
Abstract
Buyang Huanwu decoction (BYHWD), as a popular traditional Chinese medicine formula, was widely used for treating ischemic diseases. However, in the area of neurodegenerative diseases, the researches focused on BYHWD are rare but promising, and molecular mechanisms underlying are largely elusive. 6-Hydroxydopamine (6-OHDA), a dopaminergic-specific neurotoxin, is extensively used to establish neurotoxic model in vivo and in vitro. In our present study, we prepared drug-containing serum of BYHWD (Buyang Huanwu drug-containing serum [BYHWS]) based on serum pharmacology methodology. Neurotoxic model in vitro was established in PC12 cells, and innovative experimental grouping method was adopted to investigate neuroprotective effects of BYHWS on neurotoxicity induced by 6-OHDA exposure. Remarkably, BYHWS vigorously rescued PC12 cells from 6-OHDA-induced neurotoxicity even to surpass 100% in cell viability. Moreover, Hoechst/propidium iodide (PI) staining revealed that cell apoptotic rate was reduced significantly after incubation of BYHWS. Besides, BYHWS effectively restored the disruption of mitochondrial membrane potential and attenuated the elevation of intracellular reactive oxygen species level caused by 6-OHDA insult. Furthermore, BYHWS remarkably reversed the dephosphorylation of Akt (protein kinase B) and glycogen synthase kinase-3β (GSK3β) evoked by 6-OHDA. The above protective effects were attenuated by coculturing with Akt inhibitor LY294002. In summary, we concluded that the BYHWS vigorously blocked 6-OHDA-induced neurotoxicity via Akt/GSK3β pathway and provided a novel insight into roles of BYHWD in the clinical practices on neurodegenerative diseases.
Introduction
6-
BYHWD, one of the most popular TCM formulae, was widely prescribed for treatment of some ischemic cardiocerebral vascular diseases and promoting peripheral nerve regeneration. 13 –15 Besides, previous studies have demonstrated that BYHWD had various pharmacological effects, such as antioxidative, growth promoting, anti-inflammatory, and antiapoptosis. 16 –18 Among all the above effects, antioxidative, which aims at the crucial step of 6-OHDA insult, is the prominent characteristic of BYHWD. 18 So it is pretty rational to make a hypothesis that BYHWD has neuroprotective effects on 6-OHDA-induced neurotoxicity. Earlier evidences have indicated that the BYHWD has shown comprehensive effects on growth promotion. 15,17,19 However, effects of BYHWD on vascular smooth muscle cells are exactly the opposite. 20,21 BYHWD has been studied in certain neurotoxic models, such as H2O2 damage model and hypoxic model, 18,22 in which BYHWD evinced irregular neuroprotective effects to a certain extent but not remarkably. Most studies of BYHWD referred to the acute status, such as ischemic cardiovascular disease, stroke, and hypoxia. 14 However, in the field of chronic diseases, such as neurodegenerative diseases, researches focused on BYHWD are relatively rare but promising. The mechanisms involved in the protective effects of BYHWD in these acute diseases exactly aimed at the pathogenesis of chronic diseases. 16 Thus, some paradoxes and interesting phenomena remain to be elucidated in the studies of BYHWD.
After oral administration of the TCM, just parts of its ingredients are absorbed into the blood circulation and transformed into substances that might be different from the herb ingredients. Some drugs do not exert pharmacological effects until after an organism's metabolism. 23 Serum pharmacology is a scientific analysis method by which drug-containing serum is obtained from animals taking TCM orally. 24,25 This method not only considers the interactions between organism and TCM but also eliminates the biases from the complexity of TCM. In this study, we followed the principles of serum pharmacology and proposed our hypothesis.
Previous studies demonstrated that Akt/GSK3β pathway was involved in 6-OHDA-induced neurotoxicity. 26,27 Akt (protein kinase B) mediates various transduction pathways, such as metabolism, cell stress, and apoptosis 28 and phosphorylates diverse substrates, including mammalian target of rapamycin (mTOR) and glycogen synthase kinase-3β (GSK3β). Akt activation attenuates GSK3β activity by promoting its phosphorylation. 27 As a crucial downstream target of Akt, GSK3β was found to be implicated in the apoptosis induced by oxidative damage. 29,30 Besides, the activation of Akt/GSK3β stimulated proliferation in various tumor cells. 31,32 Moreover, it has been reported that BYHWD promoted angiogenesis via PI3K/Akt pathway. 33 Together, we speculated that Akt/GSK3β pathway may be involved in the neuroprotective effects of BYHWS.
