
Abstract
Our recent studies have shown that Huogu (HG) formula was effective both in clinic experience and in experimental osteonecrosis of the femoral head (ONFH). Given that defective of bone marrow stromal cells (MSCs) contribute to the development of osteonecrosis and MSCs show enormous potential in the treatment of ONFH, especially to aging people. How HG impacts the differentiation of MSCs and what is the underlying cellular and molecular mechanism remains largely unknown. Here, we found that an aqueous fraction of HG (HGA) significantly increased the alkaline phosphatase (ALP) activity, mineralized nodules, and migration of MSCs in a dose-dependent manner. Meanwhile, HGA could enhance the mRNA and protein expression of Runt-related transcription factor 2 (Runx2), Alp, Bmp2, osteocalcin (Ocn), and Osterix (Osx). Further investigation of the molecular mechanisms revealed that HGA treatment obviously increased expression, secretion, and activation of bone morphogenetic protein (BMP) 2 and β-catenin, two key regulators of the BMP or Wnt signaling pathway. Furthermore, osteogenic differentiation of MSCs could be blocked by using pharmacological inhibitors for these signaling pathways such as Noggin and Dkk-1. Besides, HGA could inhibit adipogenic differentiation of MSCs. Our study reveals that HGA promotes the osteogenesis of MSCs via the BMP and Wnt signaling pathways. Our findings provide mechanistic insights into the role of HG in treating ONFH.
Introduction
O
Traditional Chinese Medicine has been extensively used in the treatment of ONFH for centuries. Huogu (HG) formula, as a Chinese decoction, has been reported to be effective for the experimental ONFH. 10,11 Moreover, it shows good clinic effects and has obvious advantages compared with surgery due to its fewer side effects. 12 In spite of the clinical applications, the molecular mechanisms underlying the HG formula functions are largely unknown. Studies have revealed that the HG formula is able to regulate the key factors of osteogenesis and lipid metabolism by intervening bone morphogenetic protein (BMP)/Smads and Wnt signaling pathways in vivo. 10,11,13 Given that BMP/Smads and Wnt signaling pathways play an important role in MSCs differentiation, we speculated that the HG formula may have some influence on MSC differentiation, which contributes to its protection on ONFH.
To identify the main component of HG, our previous results have shown that the aqueous fraction, named HGA, was the most effective fraction in the HG extract that accounted dominantly for its pro-osteogenic action. 14 In this study, we demonstrate that the HGA is able to stimulate the osteogenic differentiation and inhibit the adipogenic differentiation of MSCs, and it exerts its effect on MSCs through activating BMP and Wnt signaling pathways. Our findings establish the mechanistic basis for the role of the HG formula in its therapeutic application for ONFH.
Materials and Methods
Preparation of HG
HG formula is composed of 12 traditional drugs, as indicated in Table 1. The drugs were purchased from Tong-Ren-Tang Ltd. All drugs except Cervi Cornus Colla were boiled twice in water [1:10(w/v) for 1.0 hours each time], then filtered, and condensed. The concentrated water solution was deposited by adding 95% ethanol to make the whole solution as 60% ethanol solution. Cervi Cornus Colla was dissolved in hot water and merged into the solution just mentioned. After 48 hours (at 4°C), the supernatant liquor was centrifuged and condensed in vacuum to a proper amount. The final concentration of HG is 1.91 g (crude herbs)/g liquid extract. Fifty grams of the liquid extract was dissolved in H2O to make 100 mL and was extracted with four different solvents successively. Each combined solution and the remaining water solution were concentrated in vacuum to gain five separated parts as an aqueous fraction (HGA, 29.38 g), petroleum ether fraction (HGP, 0.26 g), chloroform fraction (HGC, 0.08 g), ethyl acetate fraction (HGE, 0.12 g), and n-butanol fraction (HGB, 1.14 g), respectively.
Qualitative high performance liquid chromatography analysis
For high performance liquid chromatography (HPLC) analysis, HGA was dissolved in distilled water to 0.294 g/mL. Standards of 5-hydroxymethylfurfural, chlorogenic acid, albiflorin, paeoniflorin, ferulic acid, liquiritin, acteoside, ligustilide, cinnamaldehyde, lobetyolin, cinnamic acid, and benzoylpaeoniflorin were purchased from the National Institute for Food and Drug Control (Beijing, China). The purity of all compounds was more than 98%. All other chemicals were of analytical grade. The solutions were filtered through 0.22 μm membranes before HPLC analysis.
HPLC was performed on a Shimadzu HPLC System (Shimadzu Corporation). An Agilent TC(2) C18 column (250 × 4.6mm, 5 μm) was maintained at 35°C. Detection wavelength was set at 254 nm. The mobile phase for fingerprint analysis consisted of methanol (A) and 0.1% formic acid (B) at a flow rate of 1.0 mL/min. A gradient program was used as follows: 0–40–70–80 minutes, 5%–32%–66%–100% A; the injected volume was 10 μL. Online UV spectra were recorded in the range of 190–800 nm.
Bone marrow cell isolation and culture
Sprague–Dawley rats (4-week-old) were housed in the Laboratory Animal Center (China Academy of Chinese Medical Sciences, Beijing, China) according to the guidelines for the care and use of laboratory animals of the NIH and the China Academy of Chinese Medical Sciences. All experimental procedures were approved by the Committee for Animal Use of the China Academy of Chinese Medical Sciences. Bone MSCs were isolated from rats as previously described. 15 Briefly, MSCs were obtained from the femurs and tibias by flushing with 5 mL complete medium DMEM-HG (Gibco) with 10% fatal bovine serum (FBS; Gibco) and 1% penicillin/streptomycin (Gibco) and incubated at 37° with 5% CO2. Cells from the fourth passage were used in the subsequent assays.
