
Abstract
Consumption of policosanol (PCO), a refined mixture of sugar cane wax alcohols, can elevate serum levels of high-density lipoprotein cholesterol (HDL-C), although the molecular mechanism is still unknown. To investigate the mechanism of action responsible for the anti-senescence effects of PCO on lipoprotein metabolism and HDL functionality, we synthesized reconstituted HDL (rHDL) containing PCO. Encapsulation of PCO by rHDL (PCO-rHDL) enhanced anti-oxidant activity against cupric ion-mediated low-density lipoprotein (LDL) oxidation. PCO-rHDL (final concentration, 9 μM PCO) showed more potent anti-oxidant activity than vitamin C treatment (final concentration, 100 μM). PCO-rHDL inhibited fructose-mediated glycation, which is a major pathological mechanism of diabetic complications, in a dose-dependent manner. PCO also showed cytoprotective effects in monocytes and macrophages with less triggering of apoptotic processes and reactive oxygen species (ROS) production in the presence of hydrogen peroxide (H2O2). PCO-rHDL strongly inhibited uptake of acetylated LDL into macrophages, which is an initial atherosclerotic process. Surprisingly, PCO-rHDL inhibited human serum cholesteryl ester transfer protein (CETP) activity by up to 47% (final concentration, 10 μM PCO). Subcutaneous injection of PCO-rHDL dose-dependently enhanced tissue regeneration activity by 2.4-fold and 3.6-fold compared to that of the phosphate-buffered saline (PBS) control. In conclusion, PCO in HDL showed potent anti-oxidant, anti-glycation, and CETP inhibitory activities along with tissue regenerative activity, especially upon incorporation into HDL. These results suggest that PCO can enhance functionality of HDL in serum to exert anti-senescence and longevity effects.
Introduction
I
PCO is a mixture of aliphatic alcohols ranging from 24 to 34 carbon atoms, namely with octacosanol, triacontanol, hexacosanol, tetracosanol, and tetratriacotanol as major components. Many reports have shown that administration of PCO, especially the Cuban product, for 8 weeks inhibits in vitro cupric ion-mediated LDL oxidation as well as lowers LDL-C and serum triglycerides (TGs) levels. 5 However, the more precise mechanism of action of PCO remains to be investigated, especially with regard to its anti-oxidant activity and elevation of HDL-C.
HDL is a protein and lipid complex in plasma that exerts potent anti-oxidant, anti-inflammatory, and anti-atherosclerotic activities in the context of serum lipid and lipoprotein metabolism. 6 In addition to HDL-C, correlations between HDL structure and function are important to maintaining quality of HDL. 7 Apolipoprotein A-I (ApoA-I), the major protein of HDL, exerts several beneficial activities in HDL. Many researchers, including our own group, have reported that HDL quality is highly dependent on structural and functional correlations of ApoA-I during aging. 8,9 Modification of ApoA-I is directly related to production of dysfunctional HDL, which has more atherogenic and inflammatory properties that exacerbate cellular senescence. 10 Taken together, these reports strongly suggest that the functionality of HDL is highly affected by its composition and particle size. There are two types of HDL, healthy HDL and dysfunctional HDL, and the larger particle size is healthier. 11 In summary, the effects of PCO consumption on the functionality of HDL as well as lipoprotein metabolism and longevity remain to be investigated.
There are several large hurdles to investigating the in vitro physiological functions of PCO, such as its insolubility in water and the selection of an isotonic buffer for enzyme assay, cell-based assay, and animal experiments. To overcome this insolubility, a PCO mixture was incorporated into reconstituted HDL (rHDL) with ApoA-I to evaluate the physiological functions of PCO in lipoprotein metabolism.
Materials and Methods
Materials
PCO–sugar cane wax alcohol was obtained from Rainbow & Nature Pty, Ltd (Sydney, Australia). PCO contains alcohols of several chain lengths, including 1-tetracosanol, 1-heptacosanol, 1-nonacosanol, 1-dotriacontanol, 1-hexacosanol, 1-octacosanol, 1-triacontanol, and 1-tetratriacontanol. Palmitoyloleoyl phosphatidylcholine (POPC, #850457) and cholesterol (C8667) were obtained from Avanti Polar Lipids (Alabaster, AL) and Sigma (St. Louis, MO), respectively.
Synthesis of rHDL
To overcome insolubility of PCO in water and isotonic buffer, we synthesized rHDL containing PCO (PCO-rHDL). rHDL containing PCO was prepared by the sodium cholate dialysis method, as in our previous report, 12 using initial molar ratios of 95:5:1:1 and 95:5:1:5 for POPC:cholesterol:ApoA-I:PCO. The size and hydrodynamic diameter of rHDL particles were determined by 8%–25% native polyacrylamide gradient gel electrophoresis (PAGGE; Pharmacia Phast system, cat. no. 17-0542-01) based on comparison with standard globular proteins (GE Healthcare, Uppsala, Sweden) using the Pharmacia Phast System (GE Healthcare, Uppsala, Sweden).
Circular dichroism and fluorospectroscopy
Average α-helix contents of proteins in lipid-free and lipid-bound states were measured by circular dichroism spectroscopy using a J-700 Spectropolarimeter (Jasco, Tokyo, Japan) located in the Korea Research Institute of Bioscience and Biotechnology (Daejeon, Korea). Spectra were obtained from 250 nm to 190 nm at 25°C in a 0.1-cm path length quartz cuvette at a bandwidth of 1.0 nm, speed of 50 nm/min, and response time of 4 sec. Samples of lipid-free proteins were diluted to 0.07 mg/mL to avoid self-association, whereas lipid-bound proteins were diluted to 0.1 mg/mL. Four scans were accumulated and averaged. The α-helical content was calculated from the molar ellipticity at 222 nm using a mean residue weight of 115.3 for ApoA-I. 13
Fluorospectroscopy
Wavelengths of maximum fluorescence (WMF) of tryptophan residues in wild-type and mutants were determined from uncorrected spectra obtained on an LS55 spectrofluorometer (Perkin-Elmer, Norwalk, CT) using WinLab software package 4.00 (Perkin-Elmer) and a 1-cm path length Suprasil quartz cuvette (Fisher Scientific, Pittsburg, PA). The samples were excited at 295 nm to avoid tyrosine fluorescence, and the emission spectra were scanned from 305 to 400 nm at room temperature.
Purification of human lipoprotein
LDL (1.019 < d < 1.063), HDL2 (1.063 < d < 1.125), and HDL3 (1.125 < d < 1.225) were isolated from the sera of young human males (mean age, 22 ± 2 years, n = 18) who voluntarily donated blood after fasting overnight via sequential ultracentrifugation. The density was appropriately adjusted by addition of NaCl and NaBr, as detailed in our previous report, 14 and procedures were carried out in accordance with standard protocols. 15 Samples were centrifuged for 24 hr at 10°C at 100,000 × g using a Himac CP-90α (Hitachi, Tokyo, Japan) at the Instrumental Analysis Center of Yeungnam University.
Inhibition of glycation in HDL
To investigate the inhibitory ability of PCO against glycation, PCO-rHDL was incubated with
To verify the spectroscopic data, samples were subjected to 15% sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) to compare electromobilities as well as aggregation and multimerization patterns. 18 Gels were then dried and the bands stained with 0.125% Coomassie Brilliant Blue.
