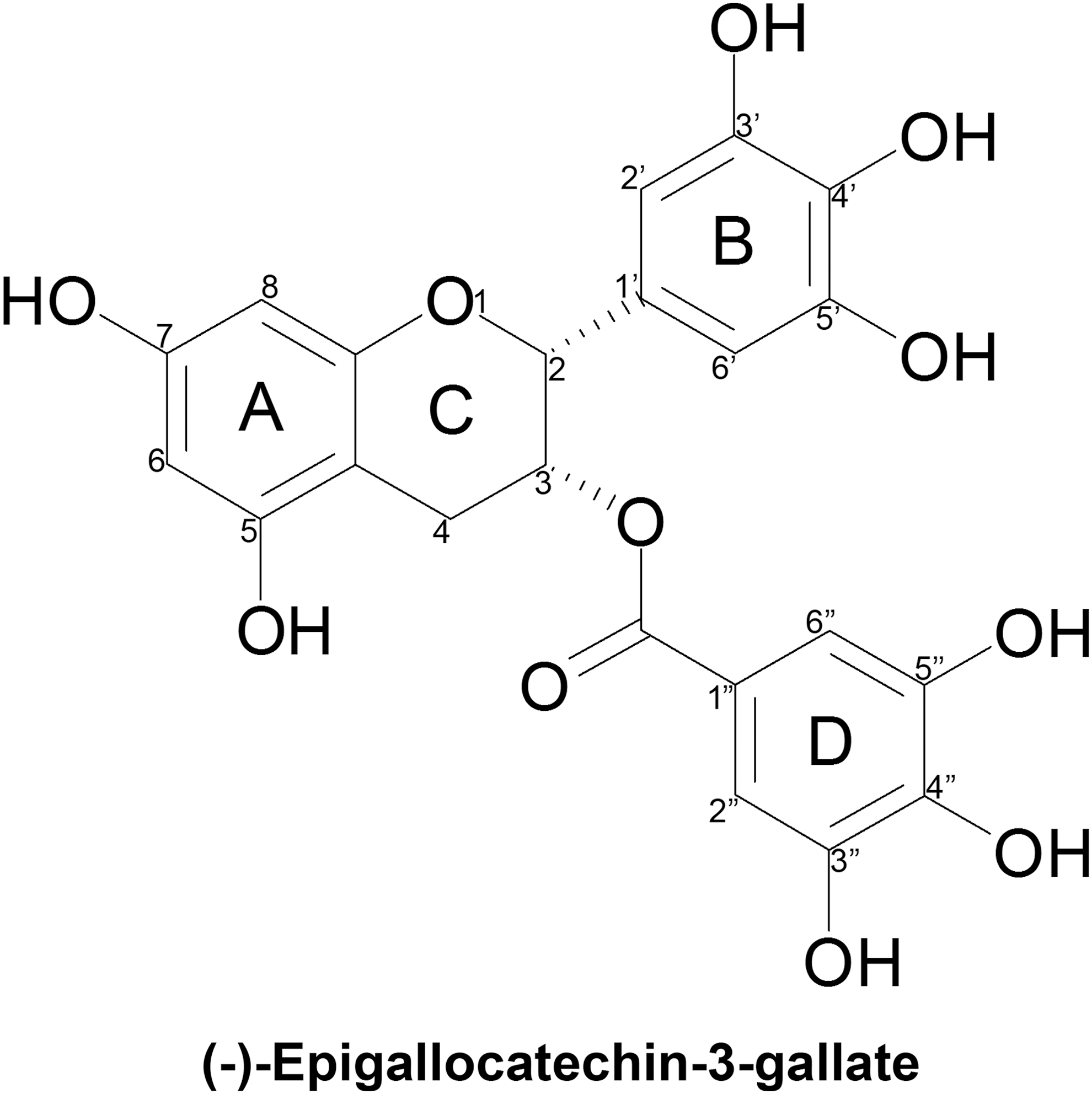
Chemical structure of epigallocatechin-3-gallate.