Osteogenic and adipogenic differentiation
MSCs were seeded as a monolayer in a 25 cm2 flask. For osteogenic differentiation, MSCs were cultured in growth medium until they were 50% confluent, and osteogenic differentiation was induced by osteogenic induction medium (OIM), which is DMEM with 0.1 μM dexamethasone (Sigma Aldrich), 10 mM β-Glycerophosphate, 50 mg/L ascorbic acid, and 10% FBS. For adipocyte differentiation, cells were cultured in the adipogenic induction medium (AIM) according to the manufacturer's instructions (ScienCell). The medium was changed every 2–3 days, and the culture was maintained for 7–21 days for different assays.
Alkaline phosphatase staining
After MSCs were treated with or without OIM and HGA (25, 50, and 100 ng/mL) for 7 days, cells were washed with phosphate-buffered saline (PBS) and fixed with 10% formalin for 15 minutes. The fixed cells were subjected to an assay for alkaline phosphatase (ALP) activity by the BCIP/NBT Alkaline Phosphatase Color Development Kit according to the manufacturer's protocol (Beyotime). The images were taken with a digital camera attached to the microscope. Image-Pro Plus software version 6.0 (IPP6.0; Media Cybernetics, Inc.) was used to calculate the ALP activity that was indicated as the integrated optical density (OD) of staining in the unit area. Then, ALP buffer was added and quantified using the microplate reader (Bio-Rad) at 405 nm. All procedures were conducted in triplicate.
Alizarin red S staining for mineralization measurement
After MSCs were treated with or without OIM and HGA (25, 50, and 100 ng/mL) for 14 days, cells were washed with PBS and fixed with 10% formalin for 15 minutes. Alizarin red S staining was performed according to the manufacturer's protocol (Sigma Aldrich). Differentiated cells containing mineral deposits were stained with Alizarin red S solution. Orange red staining indicated the position and intensity of calcium deposits. The images were captured and analyzed by IPP 6.0. The stained cells were dissolved in 10% cetylpyridinium chloride (CPC; Sigma) and quantified using a microplate reader (Bio-Rad) at 550 nm. All tests were conducted in triplicate.
Oil Red O staining
After MSCs were treated with or without AIM and HGA (25, 50, and 100 ng/mL) for 14 days, cells were fixed with 10% formalin for 15 minutes. Then, they were washed with 50% ethanol and stained with 0.2% Oil Red O (Sigma Aldrich) solution for 20 minutes at room temperature. After washing of Oil Red O stained cells with PBS, the extraction of the Oil Red O dye was performed by using extraction buffer [50% EtOH, 2% (w/v) SDS] for 15 minutes at room temperature. Quantification of the Oil Red O dye was performed at OD 595 using an ELISA reader (Bio-Rad). All tests were conducted in triplicate.
Enzyme-linked immunosorbent assay
After MSCs were treated with or without OIM and HGA (25, 50, and 100 ng/mL) for 14 days, mediums were collected and analyzed for β-catenin and BMP2 using the enzyme-linked immunosorbent assay (ELISA) kit (R&D System) according to the manufacturer's instructions.
Cell survival viability assay
Cell viability was determined by 3-(4,5-dimethyl-2-thiazolyl)-2,5-diphenyl-2H-tetrazolium bromide (MTS) method using CellTiter 96® AQueous One Solution Cell Proliferation Assay (Promega) according to the manufacturer's instructions. The experiments were carried out three times in triplicate measurements.
Wound-healing migration assay
For the wound-healing migration assay, MSCs were seeded onto 24-well plates at a density of 1 × 105 cells/well for 24 hours, and the confluent monolayer of culture was scratched with a fine pipette tip. Migration was visualized under an inverted microscope. The rate of wound closure by HGA and control medium was observed at 24 hours, respectively.
RNA isolation and quantitative real time-polymerase chain reaction
Total RNA was isolated from cultured MSC by using TRIzol Reagent (Life). cDNA was synthesized from extracted RNA using Taq-Man RT reagents according to the manufacturer's instructions (Applied Biosystems). SYBR green I was used to monitor amplification of DNA on the Cycler thermal cycler and the real-time quantitative polymerase chain reaction (PCR) detection system (Bio-Rad). The primer sequences for Runt-related transcription factor 2 (Runx2), Alp, Bmp2, osteocalcin (Ocn), and Osterix (Osx) are listed in Table 2. Gapdh was utilized as an internal control to normalize for differences in the amount of total RNA in each sample. DNA amplification was conducted by an ABI Prism 7500 real-time PCR system (Applied Biosystems). The experiments were carried out three times in triplicate measurements.
PCR, polymerase chain reaction.
Western blotting
After MSCs were cultured with or without OIM and HGA (25, 50, and 100 ng/mL) for 14 days, cell pellets were lysed in RIPA lysis buffer (50 mM Tris–HCl pH 7.4, 300 mM NaCl, 5 mM EDTA, 0.5% Triton X-100, 1 mM NaF, 1 mM Na3VO4, and protein inhibitor cocktail) for 30 minutes on ice. Lysates were centrifuged at 12,000 g for 20 minutes at 4°C. The lysates containing 80 μg of protein were separated on 10% SDS–PAGE gels and then electrotransferred onto a PVDF transfer membrane. The membranes were immunoblotted with primary antibody for β-catenin (Cell Signaling Technology), Runx2 (Cell Signaling Technology), OCN (Santa Cruz), and Osx (Abcam) in Tris-buffered saline and Tween 20 containing 1% BSA overnight at 4°C and afterward blocked in 5% nonfat dry milk in Tris-buffered saline. Horseradish peroxidase-conjugated anti-mouse and anti-rabbit antibodies (1:5000) were used as secondary antibodies. The membranes were then visualized using the ECL system (Pierce). GAPDH as an internal control (Cell Signaling Technology) was used to confirm equal protein loading.