Cholesteryl ester transfer assay
rHDL containing ApoA-I and cholesteryl oleate was synthesized in accordance with a previously described method
19
using trace amounts of [3H]-cholesteryl oleate (TRK886, 3.5 μCi/mg of apoA-I; GE Healthcare). rHDL was immobilized using CNBr-activated Sepharose 4B resin (Amersham Biosciences) for easy separation after the reaction in accordance with the manufacturer's instructions. The cholesteryl ester (CE)-transfer reaction was performed in 300-μL reaction mixtures containing each human HDL3 (20 μL, 2 mg/mL) as a cholesteryl ester transfer protein (CETP) source, rHDL-agarose (20 μL, 0.25 mg/mL) as a CE donor, and human LDL (20 μL, 0.25 mg/mL) as a CE acceptor. PCO-rHDL was added to the reaction mixture as an inhibitor. After incubation at 37°C, the reaction was halted via brief centrifugation (10,000 × g) for 3 min at 4°C. Supernatant containing CE acceptor (150 μ L) was then subjected to scintillation counting, and the percentage transfer of [3H]-CE from rHDL to LDL was calculated. The extent of CETP inhibition was calculated as follows:
where the sample is rHDL containing PCO treated as an inhibitor source, and the control is without inhibitor.
LDL oxidation
To compare the extent of oxidation, purified human LDL was incubated with PCO in the presence of 10 μM CuSO4. During incubation, the quantity of conjugated dienes in the form of oxidized product was monitored by measuring the absorbance at 234 nm (Abs234) and 37°C 20 using a Beckman DU 800 Spectrophotometer (Fullerton, CA) equipped with a MultiTemp III Thermocirculator (Amersham, Uppsala, Sweden).
To verify spectroscopic data, oxidized samples were subjected to 0.5% agarose gel electrophoresis to compare electromobilities. 21 Migration of each lipoprotein was dependent on its intact charge and size. Gels were then dried and the bands stained with 0.125% Coomassie Brilliant Blue.
Acetylation of LDL
Acetylation of LDL (acLDL) was performed using saturated sodium acetate and acetic anhydride according to a previously described method. 22 After acetylation and subsequent dialysis, acLDL protein content was determined and filtered through a 0.22-μm filter (Millex; Millipore, Bedford, MA). To detect phagocytosis of acLDL, a fluorescent cholesterol derivative (22-(N-7-nitrobenz-2-oxa-1,3-diazol-4-yl)amino-23,24-bisnor-5-cholen-3-ol [NBD-cholesterol], Molecular Probes N-1148; 70 μg of NBD-cholesterol/mg of ApoA-I) was added to the acLDL particles.
Uptake of acLDL into macrophage
THP-1 cells, a human monocytic cell line, were obtained from the American Type Culture Collection (ATCC; #TIB-202™, Manassas, VA) and maintained in RPMI-1640 medium (Hyclone, Logan, UT) supplemented with 10% fetal bovine serum (FBS) until needed. Cells that had undergone no more than 20 passages were incubated in medium containing phorbol 12-myristate 13-acetate (PMA; final concentration of 150 nM) in 24-well plates for 48 hr at 37°C in a humidified incubator (5% CO2 and 95% air) to induce differentiation into macrophages.
Differentiated and adherent macrophages were then rinsed with warm phosphate-buffered saline (PBS) and incubated with 400 μL of fresh RPMI-1640 medium containing 1% FBS, 50 μL of acetylated LDL (acLDL) (50 μg of protein in PBS), and PCO at its designated concentration for 48 hr at 37°C in a humidified incubator. After incubation, cells were stained with Oil Red O solution (0.67%) to visualize the amounts of lipid species. Cell media (0.2 mL) were then analyzed by the thiobarbituric acid reactive substances (TBARS) assay to evaluate changes in levels of oxidized species using a malondialdehyde (MDA) standard.
Anti-senescence assay
Primary human dermal fibroblasts (HDFs) were cultured, and cellular senescence-associated β-galactosidase (SA-β-gal) activity was compared with the extent of senescence, as described previously. 23 For induction of senescence, cells at passage 9 (at approximately 40% confluence) were exposed to fructose solution for the designated period. To compare inhibition of aging by PCO-rHDL, cellular SA-β-gal activities were compared, as previously described. 24
Zebrafish
Wild-type zebrafish and embryos were maintained according to standard protocols. 25 The maintenance of zebrafish was approved by the Committee of Animal Care and Use of Yeungnam University (Gyeongsan, Korea). The fish were maintained in a system cage at 28°C during treatment under a 10:14 hr light cycle with consumption of normal Tetrabit (Tetrabit Gmbh D49304; 47.5% crude protein, 6.5% crude fat, 2.0% crude fiber, 10.5% crude ash, containing vitamin A [29,770 IU/kg], vitamin D3 [1860 IU/kg], vitamin E [200 mg/kg], and vitamin C [137 mg/kg]; Melle, Germany).
Imaging of reactive oxygen species
After injection of rHDL, changes in levels of reactive oxygen species (ROS) in larvae were imaged by dihydroethidium (DHE; cat. no. 37291; BioChemika) staining as described previously. 26 Images were obtained by fluorescence observation (Ex = 588 nm and Em = 605 nm) using a Nikon Eclipse TE2000 microscope (Tokyo, Japan).
Fin regeneration
Enhancement of tissue regeneration by PCO was tested using an streptozotocin (STZ)-induced adult zebrafish diabetes model, according to our previous report. 27 Experimental zebrafish approximately 12-weeks-old were anesthetized by submersion in 2-phenoxyethanol (Sigma P1126; St. Louis, MO) in system water (1:1000 dilution). Subcutaneous injection of STZ at the nearby urostyle was carried out with a 25-gauge needle and micro-syringe (SGE, Ringwood, Australia) using 35 μL of 0.3% STZ (Sigma S0130; Sigma) in 5 mM citrate buffer at a dosage of 105 μg of STZ per zebrafish (approximately 300 mg of body weight). Control zebrafish were injected with the same volume of citrate buffer. For fin regeneration studies, STZ-induced zebrafish were anesthetized, and tail fins were cut with a scalpel close to the proximal branch point of the dermal rays within the fin. After amputation, 10 μL of PCO-rHDL (7.5 μg of ApoA-I) was injected into the tail muscle of each zebrafish near the urostyle (n = 9 for each group). Following injection, fish consumed a normal diet and were observed in a 28°C system incubator. Images of regenerating fins from live zebrafish were taken at 24-hr intervals up to 148 hr under a stereomicroscope (Motic SMZ 168; Hong Kong) and photographed using a Motic cam 2300 CCD camera with Image Proplus software version 4.5.1.22 (Media Cybernetics, Bethesda, MD, USA).
Cell survival assay
Human monocytes were cultured in Dulbecco's modified Eagle meadium (DMEM) and maintained at 70% confluency. Cells were treated with either PCO in organic solvent or PCO-rHDL and incubated for 4 hr. After incubation, the extent of apoptosis was measured by Acridine Orange (cat. no. A9231, Sigma) staining and visualized by fluorescence detection (Ex = 502 nm, Em = 525 nm). Production of ROS by cells was observed by DHE staining (Ex = 588 nm, Ex = 605 nm).