EGCG owes its anti-oxidative potential chiefly to the multiple hydroxyl functions that are found on the B and D rings at positions 3′, 4′, 5′, 3″, 4″, and 5″ positions, as well as at positions 5 and 7 of the benzyopyran ring. 40 The several ortho-dihydroxy pairings found in rings B and D account for EGCG's potent divalent metal chelating capacity. Guo and colleagues found the chelated ratios of EGCG with Fe (III) to be 2:1. 41 EGCG's numerous hydroxyl groups as well as its conjugated structure, ideal for electron delocalization, confer on this molecule important free radical scavenging properties. 42 It was found that decreasing the number of hydroxyl groups in the B ring dims the potency of its biological activities. 43 Moreover, compared to its structural analogs (−)- EGC, (−)-epicatechin (EC), and (−)-epicatechin-3-gallate (ECG) that all possess fewer hydroxyl groups, EGCG was found to be a more efficient radical scavenger. 44
Anti-oxidant and pro-oxidant mechanisms
Currently, there are numerous scientific reports about the anti-oxidative capacity of EGCG under both in vitro and in vivo conditions. 45,46 Interestingly, it has been reported that EGCG improves mitochondrial function through its anti-oxidative action. 47 In addition, EGCG mitigates lipid infusion–induced insulin resistance by activating 5′-adenosine monophosphate-activated protein kinase (AMPK) pathways and increasing the expression of superoxide dismutase (SOD) and glutathione peroxidase (GPx), endogenous anti-oxidant enzymes. 48 Moreover, it inhibits the expression of cyclo-oxygenase-2 (COX-2) and iNOS, enzymes responsible for generating pro-inflammatory mediators leading to oxidative stress. 49 Despite these properties, there are also several studies reporting the putative pro-oxidative actions of EGCG. 50 Indeed, EGCG produces hydrogen peroxide by a process of auto-oxidation. 51 The addition of anti-oxidant enzymes, such as SOD and catalase, inhibits some pro-oxidative actions of EGCG in HL60 and RAW 264.7 cells through inhibition of its auto-oxidation and dimerization. 52
Up to now, two mechanisms have been suggested for the pro-oxidative activity of EGCG, which is in fact thought to lie beneath its anti-cancer properties. The first mechanism is related to the production of hydrogen peroxide via the pyrogallol moiety in the chemical skeleton of EGCG. 53 The second mechanism is the production of ROS, such as hydroxyl radicals, by the reduction of Fe(III) to Fe(II) through the Fenton reaction. 53 Thus, when used at high concentrations, the combined formation of both hydroxyl radical and hydrogen peroxide could be held accountable for EGCG's cytotoxic potential. 53 However, at low concentrations, EGCG generates low levels of ROS, which may, by means of hormesis, stimulate cellular signal transduction pathways and subsequently trigger protective mechanisms in cells. 15 Furthermore, it has been reported that EGCG affects the cellular anti-oxidant defense system by modulating important players such as glutathione (GSH), thioredoxin, malondialdehyde (a marker of lipid peroxidation), catalase, and SOD. 54,55 Noteworthy, it must be emphasized that the direct and indirect effects of EGCG on the cellular anti-oxidant system and its metabolites may also have both anti-oxidative and pro-oxidative actions. 42,56
Pharmacokinetics and bioavailability
Numerous studies report the pharmacokinetic profile of oral administration of EGCG. 57 –59 Among all green tea catechins, EGCG possesses the longest half-life in rats, 60 and it is the only known polyphenol abundantly available in human plasma (77%–90%) in a free form, 57,61 –63 whereas other catechins are highly glucuronidated or sulfated. Nevertheless, it remains that EGCG is generally poorly absorbed when administered orally due to its high solubility, making for low membrane permeability. After gastric digestion, EGCG is stable, but under duodenal conditions this molecule reacts with digestive enzymes, and a high-molecular-weight galloyl derivative is produced. 64 Van Amelsvoort and colleagues further demonstrated that galloylation of catechins reduces their absorption. 65 Moreover, EGCG is a substrate of methylation, and 4′,4″-di-O-methyl-EGCG accounts for 15% of total EGCG circulating metabolites in humans. 62