Immunofluorescence staining
MSCs were seeded onto glass coverslips and incubated with HGA 100 ng/mL in the absence or presence of Noggin or DKK-1 for 2 days. The cells were fixed in 4% paraformaldehyde in PBS and then permeabilized with 0.1% Triton X-100 in PBS for 15 minutes. Then, the cells were incubated with primary antibody for β-catenin (Cell Signaling Technology) overnight and immunostained with Alexa 594-conjugated secondary antibodies (Molecular Probes, Inc.). Finally, the coverslips were mounted with Hoechest 33258 (Vector Laboratories) mounting media and visualized under confocal microscopy with an Olympus ×41 microscope.
Statistical analysis
All raw data were processed by authorized software SPSS 13.0. The data were analyzed by one-way ANOVA followed by Dunnett's t-test to assess the statistical significance of the differences between the study groups. Differences were considered statistically significant when p was less than 0.05.
Results
HPLC analysis of HGA
The HPLC profile at 254 nm could exhibit the whole chromatographic characteristic of HGA that was established by optimizing the chromatographic separated conditions. By comparison of the retention times and on-line UV spectra of the reference standards, 11 compounds were identified (Fig. 1). According to the chemical investigations of herbs of HGA, Acteoside and 5-hydroxymethylfurfural were attributed to Rehmanniae radix praeparata. Paeoniflorin and benzoylpaeoniflorin were owing to Paeoniae radix rubra. Ferulic and ligustilide were from Angelicae sinensis radix and Chuanxiong rhizome. Liquiritin obviously belonged to Glycyrrhizae radix et rhizoma. Cinnamaldehyde and cinnamic acid originated from Cinnamomi ramulus, and lobetyolin was ascribed to Codonopsis radix. This chemical information from HPLC analysis was preliminary and elucidated the main chemical composition of HGA. However, the content of 11 compounds was low, which needed to be further studied.
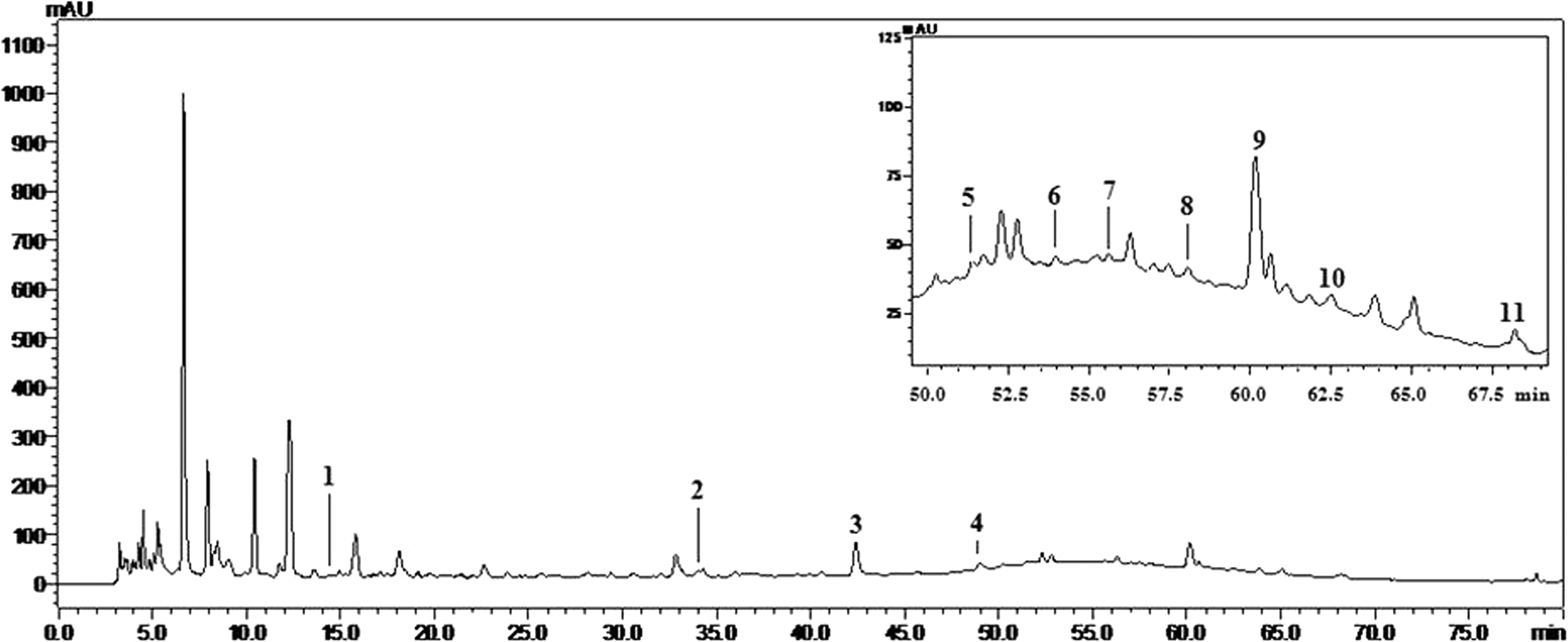
HPLC profile and the identified compounds from aqueous fraction of HGA (UV chromatograms at 254 nm). Peak 1, 5-hydroxymethylfurfural. 2, chlorogenic acid. 3, paeoniflorin. 4, ferulic acid. 5, liquiritin. 6, acteoside.7, ligustilide. 8, cinnamaldehyde. 9, lobetyolin. 10, cinnamic acid. 11, benzoylpaeoniflorin. HPLC, high performance liquid chromatography.
HGA stimulates osteogenic differentiation of MSCs
To determine the effect of HGA on osteogenic differentiation, MSCs were differentiated in OIM with or without HGA for up to 14 days. The differentiation of MSCs was evaluated by ALP staining at day 7, and calcium deposition by Alizarin red S staining at day 14 separately. A serial of paper reported 16,17 that 17β-estradiol (E2) could distinctly enhance the osteogenic activity, so we selected E2 as the positive control in our present experiment. As shown in Figure 2, HGA treatment significantly increased ALP activity in a dose-dependent manner (p < 0.05 or 0.01). At 100 ng/mL of HGA, the ALP activity was highly increased by 2.05 fold when compared with OIM alone. Consistently, AR-S staining and quantification analysis showed that calcium deposition at the end of differentiation was much more in the HGA-treatment group than in the group cultured in OIM alone (p < 0.05 or 0.01, Fig. 3), showing dense red color in the HGA-treated group and light red color in the OIM alone group, and maximal effect was observed at a concentration of 100 ng/mL. Its osteogenic effect was similar to the positive control E2 at 10−6 M.