Statistical analysis
All data were expressed as the mean ± standard deviation (SD) of at least three independent experiments with duplicate samples. Data were evaluated via one-way analysis of variance (ANOVA) using SPSS (v. 14.0; SPSS, Inc., Chicago, IL), and the differences between the means were assessed using Duncan's multiple-range test. Statistical significance was defined as p < 0.05.
Results
Characterization of PCO-rHDL
PCO-rHDL was well synthesized (Fig. 1) with a particle size of around 102–106 Å, as indicated by the black arrowhead, whereas lipid-free ApoA-I particles were about 57–68 Å in size (Table 1). Native gel electrophoresis revealed that the particle size of rHDL was reduced with increased PCO content (Fig. 1). As PCO content in rHDL increased, PCO-rHDL showed lower α-helicity and a particle size of around 101–102 Å.
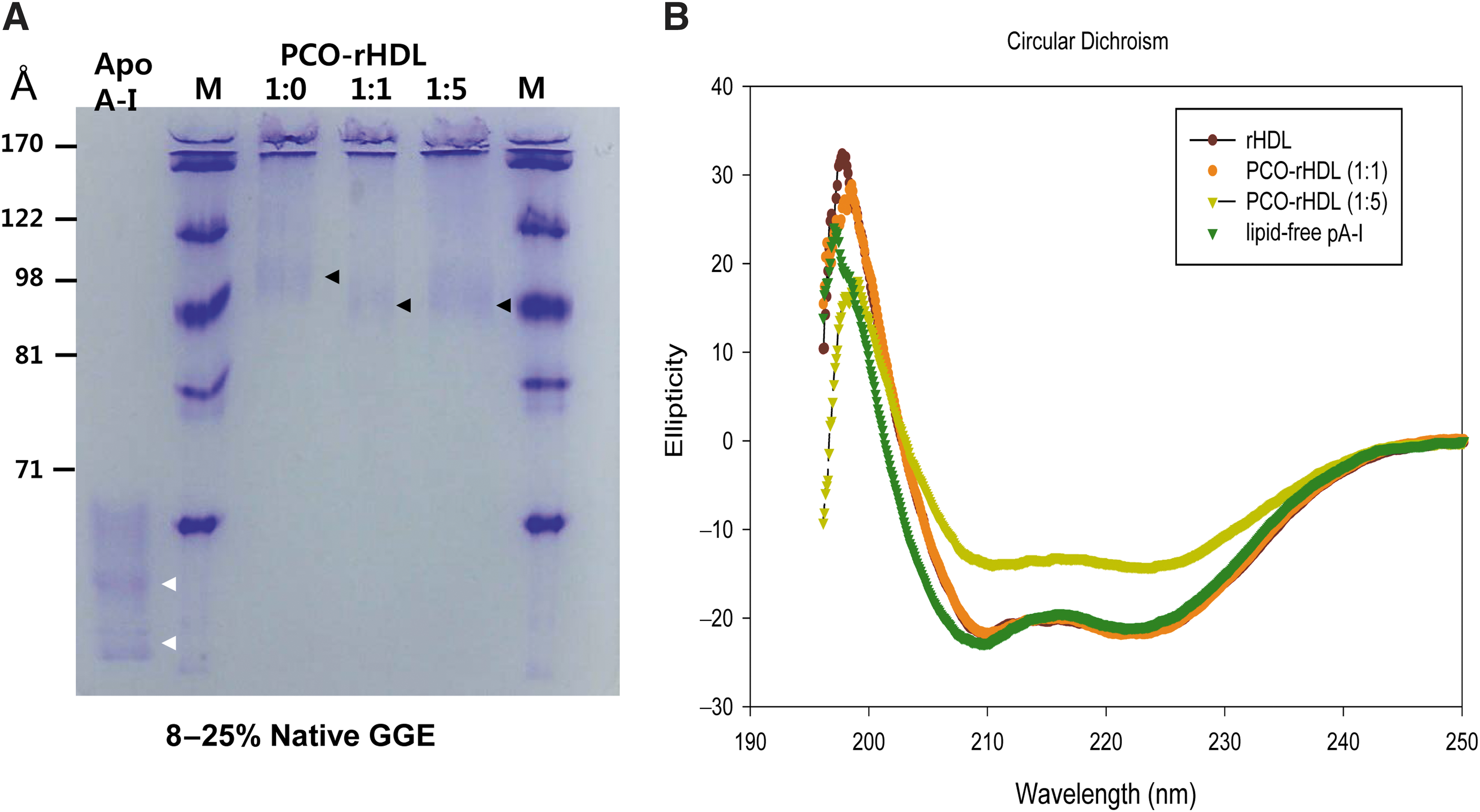
Characterization of reconstituted high-density lipoprotein (rHDL) containing policosanol (PCO). (
Molar ratio was 95:5:1:x for POPC:FC:ApoA-I:policosanol, where x is 0, 1, or 5.
Determined from 8% to 25% native-gradient gel electrophoresis with densitometric scanning analysis.
Determined from circular dichroism spectroscopy.
Determined from fluorospectroscopy to detect tryptophan (Trp) fluorescence.
rDHL, reconstituted high-density lipoprotein; POPC, palmitoyloleoyl phosphatidylcholine; FC, free cholesterol; ApoA-I and A-I, apolipoprotein A-I; PCO, policosanol; WMF, wavelengths of maximum fluorescence.
Fluorospectroscopy revealed that lipid-free and lipid-bound ApoA-I showed WMF values of 350 and 337 nm, respectively, suggesting that tryptophan (Trp) in ApoA-I moved to a more hydrophobic phase via interaction with phospholipids. As PCO content of rHDL increased, the WMF of ApoA-I in PCO-rHDL shifted slightly to red fluorescence (339 and 341 nm at molar ratios of 1:1 and 1:5, respectively). This result indicates that Trp of ApoA-I in PCO-rHDL moved to a more hydrophilic phase due to interactions with PCO and exposure to the hydrophobic phase of ApoA-I. Circular dichroism analysis revealed that lipid-free ApoA-I showed 34% α-helical content, whereas ApoA-I in rHDL showed 74% α-helical content. However, PCO-rHDL showed 67% and 41% α-helical content for (1:1)-rHDL and (1:5)-rHDL, respectively, suggesting that incorporation of PCO reduced the α-helical content of ApoA-I via hydrophobic interactions with the amphipathic domain of ApoA-I.
Anti-oxidant activity of PCO-rHDL
After 6 hr of incubation with cupric ion, LDL was oxidized and showed the fastest electromobility on a 0.5% agarose gel (Fig. 2, lanes 1). However, PCO-rHDL–treated LDL showed slower electromobility, indicating much less oxidation similar to vitamin C– (final 100 μM) treated LDL. More interestingly, after 24 hr of incubation in the presence of Cu2+, vitamin C–treated LDL (lanes 5 and 6) almost disappeared, similar to the Cu2+-treated control (Fig. 2, lanes 1). This result indicates that the anti-oxidant ability of vitamin C almost disappeared after 24 hr. However, PCO in rHDL retained anti-oxidant activity because PCO-rHDL–treated LDL showed a more distinct band with slower mobility than oxLDL (Fig. 2., lanes 1) and vitamin C–treated LDL (Fig. 2, lanes 5 and 6).