In summary, we established neurotoxic model in vitro through 6-OHDA-induced neurotoxicity in PC12 cells. By virtue of the principles of serum pharmacology, we prepared BYHWS. The objective of our present study was to demonstrate our assumption that BYHWD had neuroprotective effects against the 6-OHDA-induced neurotoxicity via Akt/GSK3β pathway based on serum pharmacology methodology.
Materials and Methods
Ethics statement and animals
Adult male Wistar rats, weighing 250 ± 30 g, were purchased from the Laboratory Animal Center of the Shandong University. All the animals were reared under conditions of temperature control with water and food ad libitum. All procedures were approved by the Institutional Animal Care and Use Committee of the Shandong University and performed in accordance with the National Institutes of Health Guide for the Care and Use of Laboratory Animals.
Reagents and drugs
BYHWD was composed of Radix Astragali Membranaceus (60 g), Radix Angelicae Sinensis (10 g), Radix Paeoniae Rubra (10 g), Rhizoma Ligustici Chuanxiong (10 g), Semen Persicae (10 g), Flos Carthami (10 g), and Pheretima (10 g) 34 and purchased from ShenNong Bencao Pharmacy (Shandong, China). Dulbecco's modified Eagle's medium (DMEM) and fetal bovine serum (FBS) were purchased from HyClone. MTT was purchased from Solarbio (Beijing, China). Mitochondrial Membrane Potential (MMP) Assay Kit, Reactive Oxygen Species (ROS) Assay Kit, Hoechst 33342, and propidium iodide (PI) were obtained from Beyotime Biotechnology (Shanghai, China). Anti-GSK3β, anti-Akt, anti-pGSK3β (Ser 9), and anti-pAkt were purchased from Cell Signaling Technology (Danvers, MA).
Cell culture
Rat differentiated PC12 cells (pheochromocytoma) were provided by the Shanghai Cell Bank of Chinese Academy of Sciences and cultured in DMEM supplemented with 10% FBS in an incubator with an atmosphere of 95% humidity and 5% CO2 at 37°C.
Preparation of drug-containing serum
Forty Wistar rats under similar conditions were divided into BYHWS and control serum (CS) groups randomly. The rats of BYHWS and CS groups were treated with BYHWD and normal saline, respectively. The administered dose per time was 6 mL/100 g at decoction concentrations of 1 g/mL. The rats were intragastric administrated at 10
Experimental groups
PC12 cells were divided into the following three groups if no special instructions: (1) 6-OHDA damage group, cells were exposed to 100 μM 6-OHDA in the presence of 10% CS for 24 hours after preculturing with 10% CS for 1 hour; (2) BYHWS protection group, cells were exposed to 100 μM 6-OHDA in the presence of 8% BYHWS plus 2% CS for 24 hours after preculturing with 8% BYHWS plus 2% CS for 1 hour; and (3) normal control group, cells were cultured in DMEM containing 10% CS without 6-OHDA for 25 hours (2%, 4%, 8%, and 10% represented the final concentrations of serums in culture medium).
MTT assay
The MTT assay was used to detect the cell viability of PC12 cells. PC12 cells were plated in 96-well plates at a density of 1 × 105/mL and intervened with different concentrations of 6-OHDA and serum for 24 hours. We added MTT (0.5 mg/mL) into each well, and then, the cells were incubated for 4 hours (at 37°C and 5% CO2). Subsequently, culture supernatant was removed, and formazan was dissolved in 100 μL dimethyl sulfoxide (DMSO). The absorption value was assessed at 570 nm using an enzyme-linked immunosorbent assay (ELISA) reader (Thorm Multiskan Mk3, Thermo Labsystems, USA).
Hoechst 33342 and PI double staining
Hoechst 33342/PI staining was performed to examine apoptotic rate of PC12 cells. PC12 cells were pretreated with different serums for 1 hour and then cocultured with 100 μM 6-OHDA for 24 hours. Procedures were carried out according to the previous report. 35 In brief, PC12 cells were incubated with 10 μg/mL Hoechst 33342 and PI at 37°C for 15 minutes and then washed with phosphate-buffered saline three times. Morphological changes in PC12 cells were detected by a fluorescence microscope (Carl Zeiss SAS, Jena, Germany). Hoechst 33342 and PI can interact with nucleic acid. Hoechst 33342, which can cross the intact cell membrane, integrates with A–T bases of DNA and manifests blue fluorescence, whereas PI, which can only cross the damaged cell membrane, manifests red fluorescence. The apoptotic rate was regarded as the percentage of apoptotic nuclei in five randomly fields.