HGA enhances the osteogenesis of MSCs by ALP staining.
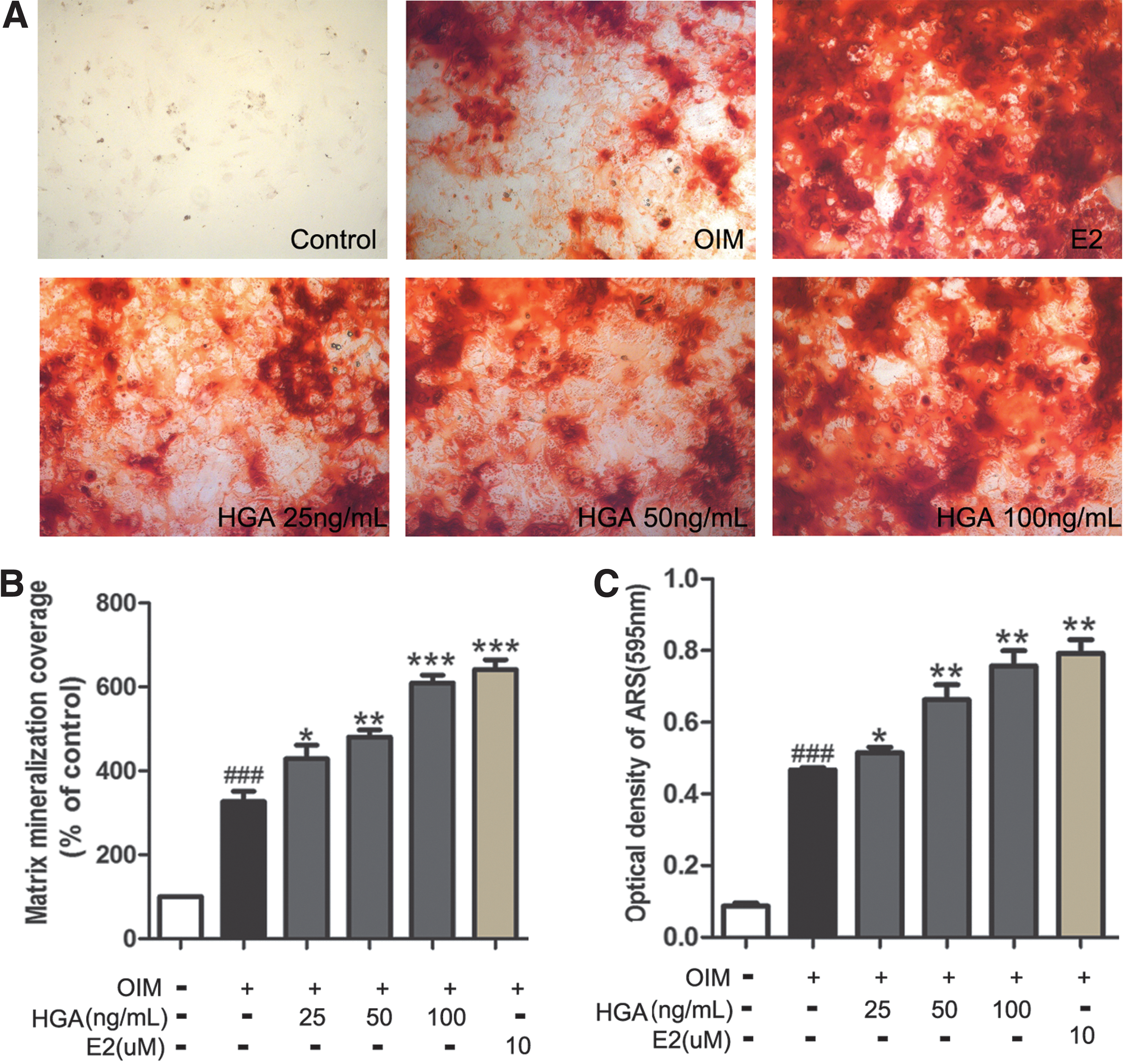
HGA increases mineralization in MSCs cultured in OIM.
Since osteoblastic differentiation is a complicated process, including proliferation and migration of MSCs, the proliferative and migratory activity of HGA decoction was also evaluated in this experiment. As shown in Figure 4, HGA markedly promoted MSCs proliferation as assessed by MTS assay. Meanwhile, the migration ability of MSCs was tested by the wound-healing assay. The results revealed that HGA significantly enhanced the migration of MSCs (Fig. 4C).

HGA promotes viability and migration of rat MSCs.
HGA increased expression of osteogenesis-related genes
Marker genes and transcription factors, including Runx2, Alp, Bmp2, Ocn, and Osx, were significantly increased in their expression during the osteogenic differentiation. To further confirm the effect of HGA, we detected the gene expression of Runx2, Alp, Bmp2, Ocn, and Osx. The real-time PCR results showed that all the tested genes were significantly up-regulated by HGA in a dose-dependent manner (Fig. 5A–E). Of all the earlier five genes, Bmp2 was the most obviously affected by HGA, reaching 6.35 fold at 100 ng/mL compared with OIM only.