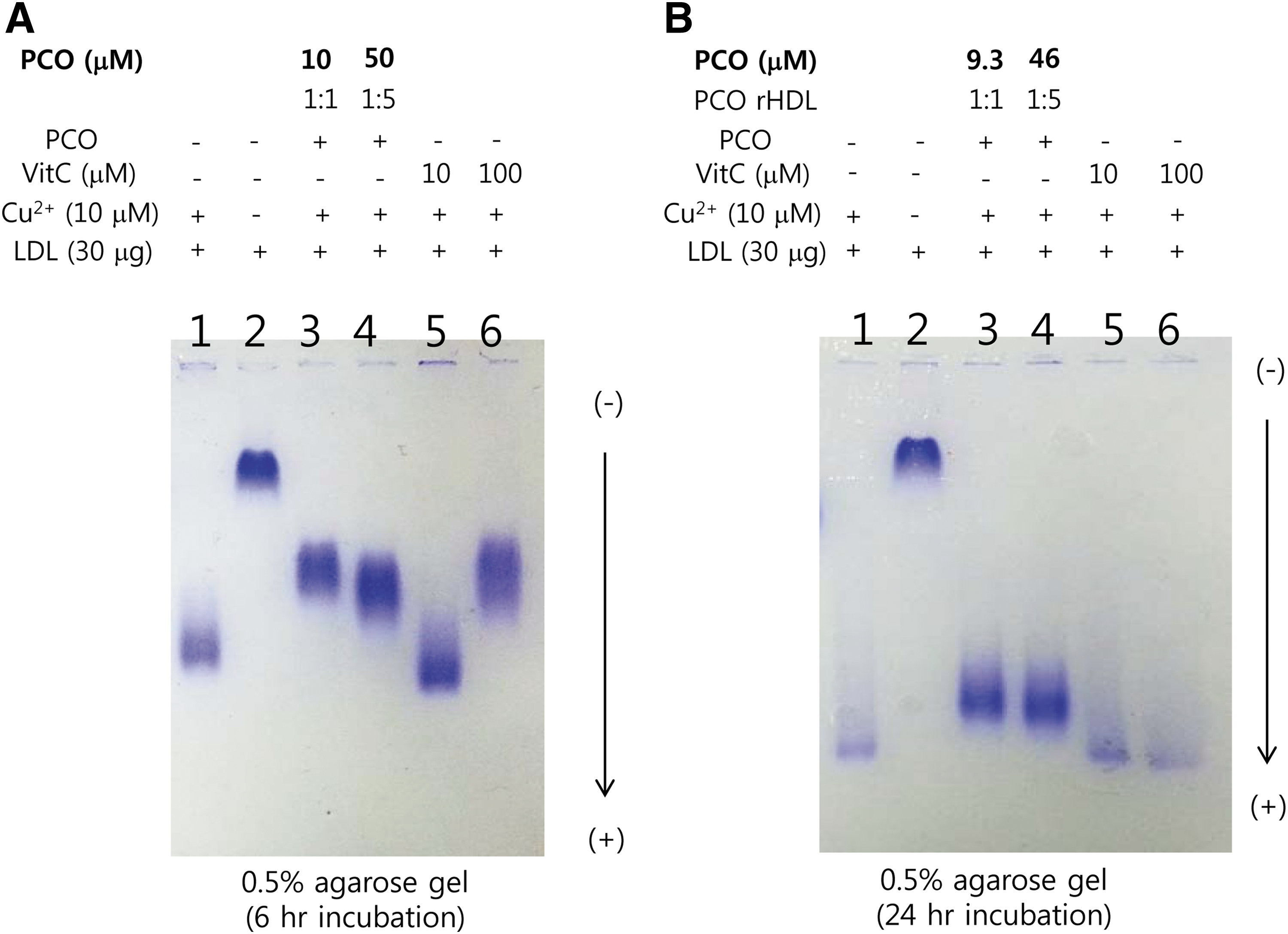
Electromobility of oxidized low-density lipoprotein (LDL) treated with policosanol (PCO) in the presence of cupric ion during 6 hr (
Anti-glycation activity of PCO
Glycation of HDL by fructose treatment caused dysfunctional HDL production. Fructose-treated HDL3 showed an 11-fold increase in glycation compared to HDL alone based on yellow fluorescence (Fig. 3A). Fructose-treated HDL3 showed a smeared ApoA-I band pattern with severe multimerization (Fig. 3B, lane 2), whereas native ApoA-I showed a single band (Fig. 3B, lane 1). In the presence of fructose, PCO treatment prevented glycation in a dose-dependent manner; treatment with PCO (final 9.3 and 230 μM) resulted in 26% and 55% inhibition of glycation with a more distinct ApoA-I band pattern and less multimerization (Fig. 3B, lanes 4–6). PCO-rHDL (final 9.3 and 230 μM) also showed up to 68% and 58% inhibition, respectively. These results suggest that the anti-glycation effect of PCO was due to its association with rHDL via putative protection from Lys and Arg modification induced by fructosylation, which is involved in the Maillard reaction of glycation and protein multimerization.
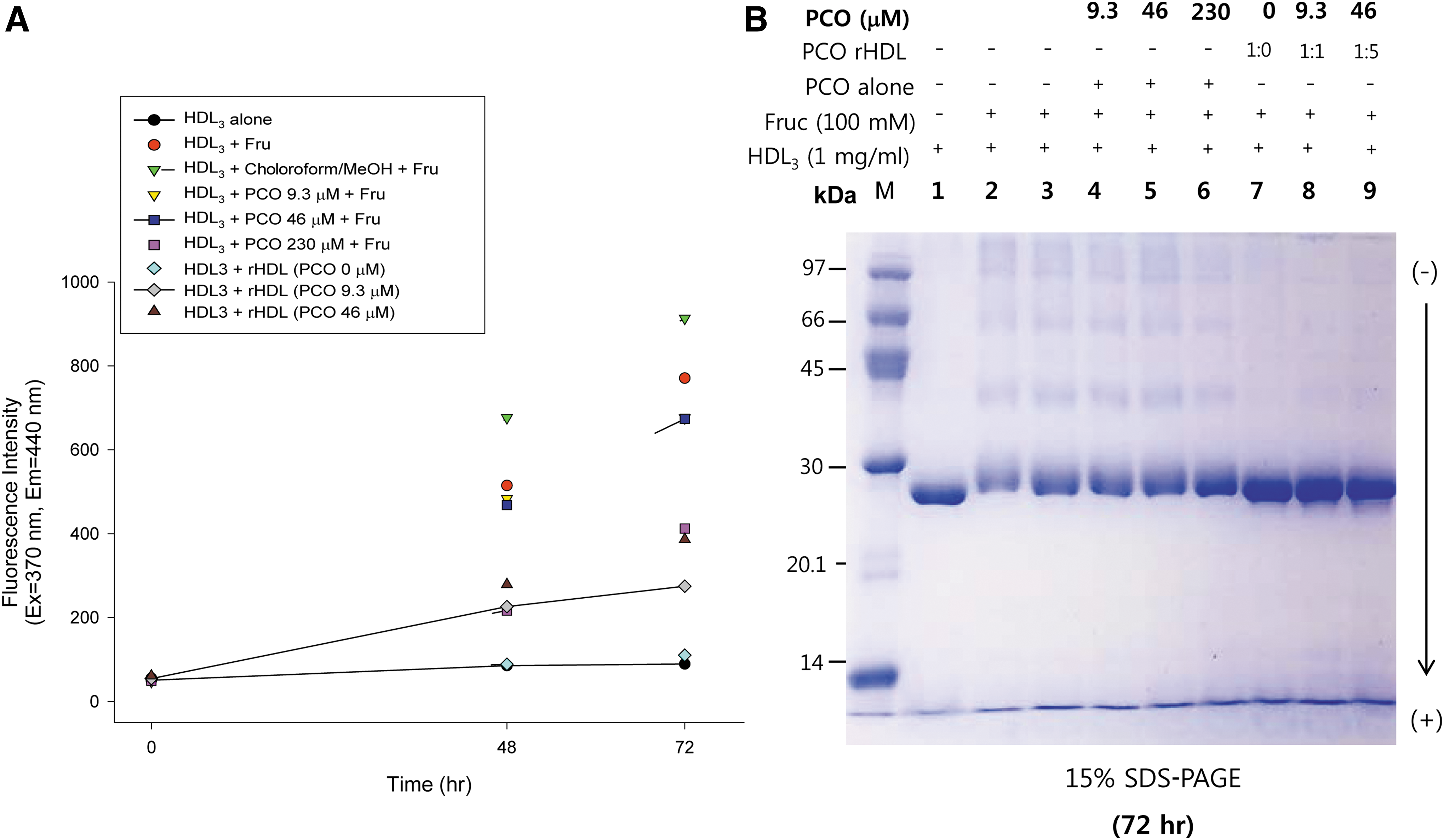
Glycation extent of HDL3 by fructose (Fruc) treatment in the presence of policosanol (PCO). (
Anti-apoptotic activity of PCO