Ullmann and colleagues evaluated the pharmacokinetic profile of EGCG after a single-dose administration ranging from 50 to 1600 mg and reported that at doses higher than 1000 mg, its maximal plasma levels (Cmax) were about 1μM (for example, after administration of EGCG at 1600 mg, Cmax was 3392 ng/mL with a range of 130–3392 ng/mL). 63 Plasma Cmax was observed between 1.3 to 2.2 hr after oral administration. Another study on the plasma kinetics of EGCG and its conjugated metabolites, performed at intervals for a 26-hr period after oral administration, indicated that the mean total EGCG area under the plasma concentration time curve between 0 hr to infinity (AUC (0–∞)) ranged from 442 to 10,368 ng * h/mL and mean terminal elimination half-life (t 1/2z) was from 1.9 to 4.6 hr. 63 In addition, another study examined the plasma kinetics of different doses of purified EGCG after administrations of 800 mg once per day and 400 mg twice per day for 4 weeks. 57 A peak in the serum levels of EGCG was observed after its administration at 400- and 800-mg concentrations. 57 It has also been reported that the bioavailability of EGCG increases after chronic administration of 800 mg. 66
The ability of EGCG to cross the blood–brain barrier (BBB) is crucial for the development of therapeutic strategies in PD. A few studies have shown that EGCG indeed crosses the BBB. 67–68 In particular, male and female mice orally administered with 200 μL of a 0.05% EGCG solution containing 3.7 MBq [ 3 H]EGCG displayed 0.32% and 0.33%, respectively, of total radioactivity in the brain 24 hr after uptake, which was comparable with most other organs. 68
EGCG and PD
Rationale for the use of EGCG in PD
Compelling epidemiological evidence supports a correlation between green tea intake and a reduced risk of developing PD. Indeed, an American epidemiological investigation showed that people that consumed two cups per day or more of tea presented a decreased risk of PD. 69 Similarly, a prospective study of ∼30,000 Finnish adults followed for a 13-year span found that drinking three cups or more of tea daily was associated with a reduced occurrence of PD. 70 However, no distinction between green and black tea, the latter containing up to four times less EGCG because it is fermented, was made in the questionnaire, making it impractical to link these findings with EGCG. The potential relevance of green or black tea consumption was investigated in the prospective cohort Singapore Chinese Health Study. Surprisingly, results showed that black tea drinking was inversely associated with PD risk, a link that was not made with green tea consumption. 71 A retrospective study reported that consumption of more than three cups of tea per day hindered age of motor symptoms onset by an average of 7.7 years in 278 Israeli PD patients. 72 Again in this study, no distinction was made between black and green tea.
On the basis of these epidemiological data, a few clinical trials were launched to better define the efficacy and safety of EGCG or green tea polyphenols in PD. A first double-blind, randomized, placebo-controlled, delayed study sought to determine the safety and efficacy of green tea polyphenols in slowing disease progression in patients with de novo PD (
These clinical and epidemiological findings offer scarce and inconclusive data on the efficacy of green tea polyphenols, EGCG, or green tea consumption in PD. For this reason, there is an imperative need for better-designed studies in humans. Investigations performed in pre-clinical models are warranted to further unravel the mechanisms of action of EGCG to help orient future clinical trials in view of obtaining convincing results. At any rate, owing to its broadly verified safety and almost inexistent side effects in humans, except for minor gastrointestinal abnormalities, 38,74 EGCG offers an advantage over current drugs for PD, which are almost always tied in with significant adverse symptoms, chiefly dyskinesia. In view of future clinical applications, it is noteworthy to underline that its isolation is inexpensive, and therefore large amounts of EGCG can be recovered from green tea. The following sections review pre-clinical literature on EGCG neuroprotection, addressing core issues in PD, such as oxidative stress, neuroinflammation, protein aggregation, and, more recently, mitochondria homeostasis. The most salient results are summarized in Table 1.