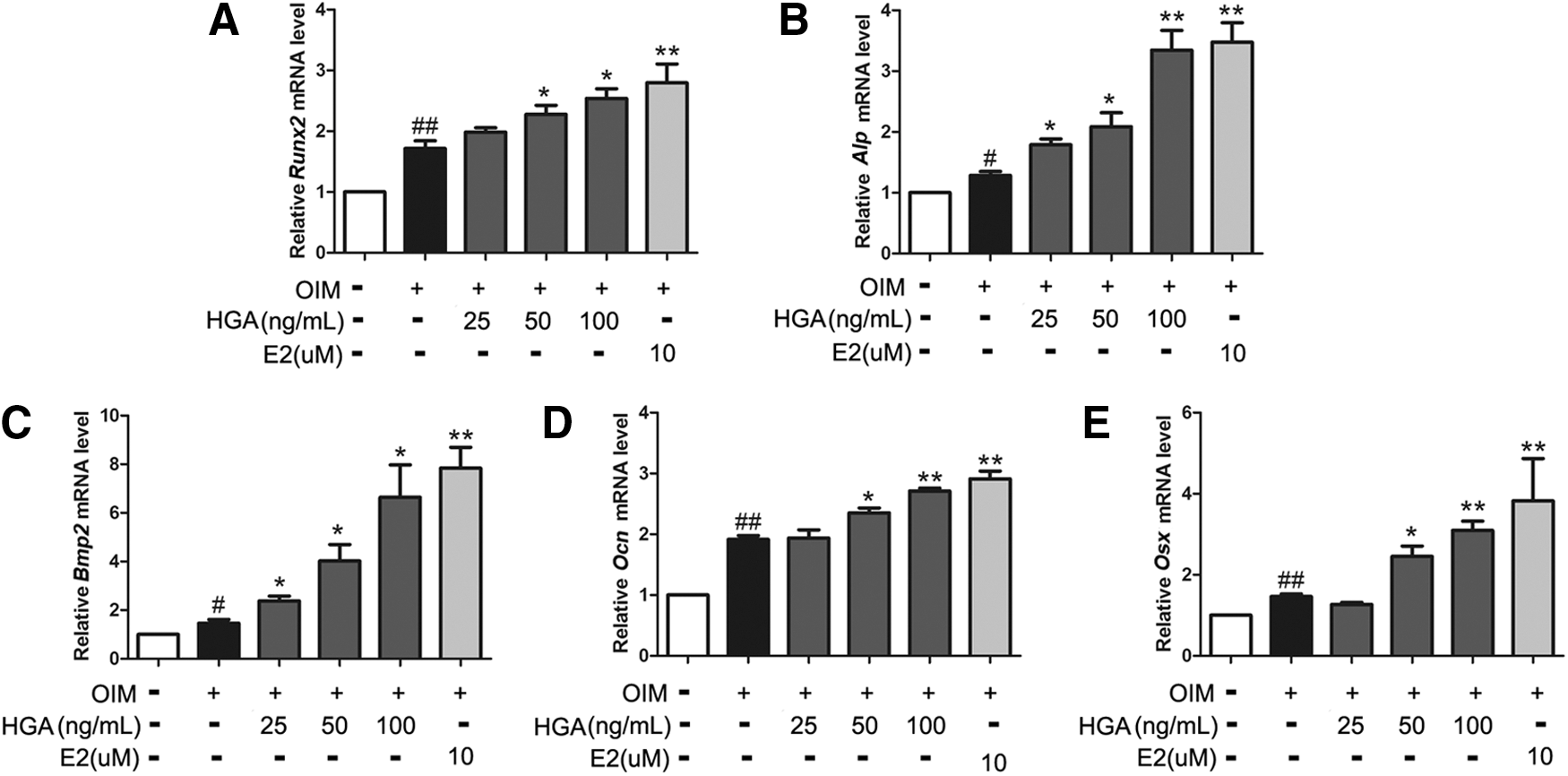
HGA up-regulated mRNA expression levels of Runx2
BMP and Wnt signaling pathways are involved in the effects of HGA on the differentiation of MSCs
BMP and Wnt signaling pathways have been shown to play an important role in the osteogenic differentiation of MSCs. 18 –20 Given that HGA enhanced osteogenic differentiation of MSCs, we were intrigued to determine whether BMPs and Wnt signaling pathways contribute to the function of HGA. The level of the key signal moleculars such as BMP2 and β-catenin in the medium was analyzed by ELISA when MSCs were treated with or without HGA. As shown in Figure 6, treatment of MSCs with HGA resulted in a dose-dependent increase of BMP2 and β-catenin in the medium compared with that in the absence of HGA.