Measurement of MMP
The alterations of MMP were estimated with the fluorescent probe JC-1. The cells were plated in 24 wells at a concentration of 1 × 105/mL. After different interventions, PC12 cells were incubated with 10 μM JC-1 working solution at 37°C for 20 minutes and then rinsed twice with JC-1 washing buffer. The cells were observed under a fluorescence microscope. The fluorescence intensity was analyzed using Image-Pro Plus.
ROS assays
DCFH-DA, as a fluorescent probe, was used to investigate the level of intracellular ROS. The PC12 cells were plated in 24 wells at a concentration of 5 × 104/well. Briefly, the cells were cocultured with 250 μL DCFH-DA (1:1000 mixed with culture medium) at 37°C for 20 minutes after the drug interventions. Subsequently, the cells were washed with DMEM three times and 250 μL DMEM was added into every well and then observed in microscopy. All images were captured under fluorescence microscopy and analyzed by ImageJ.
Western blotting
PC12 cells were lysed by radioimmunoprecipitation assay (RIPA) buffer, and total proteins were extracted after interventions. The concentrations of protein samples were measured by BCA Protein Assay Kit. Protein samples were separated by 10% sodium dodecyl sulfate–polyacrylamide gel electrophoresis and then electrotransferred to nitrocellulose membrane. Protein bands of interest were collected and blocked by blocking buffer containing 5% skim milk for 2 hours at room temperature. Incubation was performed with the primary antibodies (1:1000) against GAPDH, pAkt, pGSK3β (Ser 9), Akt, and GSK3β at 4°C overnight and subsequently with the secondary antibodies (1:5000) for 1 hour at room temperature. The bands were tested using the western immunoblotting detection reagents and semiquantitatively analyzed by ImageJ.
Statistical analysis
All data are expressed as mean ± standard error of the mean. Statistical analysis was performed with one- or two-way (not repeated measures) analysis of variance, followed by Tukey's post hoc analysis. p < 0.05 was considered as statistically significant. GraphPad Prism software was used in all statistical analysis.
Results
BYHWS vigorously rescues PC12 cells against 6-OHDA-induced neurotoxicity
Neurotoxic model in vitro was constructed by treating PC12 cells with a series of concentrations of 6-OHDA. Five concentrations (50, 75, 100, 125, and 150 μM) were selected to determine the optimum damage concentration. Morphological alterations were obvious that cells grew into bright round shape with shorter neurites and less number in the 6-OHDA interventional groups. There was a distinguished dose-dependent neurotoxicity according to the results of the MTT assay (Fig. 1H). Compared to the normal control group, PC12 cells exposed to 100 μM 6-OHDA had a cell viability of 0.54 ± 0.04 (p < 0.001). Therefore, 100 μM was determined as the optimum concentration in the following experiments.