As shown in Fig. 4, organic solvent– (CHCl3:Me-OH) treated cells (Fig. 4Ab) showed six-fold higher apoptosis than control cells (Fig. 4Aa), as visualized by Acridine Orange staining. Solvent-treated cells showed up to 12-fold higher production of ROS (Fig. 4Ab) than control (Fig. 4Aa) via DHE staining. Furthermore, PCO treatment (final 9 to 230 μM) resulted in 71% reduction of apoptosis and up to 82% reduction of ROS production in a dose-dependent manner. Last, PCO (final 230 μM) resulted in up to 82% reduced ROS production compared to solvent treatment.
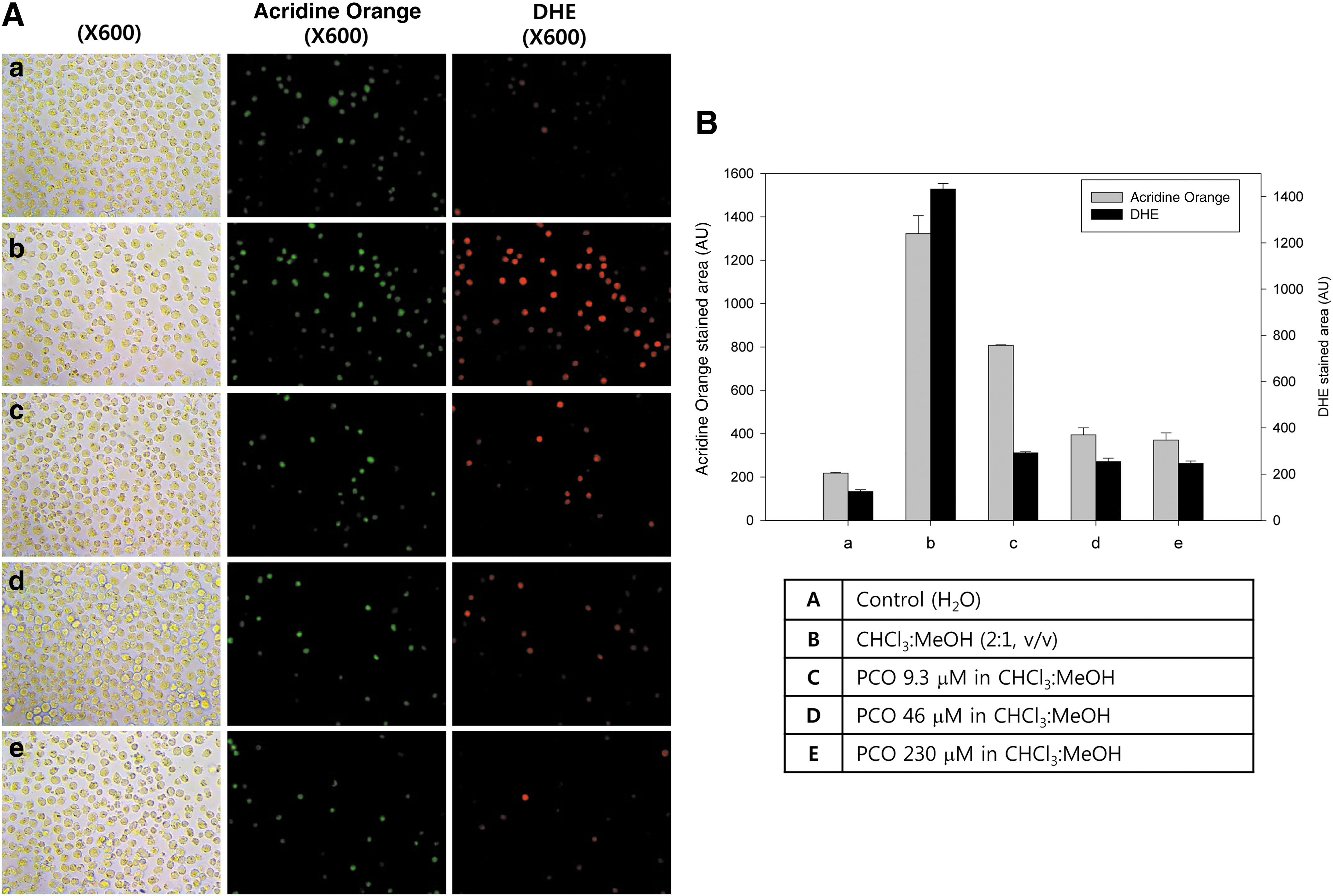
Cytoprotective effect of policosanol (PCO). (
Cytoprotective activity of policosanol against oxidative stress
In the presence of hydrogen peroxide (H2O2) (final 0.03%), monocyte cells showed greater apoptosis and ROS production (Fig. 5) due to oxidative stress. PCO-treated cells showed reduced apoptosis in a dose-dependent manner; 9 μM, 46 μM, and 230 μM (final) treatment resulted in 5%, 22%, and 45% reduced apoptosis, correspondingly, on the basis of green fluorescence. ROS production also decreased with increased PCO dosage; 9 μM, 46 μM, and 230 μM (final) treatment resulted in 38%, 65%, and 70% reduced ROS production, correspondingly, by DHE staining.

Cytoprotective effect of policosanol (PCO) in the presence of hydrogen peroxide (H2O2) (final 0.03%). (
Inhibition of acLDL uptake by PCO-rHDL
As shown in Fig. 6, uptake of acLDL into macrophages was strongly facilitated, as visualized by NBD-cholesterol detection with green fluorescence (Fig. 6Ab), compared to the PBS control (Fig. 6Aa). However, phagocytosis of acLDL was remarkably blocked up to 96% by PCO-rHDL in a dose-dependent manner, whereas rHDL alone caused 85% inhibition.