6-OHDA, 6-hydroxydopamine; MPP+, 1-methyl-4-phenylpyridinium; ROS, reactive oxygen species; SIRT, sirtuin; PGC-α, peroxisome proliferator-activated receptor gamma co-activator-α; SOD, superoxide dismutase; GPx, glutathione peroxidase; GSH, glutathione; GSK-3, glycogen synthase kinase-3; PKC, protein kinase C; ERK1/2, extracellular-signal-regulated kinase 1/2; TNF-α, tumor necrosis factor-α; MPTP, 1-methyl-4-phenyl-1,2,3,6- tetrahydropyridine; p.o., per os; DA, dopamine; TH, tyrosine hydroxylase; PKC, protein kinase C; FasL, Fas ligand; Gadd45, growth arrest and DNA damage protein; LPS, lipopolysaccharide; NO, nitric oxide; iNOS, inducible nitric oxide synthase; CM, conditioned medium; nNOS, neuronal nitric oxide synthase; EGCG, epigallocatechin-3-gallate.
Oxidative stress
Several models of PD exist in vivo as well as in vitro that depend on pro-oxidative mechanisms to induce parkinsonian-like features of DAergic neuronal death. Some neurotoxic compounds such as 6-hydroxydopamine (6-OHDA), rotenone, paraquat, and 1-methyl-4-phenylpyridinium (MPP+, the active metabolite of 1-methyl-4-phenyl-1,2,3,6- tetrahydropyridine [MPTP]) interfere with the electron transport chain, generating acute ROS over-production, which leads to oxidative stress. 75 –78 Hydrogen peroxide, a reactive oxygen species, as well as the DA precursor levodopa have also been used to generate oxidative stress in cell cultures. It is important to mention that, among all CNS cell types, DAergic neurons are particularly vulnerable to these toxins at different levels. EGCG has been shown to promote the survival of PC12 cells, an in vitro catecholaminergic model, against treatment with 6-OHDA, 79,80 MPP+, 81 paraquat, 82 levodopa, 83 and hydrogen peroxide. 84,85
When comparing several green tea polyphenols, both EGCG and its analog ECG were shown to confer the greatest polyphenols against 6-OHDA–induced apoptosis. 79,80 Interestingly, although MPP+or EGCG alone did not cause any significant changes in silent mating type information regulation 2 homolog 1 (sirtuin 1, SIRT1) and peroxisome proliferator-activated receptor gamma co-activator 1-alpha (PGC-1α) levels, a combined treatment on differentiated PC12 was found to increase their expression. 81 SIRT1 deacetylates PGC-1α, which activates its transcriptional activity.
Downstream transcriptional targets of PGC-1α, such as the anti-oxidant enzymes Cu/Zn SOD and GPx-1, were found to be up-regulated by EGCG pre-treatment, and ROS levels were concomitantly diminished. 81 EGCG was also shown to improve levels of GSH while reducing lipid peroxidation and ROS levels. 83 Moreover, EGCG protects SH-SY5Y cells, a neuronal DAergic in vitro model, from oxidative stress resulting from 6-OHDA administration, 86 although Chung and colleagues have shown, on the other hand, that pre-treatment with EGCG potentiates rotenone toxicity in these same cells. 87
This could be explained by differences in experimental design and conditions, notably the use of considerably higher concentrations of EGCG in the latter investigation, 87 which the former team showed to be indeed toxic. 86 In vivo, orally administered EGCG was compared to intra-peritoneal injections of green tea extract 88 or subcutaneously administered R-apomorphine 89 pertaining to its anti-oxidative action and tyrosine hydroxylase (TH)-positive neuron-sparing effect in MPTP-treated mice. In particular, EGCG prompted the recovery of DA levels and turnover rates, and restored SOD and catalase activities. 88 Interestingly, EGCG did not inhibit monoamine oxidase B (MAO-B) activity in mice at the concentrations used, thus proving that its protective effects are not observed due to impaired transformation of MPTP to its active metabolite MPP+, which requires the enzyme. 88