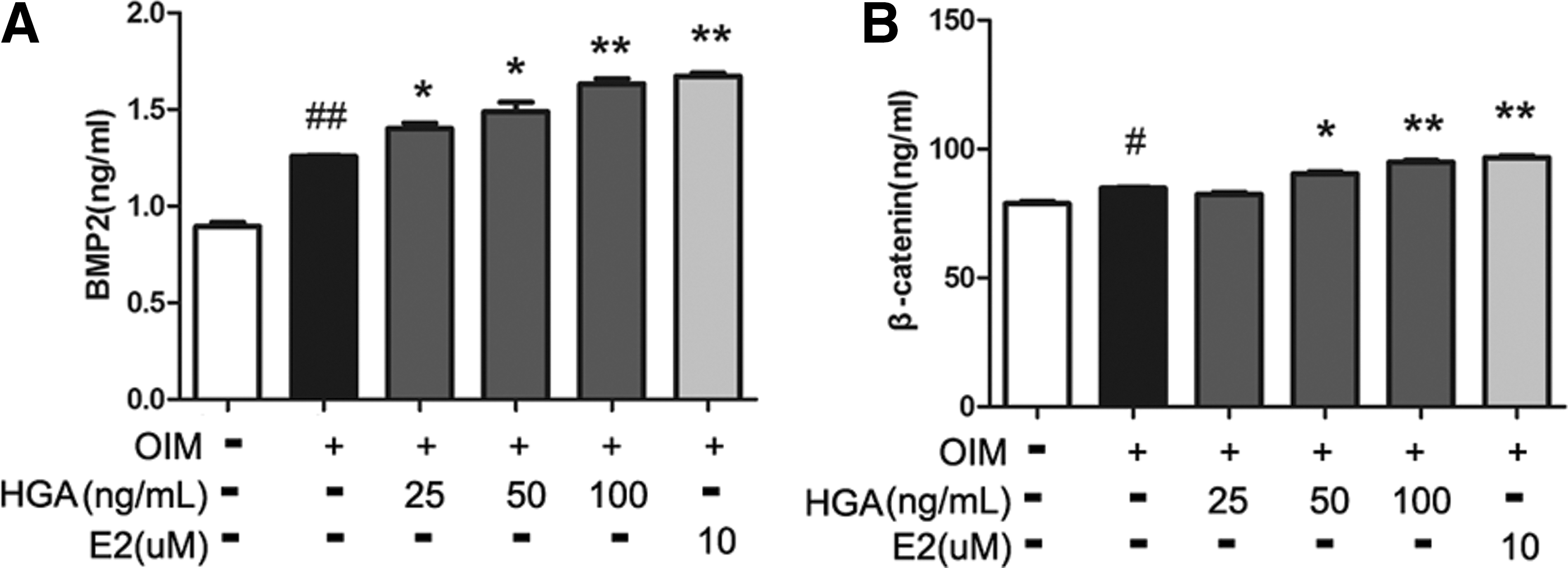
HGA increased the secretion of BMP2 and β-catenin by MSCs into the medium. MSCs were incubated in exposure to HGA (25, 50, 100 ng/mL) in OIM for 14 days. The BMP2
Noggin and Dkk-1 inhibit the effect of HGA on osteogenesis in MSCs
The data cited so far have suggested the possible involvement of BMP and Wnt signaling pathways in HGA-promoted osteogenesis of MSCs. To further prove the involvement of BMP and Wnt signaling pathways during this process, Noggin (an inhibitor of BMPs) and Dkk-1 (an inhibitor of Wnt) were applied to block the activation of BMP and Wnt signaling pathways. As shown in Figure 7A–C, the treatment with the inhibitors for BMPs and Wnt could almost completely abolish HGA-induced osteogenesis of MSCs tested by Alizarin Red S staining. However, the total protein of the MSCs treated with or without HGA and/or inhibitors remain almost unchanged (Supplementary Figure S1, Supplementary Data are available online at
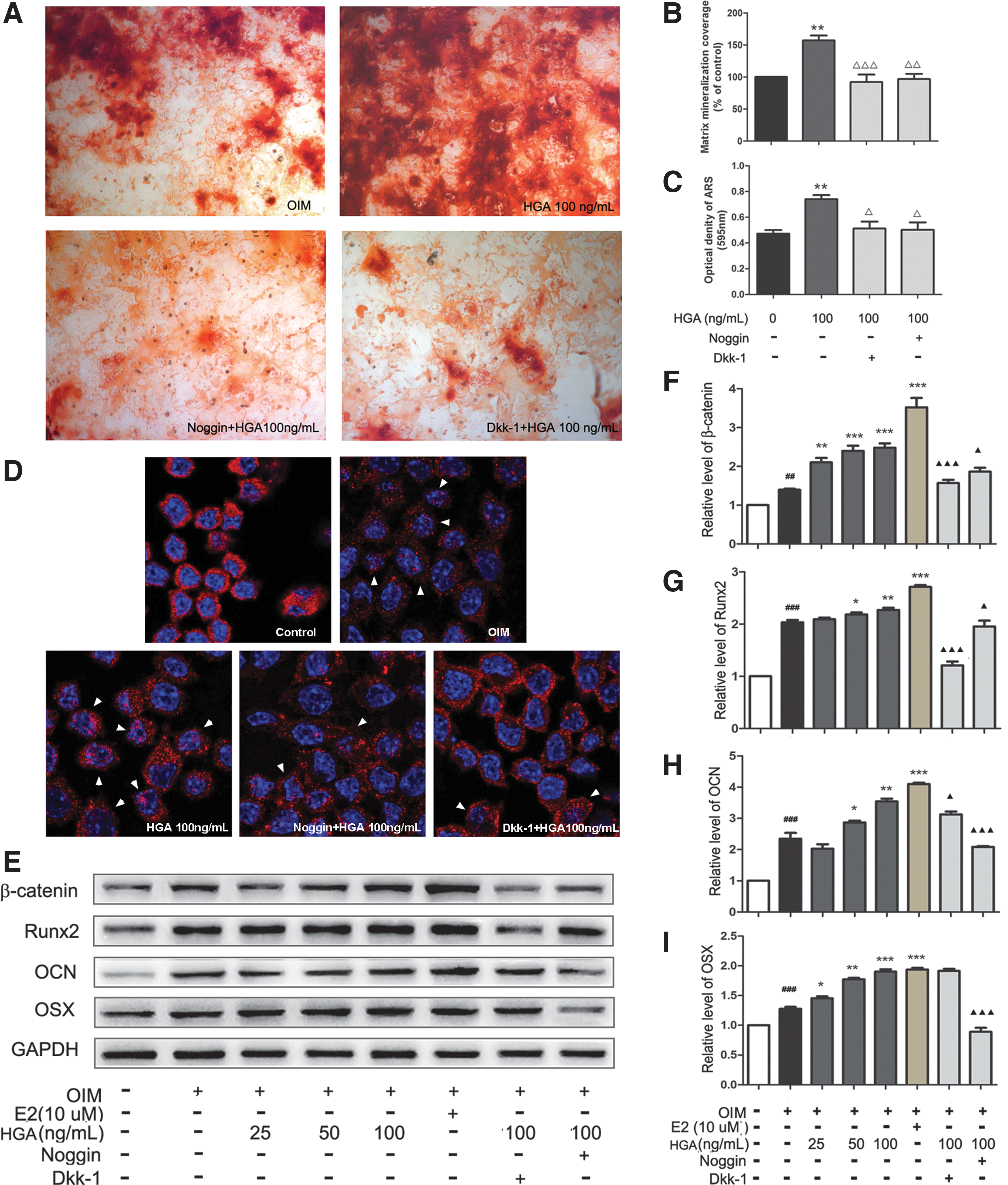
Noggin and Dkk-1 reversed HGA-mediated regulation of osteogenesis of MSCs.