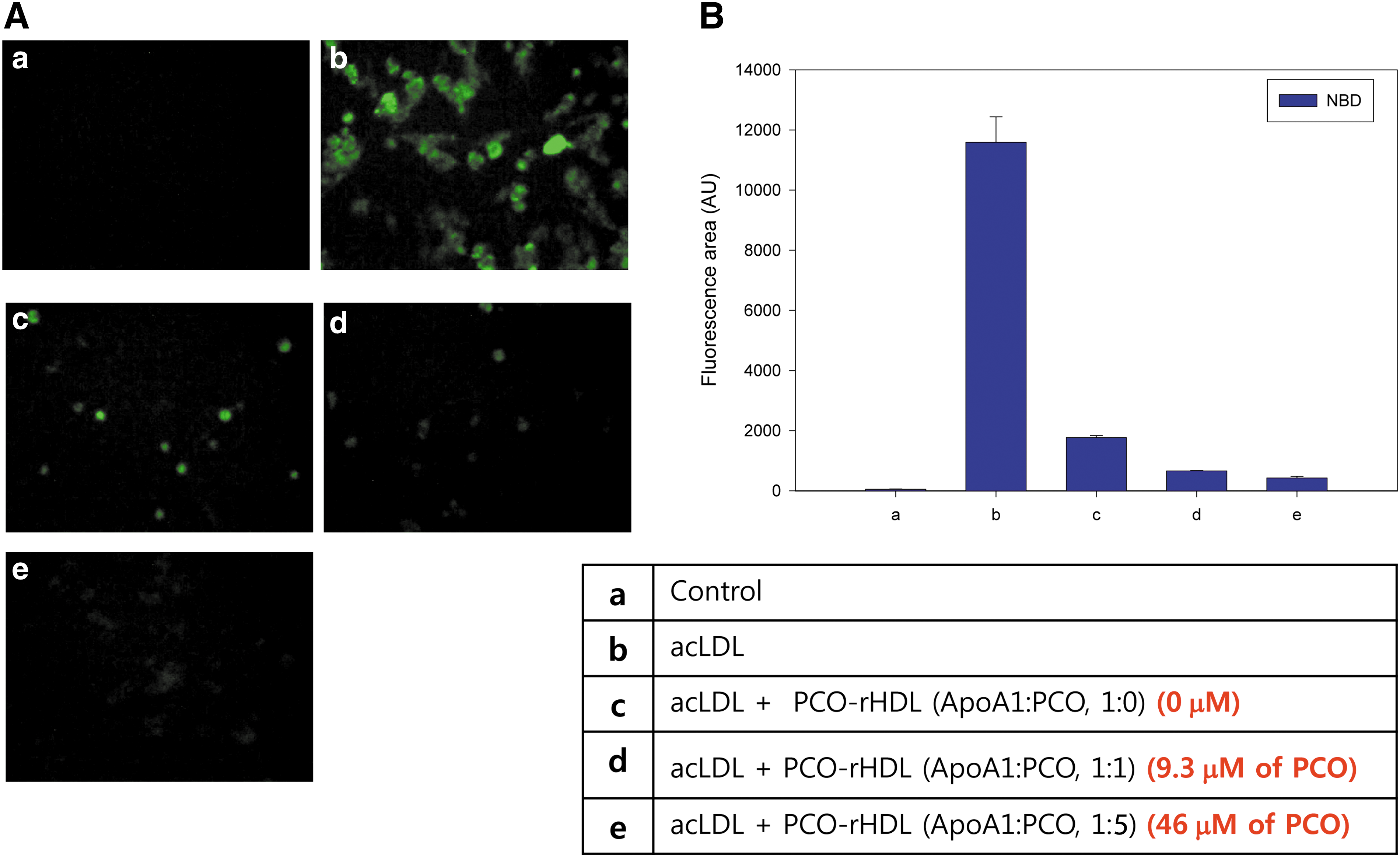
Anti-atherosclerotic effect of policosanol (PCO) in the presence of acetylated low-density lipoprotein (acLDL). (
Anti-senescence activity of PCO in dermal cells
As shown in Fig. 7, organic solvent (CHCl3:Me-OH) caused a three-fold increase in the number of SA-β-gal–positive cells (Fig. 7Ab), but a 42% decrease in overall cell number than the control (Fig. 7Aa), indicating that the organic solvent had strong cytotoxic and pro-senescent effects. However, PCO in solvent dose-dependently inhibited cellular senescence up to 65% and 70% at 9 and 46 μM (final), respectively, along with an increase in cell number, suggesting that PCO had anti-senescent and cell proliferation effects (Fig. 7c, d). Moreover, PCO-rHDL showed a stronger anti-aging effect and greater cellular proliferation in a dose-dependent manner. Compared to the PBS control, PCO-rHDL–treated cells (Fig. 7f, g) showed 68% and 80% reduction of SA-β-gal–positive cells, whereas rHDL-treated cells (Fig. 7e) showed 52% reduction. PCO-rHDL also showed a cell proliferation effect of up to 1.9-fold and 1.6-fold for (1:5)-rHDL and (1:1)-rHDL, respectively, compared to rHDL alone (photo e).

Suppression of cellular senescence in human dermal fibroblasts (HDFs) treated with policosanol (PCO) with or without reconstituted high-density lipoprotein (rHDL). Percentage of senescence-associated β-galactosidase (SA-β-gal)–positive cells per 7.4 mm2 of cell culture area during treatment with rHDL or rapamycin (dissolved in CHCl3:Me-OH, 2:1). (
PCO inhibits CETP activity
ApoA-I and HDL itself can inhibit CE-transfer activity from HDL (CE donor) to LDL (CE acceptor) by 5–10%, as in our previous report. Unexpectedly, PCO-rHDL showed remarkably higher inhibitory ability against CETP activity than rHDL alone; 47% and 40% inhibitory abilities at ApoA-I:PCO molar ratios of (1:1)-rHDL and (1:5)-rHDL, respectively (Fig. 8). Interestingly, CETP inhibition increased at a higher PCO concentration in rHDL. However, final concentrations of 10 and 1.7 μM for PCO and ApoA-I with (1:1)-rHDL, respectively, resulted in higher CETP activity than that of (1:5)-rHDL, which has 50 μM and 1.7 μM concentrations for PCO and ApoA-I. These results indicate that CETP inhibitory activity was more enhanced by incorporation of PCO into rHDL, and a low concentration (10 μM) was sufficient to exert CETP inhibition.