EGCG mixed in diet improved the survival rate of iron and/or paraquat-exposed Drosophila melanogaster. 90 In particular, EGCG was able to up-regulate mRNA levels of anti-oxidant enzymes such as SOD, 81,88 catalase, 88 and GPx. 81 EGCG's anti-oxidative actions often resulted in the modulation of apoptotic factors such as caspases, 83,86,91,92 Fas ligand (FasL), 92 growth arrest and DNA damage protein (Gadd45), 92 B-cell lymphoma 2 (Bcl-2) family members, 51,86,89,91,92 second mitochondria-derived activator of caspases (Smac), 51 poly(ADP-ribose) polymerase-1 (PARP-1), 84 and release of cytochrome c. 84 Other pathways involving protein kinase C (PKC), 86,89 extracellular-signal-regulated kinases (ERK), 86 and the Akt/glycogen synthase kinase-3 (GSK-3) axis 84 were modulated by EGCG treatment. Remarkably, inhibition of PKC was able to reverse the beneficial effects of EGCG in SH-SY5Y cells challenged with 6-OHDA. 86 Noteworthy, oral administration of EGCG was able to rescue climbing activity in iron and/or paraquat-treated Drosophila 90 as well as motor behavior in 6-OHDA–lesioned rats. 93 However, it is important to mention that in this 6-OHDA rat model, DAergic lesions were not improved by EGCG, which implies that locomotor improvement was seemingly independent of TH-positive neuron protection. 93
Neuroinflammation
It is widely accepted that excessive neuroinflammation is a key pathological contributor in PD. The most common paradigm to study inflammation makes use of lipopolysaccharide (LPS), a pro-inflammatory component of Gram-negative bacteria outer membranes that upon activating Toll-like receptor 4 (TLR4) causes conspicuous inflammatory changes. However, parkinsonian toxins such as MPTP have also been used to induce neuroinflammation. Earlier studies have enlightened EGCG's ability to prevent the activation of nuclear factor-kappa B (NF-κB), 94 a pro-inflammatory transcription factor, offering a good basis to studying its anti-inflammatory effects in DAergic neuronal degeneration. In macrophages treated with LPS, the most important effect of NF-κB inhibition was the reduction of expression of iNOS. 94 This enzyme is responsible for NO production, a key mediator in neuroinflammation.
In primary microglia isolated from 1- or 2-day-old Sprague Dawley rats and further treated with LPS, EGCG was able to counteract inflammatory processes. 95 In addition, LPS-activated primary microglia pre-treated with EGCG whose conditioned medium (CM) was transferred into wells containing SH-5YSY DAergic cells or primary rat mesencephalic cultures were incapable of inducing neuronal degeneration due to impaired secretion of pro-inflammatory factors. 95 In vivo, intra-peritoneal injections of EGCG for 7 days were found to be robustly anti-inflammatory in LPS-treated rats. 96 In addition, a single EGCG intra-peritoneal injection prevented microglial activation in MPTP-treated mice, as shown by diminished expression of cluster of differentiation molecule 11B (CD11b) expression and reduced the number of CD11b-positive microglia, a marker of microgliosis, which correlated with the rescue of TH-positive neurons in the SNc, 97 i.e., neurons expressing a key enzyme upstream in the DA production pathway.