HGA suppresses adipogenic differentiation of MSCs
There is a reciprocal relationship between the differentiation of adipocytes and osteoblasts of MSCs in the bone marrow. 22 Besides the osteogenesis, we also investigate the effect of HGA on adipogenic differentiation in rat MSCs. Oil Red O staining was used to visualize the formation of adipocytes after 14 days of adipogenic induction. As shown in Figure 8, HGA significantly inhibited the formation of adipocytes. At 100 ng/mL of HGA, the number of adipocytes was decreased by 54.3% (p < 0.01). Its effect was similar to the positive control Sim in decreasing the number of adipocytes by about 57.9% (p < 0.01).
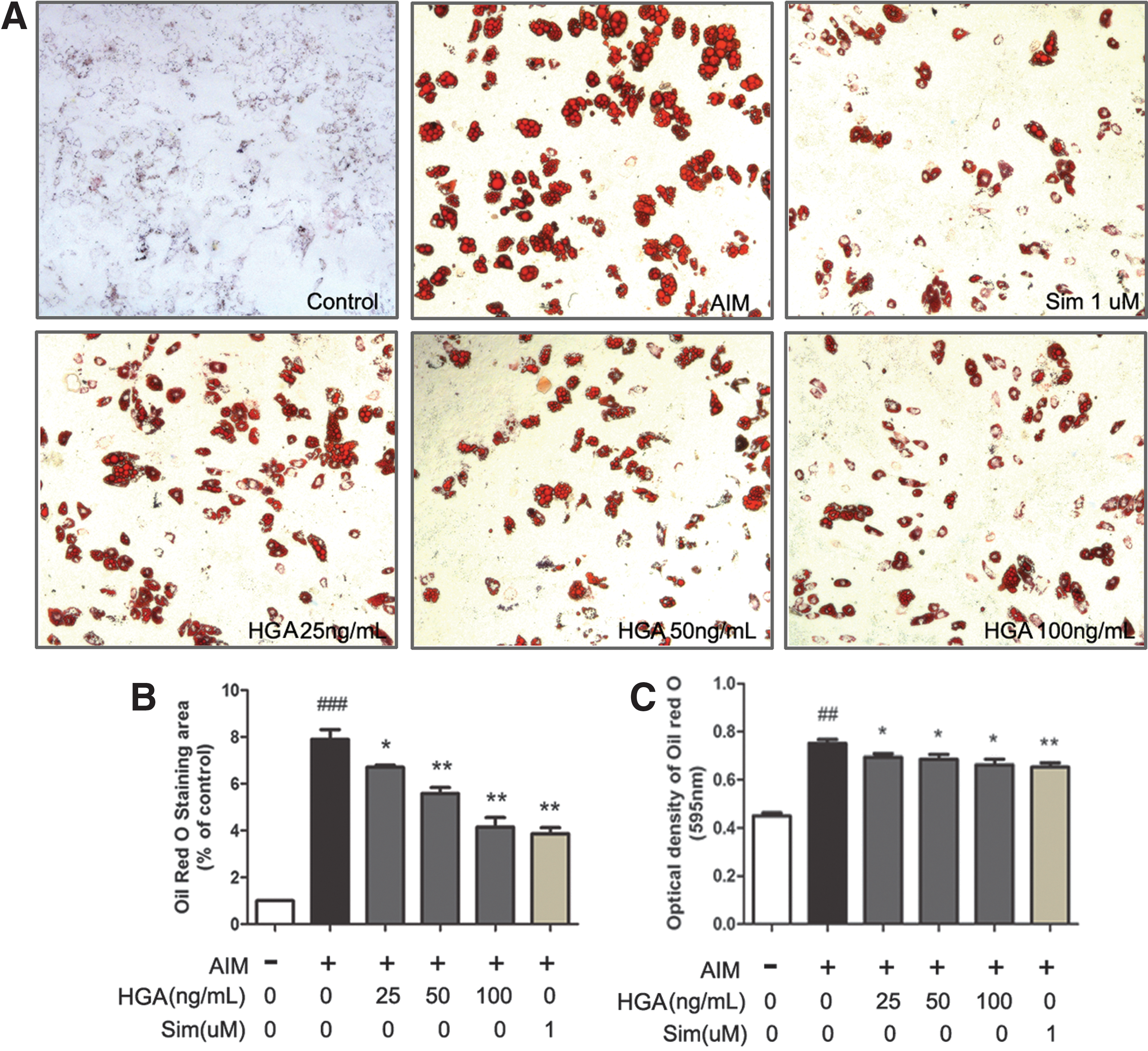
HGA inhibited adipogenic differentiation of MSCs.
Discussion
Our previous studies have found that HG is able to prevent the development of nontraumatic osteoporosis through different ways, such as regulation of lipid metabolism, increase of bone formation-related factors, and enhancement of the angiogenesis. The results from the current study showed that HGA, the aqueous fraction of HG extract, could inhibit adipogenic differentiation of rat MSCs. Most importantly, we found that HGA could promote osteogenesis of MSCs through activating both BMP2 and Wnt signaling pathways.
Bone marrow-derived MSCs are multi-potent progenitors that can commit to osteoblast, chondrocyte, adipocyte, and several other lineages. Accumulating evidence suggested that the imbalance between osteogenesis and adipogenesis of MSCs differentiation is a key factor of ONFH. 6,9 There is an inverse relationship between osteogenesis and adipogenesis; typically, increased osteogenesis is associated with decreased adipogenesis. 23 –25 Furthermore, quite a number studies have manifested that transplanted autologous or allogeneic MSCs were capable of survival, proliferation, and differentiation into osteoblasts directly in the necrotic femoral head on humans 26 –28 and a variety of animal models; for example, in mouse, 29 rabbit, 30,31 dog, 32 and sheep. 33 Thus, regulation on the osteogenesis and adipogenesis of MSCs is the novel therapeutics for ONFH treatment. Therefore, we explored the molecular mechanism by which HG impacts the function of ONFH during MSC differentiation.