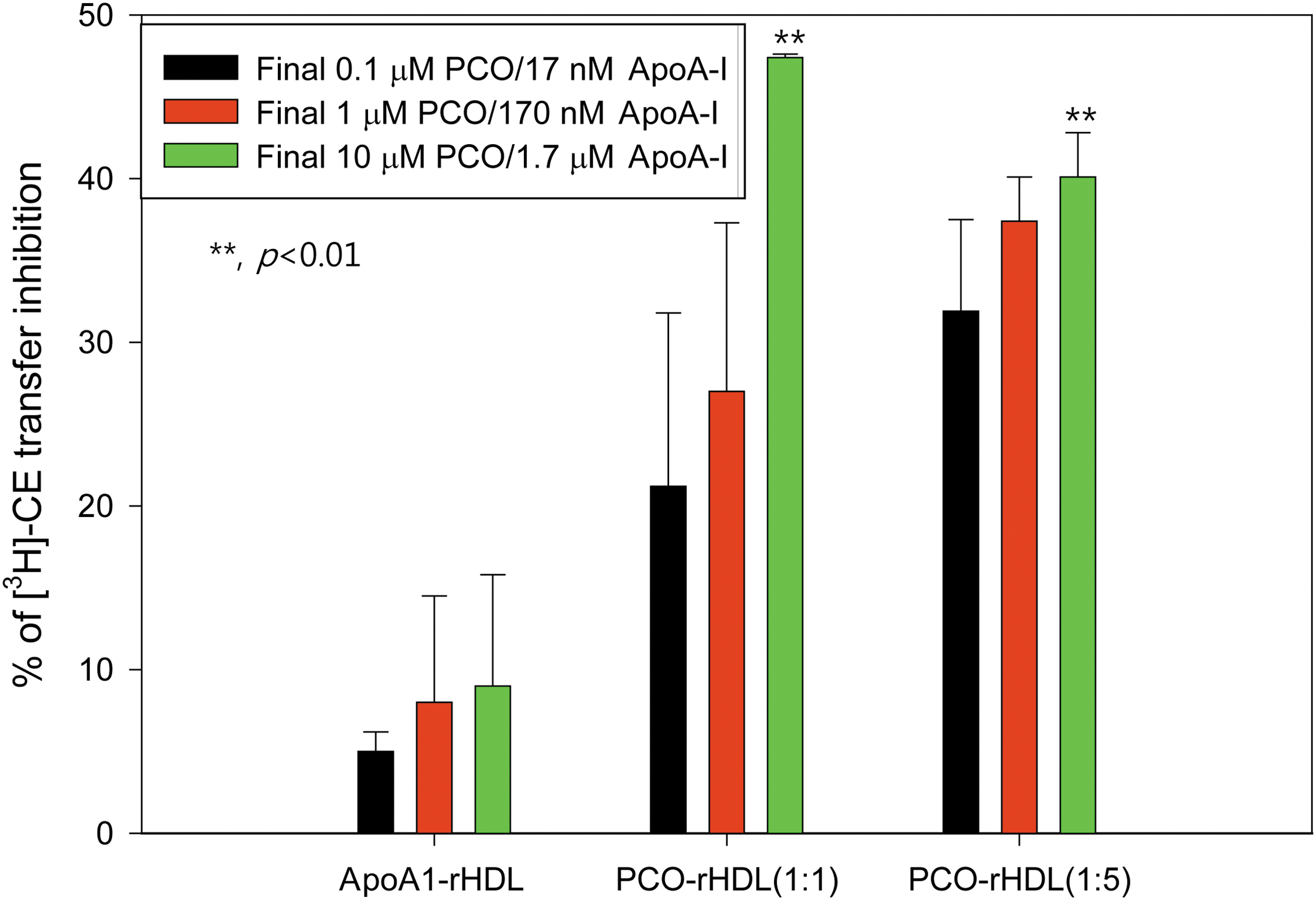
Cholesteryl ester transfer protein (CETP) inhibitory activity of reconstituted high-density lipoprotein (rHDL) containing policosanol (PCO). Data shown are the mean ± standard deviation (SD) of three independent experiments performed in duplicate. Cholesteryl ester (CE) transfer from [3H]HDL (50 μg of apolipoprotein A-I (ApoA-I; 30,000 cpm) to human low-density lipoprotein (LDL) (50 μg of protein) by human HDL3 (25 μg of protein) was inhibited by rHDL containing PCO. Color images available online at
Tissue regeneration
Under normal diet (ND) consumption, the rHDL-injected group showed two-fold higher tissue regeneration activity than the PBS-injected group over a period of 144 hr (Fig. 9A). The PCO-rHDL–injected group showed 2.4-fold and 3.6-fold higher tissue regeneration activity for (1:1)-rHDL and (1:5)-rHDL, respectively, than the PBS control. From DHE staining, ROS production in amputated tissue was reduced by 29% upon rHDL treatment compared to the PBS control. PCO-rHDL–injected zebrafish showed the smallest ROS production in a dose-dependent manner (up to 53% reduction) by DHE staining compared to the PBS control.
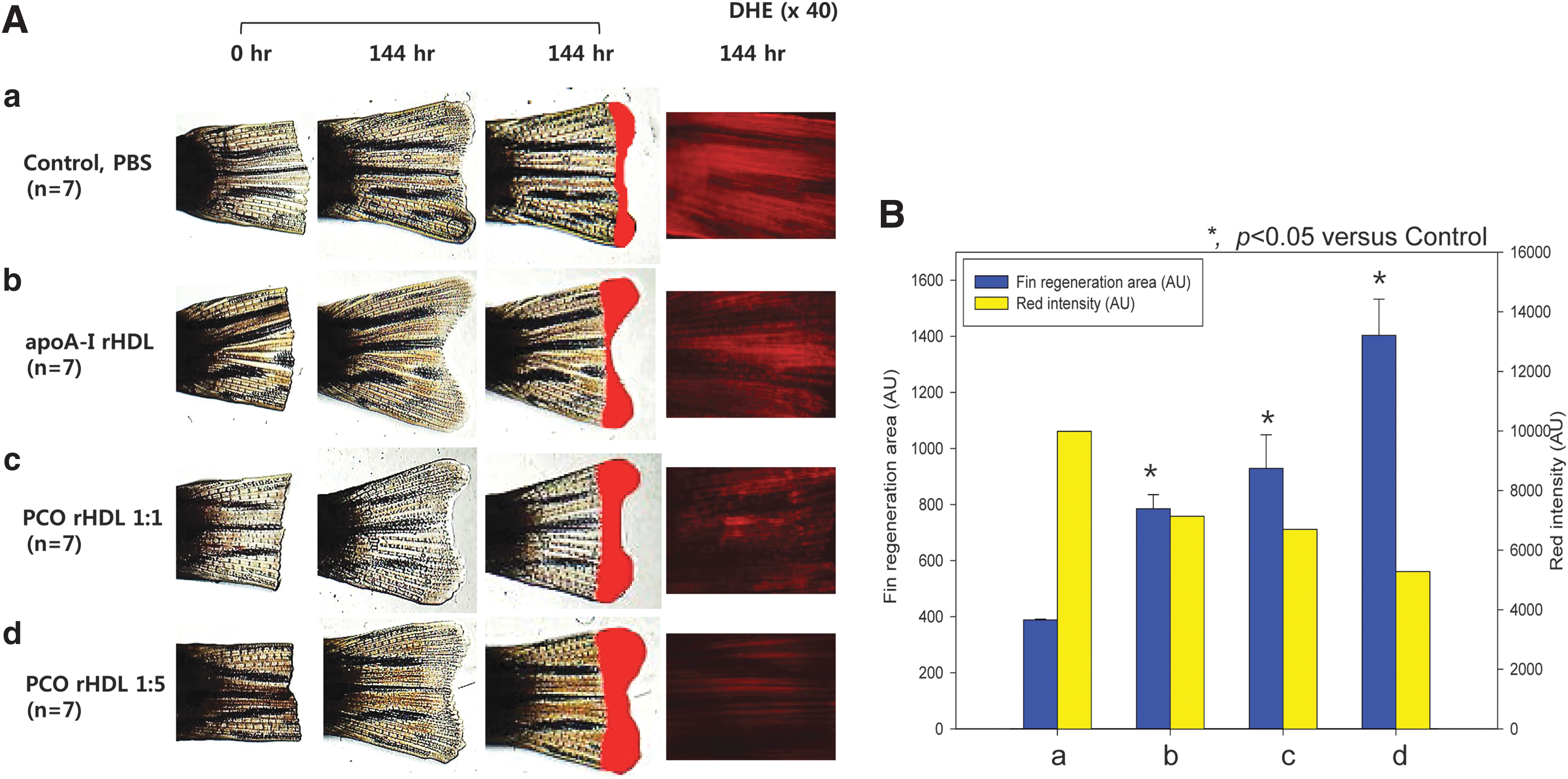
Tissue regenerative activity of policosanol (PCO). Enhancement of zebrafish fin regeneration by injection of reconstituted high-density lipoprotein (rHDL) containing PCO and rHDL (2 μg of apolipoprotein A-I [Apo-A-I]). (
Discussion
It is well known that PCO is a lipid-lowering agent that increases HDL-C levels, although the mechanism of action is unclear. Furthermore, there is controversy surrounding the lipid-lowering effect of PCO, exacerbated by its unknown mechanism. 28,29 Indeed, it is very difficult to elucidate the molecular mechanism of PCO due to its high insolubility in a physiologic aqueous buffer. In the current study, to overcome the insolubility of PCO, we synthesized PCO-rHDL and tested its physiological activity in an aqueous buffer system.