Another team administered EGCG by gavage or a green tea infusion during MPTP injections in mice spread throughout 5 days and found that both treatments were anti-inflammatory, paralleled with prevented loss of TH-positive neurons in the SNc. 98 In particular, EGCG was able to prevent neuroinflammatory mechanisms at several levels, including decreasing the expression, transcription, and activity of neuronal nitric oxide synthase (nNOS, the non-inducible form), 98 and iNOS 99 , as well as inhibiting the secretion of NO 96 and the pro-inflammatory cytokine tumor necrosis factor-α (TNF-α). 95,96,98,86 Interestingly, rotenone toxicity in murine mesencephalic primary neurons and striatal slice cultures was only partially prevented by EGCG co-treatment, by diminishing NO production, but had no effect on neurite length and number, DAergic neuron survival, and superoxide anion formation. 100
Protein aggregation
Alpha-synuclein protein misfolding and formation of β-sheet—rich amyloid fibrils or aggregates are related to cellular toxicity and decay in PD. EGCG is one of the most expansively studied polyphenols regarding its fibril-destabilizing potential, not only in PD but also in Alzheimer's disease 101,102,103 and type 2 diabetes. 104 An early influential study reported that EGCG interfered with an early step in the fibrillization cascade by binding directly to natively unfolded alpha-synuclein and amyloid-beta polypeptides, thus inhibiting their aggregation and redirecting them into an alternative nontoxic “off pathway.” 105 Further studies showed that EGCG inhibits toxic aggregation of amyloidogenic alpha-synuclein in a variety of in vitro models. 101,106 –108 It is thought that EGCG interferes in β-sheet folding on account of hydrophobic and π stacking interactions, but also maybe by direct covalent bonding via the formation of Schiff bases. 109 EGCG may then promote the formation of new kinds of oligomers that were found to be non-toxic to mammalian HEK-293 cells and PC12 cells. 101,105 In particular, EGCG was able to impair the membrane destabilizing capacities of preformed fibrils. 106 –108
Although EGCG has been shown to be an efficient inhibitor of membrane permeabilization by fibrils, Caruana and colleagues pointed out that the tri-hydroxylated rings appear to confer this polyphenol a lower activity compared to di-hydroxylated counterparts. 107 In addition to binding natively unfolded protein species, EGCG was also found to stabilize V30M mutant transthyretin (TTR), the protein responsible for familial amyloidotic polyneuropathy (FAP), in its folded tetramer conformation. 110,111 Indeed, EGCG in vitro as well as orally administered in mice was shown to inhibit TTR fibril formation as well as aggregation by stabilizing folded TTR tetramers. EGCG's fibril-destabilizing effects were further tested in vivo in MPTP-treated mice following oral administration of the compound and, indeed, EGCG was able to prevent alpha-synuclein accumulation. 89 More recently, in Drosophila expressing human alpha-synuclein, ECG, a close tea polyphenol relative of EGCG, mixed in the diet was found to increase life span, recover loss of climbing ability, reduce lipid peroxidation, and mitigate death of brain cells. 112 Furthermore, a mixture of green tea polyphenols, containing several catechins although mainly EGCG, orally administered to MPTP-lesioned cynomolgus monkeys was found to significantly reduce alpha-striatal synuclein oligomers, while improving motor deficits and increasing the number of nigral TH-positive neurons, levels of DA and its metabolites in the striatum. 113
Mitochondrial homeostasis
Lately, mitochondrial dysfunction in PD has gained much attention due to the role of risk genes, such as parkin, DJ-1, PINK1, and LRRK2, in mitochondria homeostasis. The metabolic power player AMPK is a molecular crossroad responsible for directly or indirectly mediating the broad variety of cellular effects of polyphenols, such as resveratrol 114 and quercetin, 115 in the CNS. AMPK “perceives” disturbances in intracellular ratios of ATP:AMP, thus acting as an energy sensor and guardian of mitochondrial homeostasis in several organs, including the brain. 116 Interestingly, orally administered EGCG was shown to suppress mitochondrial dysfunction in DAergic neurons of parkin-null and mutant LRRK2 transgenic Drosophila in an AMPK-dependent manner. 117 Indeed, genetic inactivation of AMPK abolished EGCG's neuroprotective effects, which included promoting the survival of TH-positive neurons and improving the climbing activity of parkin-knockout Drosophila. 117
Direct beneficial effects of EGCG treatment on mitochondrial function were observed in PC12 cells, whereby the mitochondrial transmembrane potential (Δψm) was recovered after paraquat 82 or hydrogen peroxide 85 challenges. In addition, EGCG has been shown to hamper cytochrome c release by damaged mitochondria in vitro. 84,106 Noteworthy, EGCG was found to activate AMPK in hepatocytes in vivo, which correlated with an increase in autophagic flux. 118 This study hints at a possible role for EGCG in autophagy-mediated protein aggregate clearance dependent on AMPK activation. Research conducted in this particular field is still at a preliminary stage but seems very promising in uncovering possible mechanisms underlying EGCG mediated neuroprotection.