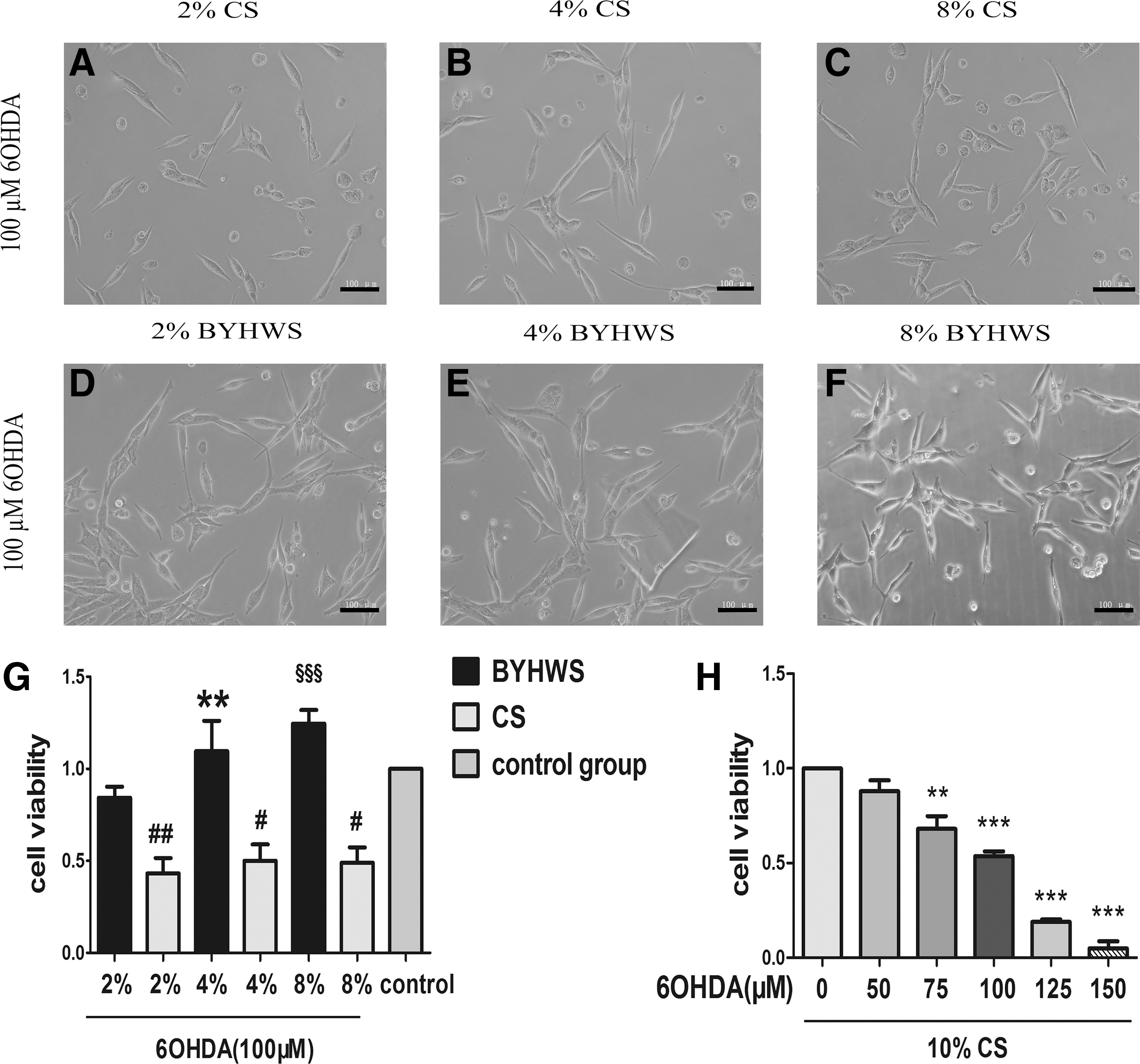
BYHWS vigorously rescues PC12 cells against 6-OHDA-induced neurotoxicity. PC12 cells were cultured in different concentrations of 6-OHDA (0, 50, 75, 100, 125, and 150 μM) and 10% CS for 24 hours.
To provide more persuasive evidences for our study, we elaborated two kinds of experimental grouping method. As shown in Figure 1A–G, morphological injuries were almost reversed after the incubation of BYHWS. Cells in the BYHWS group had more prominent neurites, which showed a tendency to form a network and less bright round vacuolar cells. Consistently, the MTT results indicated that BYHWS vigorously ameliorated cell viability in a dose-dependent manner. In the presence of 4% and 8% BYHWS, cell viabilities (1.10 ± 0.17 and 1.25 ± 0.07) even exceeded the control group, which was designed as a standard.
Considering the limitation of the above method, which could not perform consecutive multiple comparisons due to different concentrations of serum, we described another method in which the total concentration of serum, including BYHWS and CS, added up to 10% in every group. As shown in Figure 2A–D and I, under the condition of 100 μM 6-OHDA, cell viabilities of BYHWS protection groups were significantly higher than that of 6-OHDA damage group, and the morphological alterations of corresponding groups confirmed the MTT results. In the presence of 4% and 8% BYHWS, cell viabilities exceeded the normal control group, which was consistent with method 1. Surprised by this phenomenon, we increased the concentration of 6-OHDA to 150 μM. As shown in Figure 2I, BYHWS protection groups produced a statistically significant improvement in cell viability compared to 6-OHDA damage groups when intervened with 150 μM 6-OHDA, which was the lethal concentration. With the increase in concentration of BYHWS, cell viability increased steadily in a dose-dependent manner. Morphological alterations (Fig. 2E–H) verified the MTT results.