ALP is considered an early marker of osteogenic differentiation, and induced mineralization during the differentiation process. 34,35 To evaluate the effect of HGA on osteoblastic differentiation of MSCs, ALP staining and Alizarin Red S staining were first performed. The data from the present study indicated that HGA significantly increased the activity of ALP in MSCs. As expected, HGA also enhanced calcium nodule formation, a functional marker of mineralization confirmed by Alizarin Red S staining. All these results showed that HGA could enhance the osteogenic differentiation of MSCs.
During the differentiation process, osteogenesis-related markers were expressed time dependently, which have been extensively documented in previous studies. Such markers as Runx2 and ALP (early/middle), Osx and OCN (late) 36 –40 were observed in our experiment. Runx2, regarded as the master regulator of osteogenesis, was expressed in the early stages of osteoblastic differentiation. Runx2 also serves at the intersection of many signaling pathways regulating osteoblast differentiation. ALP plays an important role in bone mineralization for it degrades pyrophosphate to generate phosphate, which reacts with calcium to form hydroxyapatite. 41 OCN is regarded as the most specific and latest described marker of osteogenic differentiation. 42 Osx acts downstream of Runx2, and Osx in combination with Runx2 seems to serve as a candidate pair to enhance the osteogenic potential of MSCs. Our results indicated that HGA significantly increased the mRNA of Runx2, Alp, Ocn, and Osx. These results further confirmed that HGA enhanced osteogenic differentiation of MSCs in both early and late stages.
Expression and activity of transcription factors such as Runx2 are regulated by multiple signaling pathways. Previous studies reported that BMP, Wnt, and Notch signaling pathways all play important roles in MSCs differentiation. 43 –45 Among all the BMP ligands identified now, BMP2, the most commonly studied BMP ligand, induces MSC osteogenesis both in vitro and in vivo. 46,47 BMP signaling is usually initiated by the binding of ligands, which phosphorylate BMP-responsive Smads 1/5/8 and then subsequently regulate osteogenic transcription. 48 In addition, BMP2 treatment is both necessary and sufficient for osteogenic commitment in the C3H10T1/2 cell line. 49 Furthermore, recombinant BMP2 has been approved by the FDA for the regeneration of specific regenerative indications. 50 To explore whether the BMP signaling pathway was also involved in the HGA-mediated osteogenic induction in MSCs, we detected the mRNA expression and protein levels secreted into the media after being induced by OIM. The results from this study showed that HGA significantly increased both the mRNA and protein levels of BMP2 in a dose-dependent manner. To further confirm our finding, we also investigated whether blocking the BMP signaling pathway with its inhibitor could impair the effect of HGA on osteogenesis. Our results showed that calcium nodule formation was highly suppressed, and osteogenesis-related markers were significantly decreased when pretreated with noggin. These data clearly indicated an involvement of the BMP signal pathway in the HGA-enhanced osteogenic differentiation of MSCs.
A second major signaling pathway involved in osteogenesis is Wnt signaling. This pathway leads to intracellular accumulation of β-catenin, a signaling protein that can then enter the nucleus to activate transcription of target genes. 44 Our results demonstrated that HGA significantly increased the β-catenin expression, secretion, and nuclear translocation in a dose-dependent manner. Furthermore, addition of Wnt inhibitors Dkk-1 could efficiently inhibit the effect of HGA on calcium nodule formation, expression of Runx2 and β-catenin in MSCs. These data suggested that the Wnt/β-catenin pathway was involved in the HGA-mediated osteogenic differentiation of MSCs. Given the fact that both Wnt and BMP signaling pathways are extremely critical for osteoblast differentiation and bone formation, HGA is a potential candidate for stimulating bone formation via the BMP and Wnt signaling pathways.
The process of bone formation involves four steps: migration of MSCs to site, condensation, differentiation, and mineralization of the extracellular matrix. We also investigate the effect of HGA on MSCs migration. Our results showed that HGA could promote the MSCs migration and proliferation. Between osteogenic and adipogenic lineage commitment and differentiation, a theoretical inverse relationship exists, such that differentiation toward an osteoblast phenotype occurs at the expense of an adipocytic phenotype. So, we checked how HGA impacts the adipogenic differentiation of MSCs. Data from our study indicated that HGA showed the tendency to inhibit adipogenesis, though without an obvious dose-dependent manner.
Potential limitations of this study include that there are distinct differences in the biological properties between rats and human. In addition, our results indicated the involvement of BMP and Wnt signaling pathways in the effect of HGA on MSCs differentiation, the regulation of Wnt and BMP pathways by HGA at which level and what is the key molecular target remain unclear. Furthermore, the effect of HGA on MSCs differentiation in vivo needs to be investigated. These limitations provide an agenda for further research in this area.
Conclusions
In conclusion, the present data demonstrated that HGA promoted osteogenesis of MSCs via the BMP and Wnts signaling pathways. Despite limitations identified, our findings provide mechanistic insights into the role of HG in treating ONFH.
Footnotes
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China (Nos. 81173417 and 81373656).
Authors' Contributions
X.Y.K. participated in the design of the study, performed the statistical analysis, and drafted the article. X.M.L. carried out the key experiments, performed the statistical analysis, and prepared figures and tables. H.Y.W., J.Z., C.L.L., and H.C.S. performed cell culture experiments and staining. L.L.Z. participated in the real time-polymerase chain reaction (RT-PCR). C.Z. prepared the plant material and chemical analysis. W.H.C. and N.L. conceived of the study, participated in its design and coordination, and helped draft the article. All authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.