Discussion
Accumulating evidence supports a neuroprotective role for EGCG in PD on the basis of its anti-oxidative, anti-inflammatory, and fibril-destabilizing effects. EGCG was shown to cross the BBB and to be well tolerated in humans. Of course, obstacles still remain regarding its rapid metabolization in the liver, although several new initiatives in drug delivery research have shown promising results for increasing the bioavailability of EGCG. Among these, encapsulation of EGCG in nanoparticles or nanolipidic capsules was found to increase uptake of the polyphenol in vitro as well as in vivo while enhancing its bioactivity. 119 –121
Despite the great scientific interest in the neuroprotective properties of EGCG and the promising actions of this polyphenol confirmed by pre-clinical studies, reviewed in this paper, translating these efforts to clinical studies has been very limited. As previously mentioned, there exists a single clinical trial testing green tea polyphenols in de novo PD patients (NCT00461942) that ended in 2011 and results were non-conclusive. While epidemiological observations lend neuroprotective virtues to green tea, 122 data accumulated as of yet are insufficient to conclude that EGCG can really act as a disease-modifying agent in PD.
In the last 10 years, new paradigms in drug administration have emerged. One of these is multi-drug therapy whose development was warranted to overcome obstacles in stand-alone treatments such as low efficacy, resistance, and undesirable side effects. Owing to their wide spectrum of activity, the use of polyphenols and their combination in treating multiple aspects of diseases is a particularly interesting alternative to western medicine. Their synergistic actions arise from mechanisms such as: (1) Regulation of either the same or different targets in various pathways, (2) regulation of transporter enzymes responsible for intestinal absorption or renal clearance and regulation of detoxifying enzymes in the liver leading to increased bioavailability, and (3) overcoming resistance mechanisms in microorganisms or cancer cells. 123 Lately, EGCG has been used together with other substances as a multi-drug treatment, and new interesting synergistic actions have emerged. In MPTP-treated mice, subliminal doses of EGCG and rasagiline, an irreversible inhibitor of MAO B, acted synergistically to restore DAergic neurons of the nigrostriatal axis. 124 Moreover, a study conducted in humans showed that EGCG is an inhibitor of hepatic cathechol-O-methyl transferase (COMT)-mediated O-methylation of levodopa, meaning that oral administration of EGCG may have a significant beneficial effect in PD patients treated with levodopa and carbidopa by reducing their breakdown in the liver. 125 These recent findings may arouse future interest for investigating the clinical potential of EGCG in combination with already existing PD drugs.
Conclusions
In this review, we have provided information on the potential beneficial role of EGCG, the most abundant polyphenolic compound found in C. sinensis L. and tea prepared from this plant's leaves, in relieving DAergic neurodegeneration in vitro and in vivo with regard to the multiple mechanisms underlying PD. It can be concluded that EGCG's anti-oxidant, anti-inflammatory, and fibril-destabilizing effects confer on this polyphenol a broad spectrum of activities that warrant further investigation in the goal to tackle the multi-dimensional causes of PD. Although it is now apparent that EGCG is a promising target from which to develop complementary treatments for PD, much work has yet to be accomplished before its mechanisms of action are fully deciphered. Data accumulated up until today only emphasize the need for well-designed trials in humans to assess the promising effects of EGCG for PD already supported by pre-clinical studies.
Footnotes
Acknowledgments
This work was supported by grants from the National Science and Engineering Research Council of Canada (NSERC) accorded to M.G.M. J.R. is a NSERC-Vanier scholar.
Author Disclosure Statement
No competing financial interests exist.