BYHWS shows significant neuroprotective effects against 6-OHDA under the condition of total 10% mixed serum (BYHWS plus CS). PC12 cells were preincubated with different concentrations of BYHWS and CS for 1 hour. The total concentration of serum, including BYHWS and CS, added up to 10%. Then, PC12 cells were cultured for 24 hours in the presence of 100 μM
Otherwise, the growth-promoting effects of BYHWS were easily observed. Results demonstrated that BYHWS groups had strikingly higher cell viabilities than those of CS groups under three similar serum concentrations without 6-OHDA (Fig. 2J, p < 0.001). However, the dose-dependent manner of growth promotion in 2%, 4%, and 8% serum groups was inconspicuous (p > 0.05).
Investigate the neuroprotective pattern of BYHWS: extracellular neutralization or cellular protection?
Interesting in this vigorous neuroprotective effects of BYHWS, we considered whether the neuroprotective pattern was extracellular neutralization or cellular protection. Given this suspect, we designed an experiment to explore the neuroprotective pattern of BYHWS. If the neuroprotective pattern of BYHWS was extracellular neutralization, the effects of 6-OHDA would be terminated immediately after adding BYHWS, and there would be no difference in cell viability between the 6-OHDA-removed groups and 6-OHDA-maintained groups. According to the results (Fig. 3C), the optical density (OD) values of 6-OHDA-removed groups (b, e, and h) were obviously higher than those of 6-OHDA-maintained groups (a, d, and g), respectively. They were statistically significant between a and b (p < 0.001) and g and h (p < 0.05). The OD values of groups without 6-OHDA treatment (c, f, and i) were considered as control, which aimed at excluding the biases from complicated processes of a series of interventions. From a holistic perspective, OD values of groups with different secondary intervention times decreased in a time-dependent manner (Fig. 3B, p < 0.001).
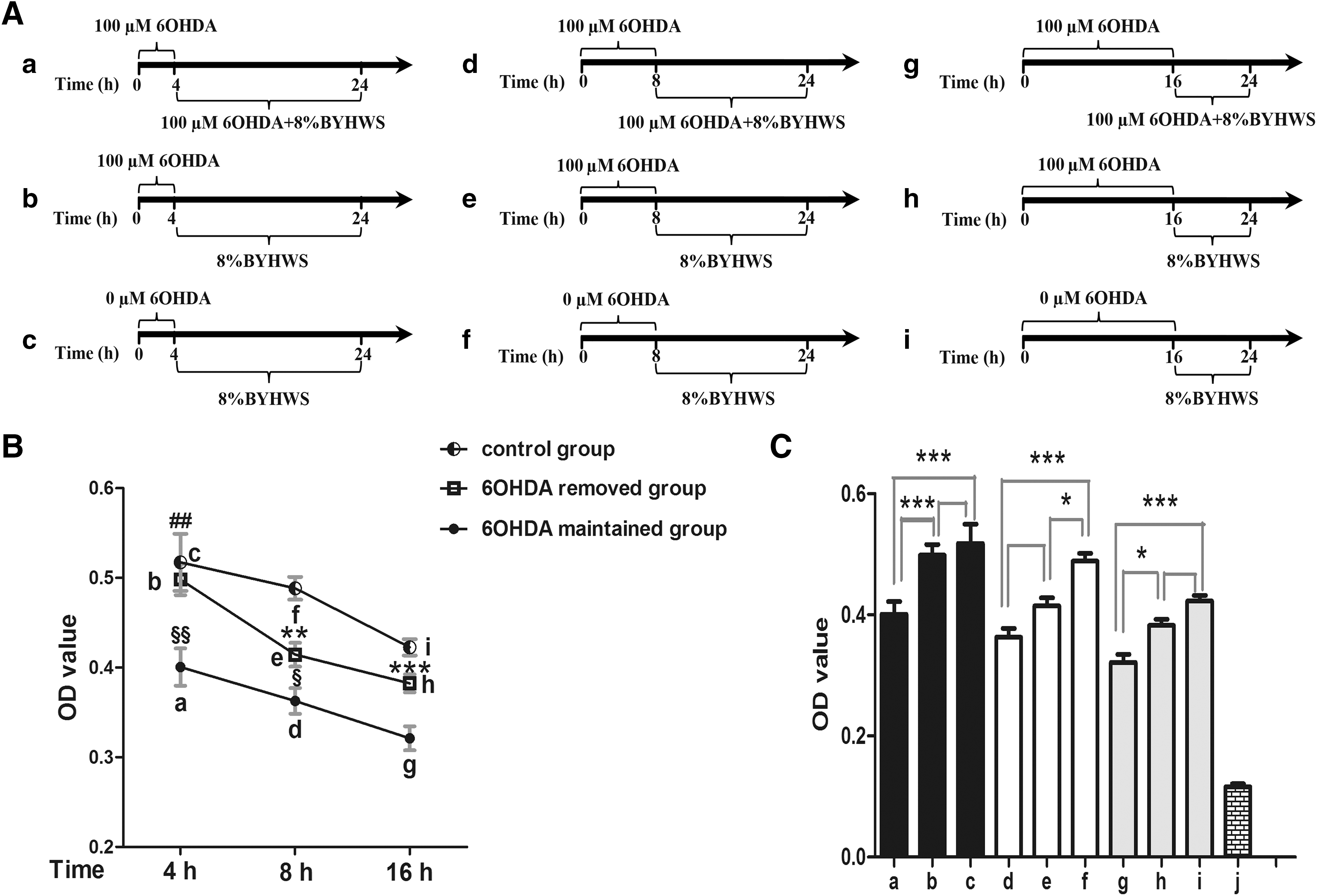
BYHWS exerts cellular neuroprotection against 6-OHDA damage. We divided the cells into three blocks (black, white, and gray), which were further subdivided into nine groups. In group
BYHWS decreases PC12 cell apoptosis against 6-OHDA-induced neurotoxicity
Chromatin condensation and nuclear shrinkage, which are represented in the form of bright blue fluorescence in the Hoechst 33342/PI staining, are remarkable morphological changes of cell apoptosis. Red fluorescence indicates dead cells as the result of penetration of PI, whereas the viable cells are merely stained by Hoechst 33342 and thus show dull blue fluorescence. As the microphotographs and the histogram illustrated, more apoptotic cells with bright blue nuclei were detected in the 6-OHDA damage group (Fig. 4A, B, and D). However, in the presence of BYHWS, the count of apoptotic nuclei decreased obviously, and the apoptotic rate of PC12 cells remarkably dropped from 48.7% ± 5.2% to 6.2% ± 1.2% (p < 0.001).
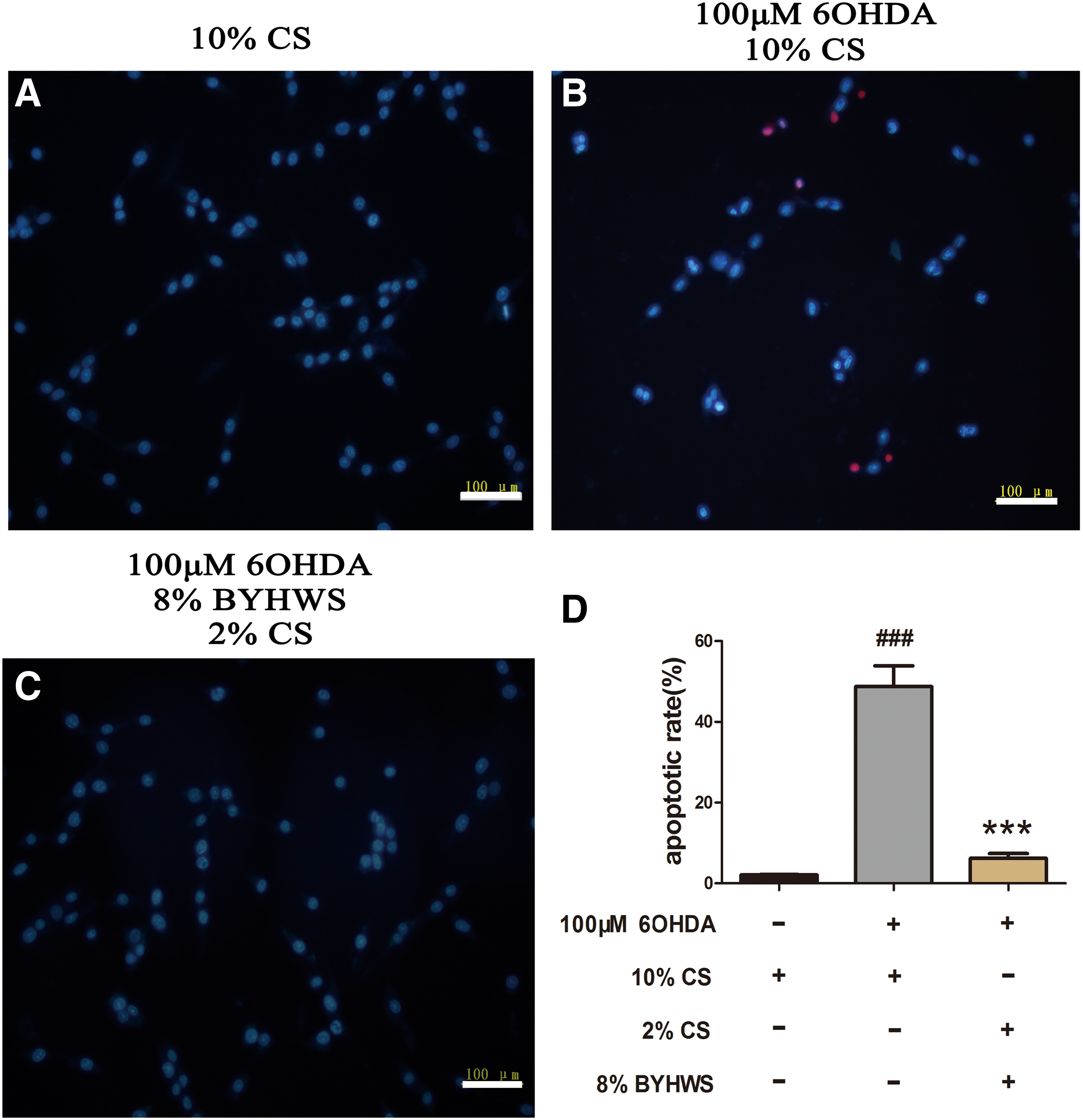
BYHWS decreases the apoptosis of PC12 cells. PC12 cells were pretreated with 10% mixed serum for 1 hour and then cocultured with 100 μM 6-OHDA for 24 hours. Bright blue fluorescence indicates apoptotic cells, whereas the red fluorescence indicates necrotic cells.
BYHWS obviously elevates MMP and decreases the intracellular ROS level