To the best of our knowledge, this is the first report of the molecular mechanism of the anti-aging activity of PCO in an in vitro experiment, cell-based assay, and vertebrate model. Synthesis of PCO-rHDL and particle formation were detected, whereas particle size and α-helicity decreased slightly upon PCO incorporation (Fig. 1). PCO-rHDL showed potent anti-oxidant activity against LDL oxidation (Fig. 2), anti-glycation activity (Fig. 3), and anti-apoptotic activity in monocytes (Figs. 4 and 5). PCO-rHDL also resulted in potent inhibition of oxLDL uptake into macrophages (Fig. 6) and cellular senescence of human dermal fibroblasts (Fig. 7). Finally, PCO-rHDL exerted potent CETP inhibitory activity (Fig. 8) and greater enhancement of tissue regeneration than rHDL alone (Fig. 9).
Although clinical trials suggest that a 10-mg daily dose is sufficient for the lipid-lowering effect of PCO, the reasoning remains unclear. In this study, anti-oxidant (Fig. 2), anti-glycation (Fig. 3), anti-atherosclerotic (Figs. 4 –6), and CETP inhibitory activities of PCO were detected at lower dosages of 10 μM (final) in rHDL. These results might explain why the low dosage of PCO was effective in improving the serum lipid profile.
The results of our in vitro study on suppression of LDL oxidation due to the anti-oxidant activity of PCO (Fig. 2) are in good agreement with previous clinical data. 30 Susceptibility to LDL oxidation in vitro was reduced by 5–10 mg/day of PCO for 8 weeks, indicating that reduction of plasma LDL-C is associated with reduced susceptibility to LDL oxidation. However, these reports cannot explain why PCO has atheroprotective and pro-longevity activities. A previous report from Dullens et al. showed that individually emulsified PCO components with different chain lengths were unable to improve lipid profiles of LDL receptor-deficient mice (LDLr+/− mice). 31 Despite this result, LDLr+/− mice lack serum CETP activity and display remarkably different lipid and lipoprotein metabolism compared with humans. Thus, any conclusion on the basis of results from mice is limited because HDL is the major carrier lipoprotein for cholesterol in mice, whereas LDL is the major lipoprotein to carry cholesterol in humans.
In the current study, PCO was solubilized via incorporation into HDL to elucidate its physiological functions in lipid metabolism. The most important finding of this study is that PCO displayed potent CETP inhibition activity in the context of CE transfer from HDL to LDL. CETP plays a critical role in lipid distribution among lipoproteins and is recognized as an atherogenic factor. 32 Several CETP inhibitors have been developed by big pharmaceutical companies as next-generation drugs for treatment of cardiovascular disease (CVD). Among CETP inhibitors, anacetrapib is currently in phase III clinical development. 33 Treatment with a CETP inhibitor (100 mg of anacetrapib/day) for 52 weeks has been shown to reduce LDL-C by 30% and elevate HDL-C levels two-fold. 34 Treatment with 150 mg of anacetrapib for 8 weeks had the same beneficial effect in the form of a 1.4-fold higher ApoA-I level and 30% lower Apo-B level. 35 In the same context, the current results show that PCO-rHDL possesses much higher CETP inhibitory activity than rHDL alone, which showed 10% inhibition at 1.7 μM ApoA-I. 25 This result suggests that PCO plays a critical role in lipid homeostasis between HDL and LDL via exchange of CE and TGs.
A mutation of CETP, I406V, has pro-longevity effects via elevation of large-sized HDL and HDL-C levels. 36 Low HDL-C levels and prevalence of dysfunctional HDL are risk factors of autoimmune diseases characterized by inflammation, such as vitiligo 37 and systemic lupus erythematosus. 38 PCO contains both hydrophobic and hydrophilic moieties and can thus interact with the hydrophobic active site of CETP, as described previously. 39,40 Thus, inhibition of CETP by PCO might have anti-inflammatory and anti-aging effects, resulting in cellular replication (Figs. 4 –7) and tissue regeneration (Fig. 9). These results might explain why PCO consumption is associated with increased HDL-C and enhanced HDL functionality. Enhanced HDL functionality is associated with anti-apoptotic (Fig. 5), anti-atherosclerotic (Fig. 6), anti-senescent (Fig. 7), and tissue regeneration activities (Fig. 9). Similarly, rHDL containing V156K (V156K-rHDL) shows enhanced functionality that stimulates more insulin secretion and tissue regeneration 27 as well as CETP inhibition. 23
Other in vitro and animal studies have reported that PCO inhibits cholesterol synthesis via modulation of 3-hydroxy-3-methylglutaryl-coenzyme A (HMG-CoA) reductase 41 possibly by activating adenosine monophosphate (AMP)-kinase. 42 However, there is no direct evidence of inhibition of HMG-CoA reductase activity. PCO is insoluble in water, thus it is very difficult to assess its function in vitro. However, our current results clearly indicate that PCO has potent CETP inhibitory activity and an anti-aging effect. These results help explain the mechanism of action of PCO and its rejuvenation activity via modulation of CETP activity. To the best of our knowledge, this is first report on the beneficial functions of PCO at the molecular level of lipoprotein metabolism.
In conclusion, PCO enhances the beneficial functions of HDL to maximize its anti-oxidant, anti-glycation, and anti-atherosclerotic activities along with inhibition of CETP. These results may shed more light on the association between PCO activity and HDL functionality to enhance longevity.
Footnotes
Acknowledgments
This work was supported by the Mid-carrier Researcher Program (2014-11049455) and the Medical Research Center Program (2015R1A5A2009124) through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT and Future Planning.
Author Disclosure Statement
All authors indicate that there are no financial interests relevant to this article and there are no potential conflicts to disclose.