The alteration of MMP (ΔΨm) is an early event of apoptosis. Existence form of JC-1 alternates in a potential-dependent manner. JC-1 aggregates form under high MMP, which are represented as yellow fluorescence and depolymerize under low MMP, which are represented as green fluorescence. As shown in Figure 5A and C, 6-OHDA significantly decreased the level of MMP, while preintervention with BYHWS significantly reversed the disruption of MMP. Notably, BYHWS decreased fluorescence intensity from 89.0% ± 4.3% to 8.3% ± 2.4% (p < 0.001) compared to the 6-OHDA damage group. ROS is a critical part of apoptotic pathway of PC12 cells caused by 6-OHDA exposure. The involvement of oxidative stress was investigated by virtue of measuring intracellular ROS level by the DCFH-DA probe. Figure 5B and D illustrated that intracellular ROS level increased after 6-OHDA insult; however, BYHWS effectively attenuated the intracellular level of ROS compared to the 6-OHDA damage group (p < 0.001).

BYHWS rescues MMP (ΔΨm) and downregulates the intracellular ROS level. PC12 cells were pretreated with 10% mixed serum for 1 hour and then incubated with 100 μM 6-OHDA for 24 hours.
BYHWS inhibits the dephosphorylation of Akt/GSK3β pathway
Western blotting was exhibited to explore the Akt/GSK3β expression in PC12 cells exposed to 6-OHDA combined with BYHWS. As shown in Figure 6, BYHWS obviously inhibited the dephosphorylation of Akt/GSK3β and reversed the decrease in the ratio of pAkt/Akt (p < 0.01) and pGSK3β/GSK3β (p < 0.05) caused by 6-OHDA insult. However, there was no statistical difference in the expression of total proteins of Akt and GSK3β under the condition of serum interventions (p > 0.05).
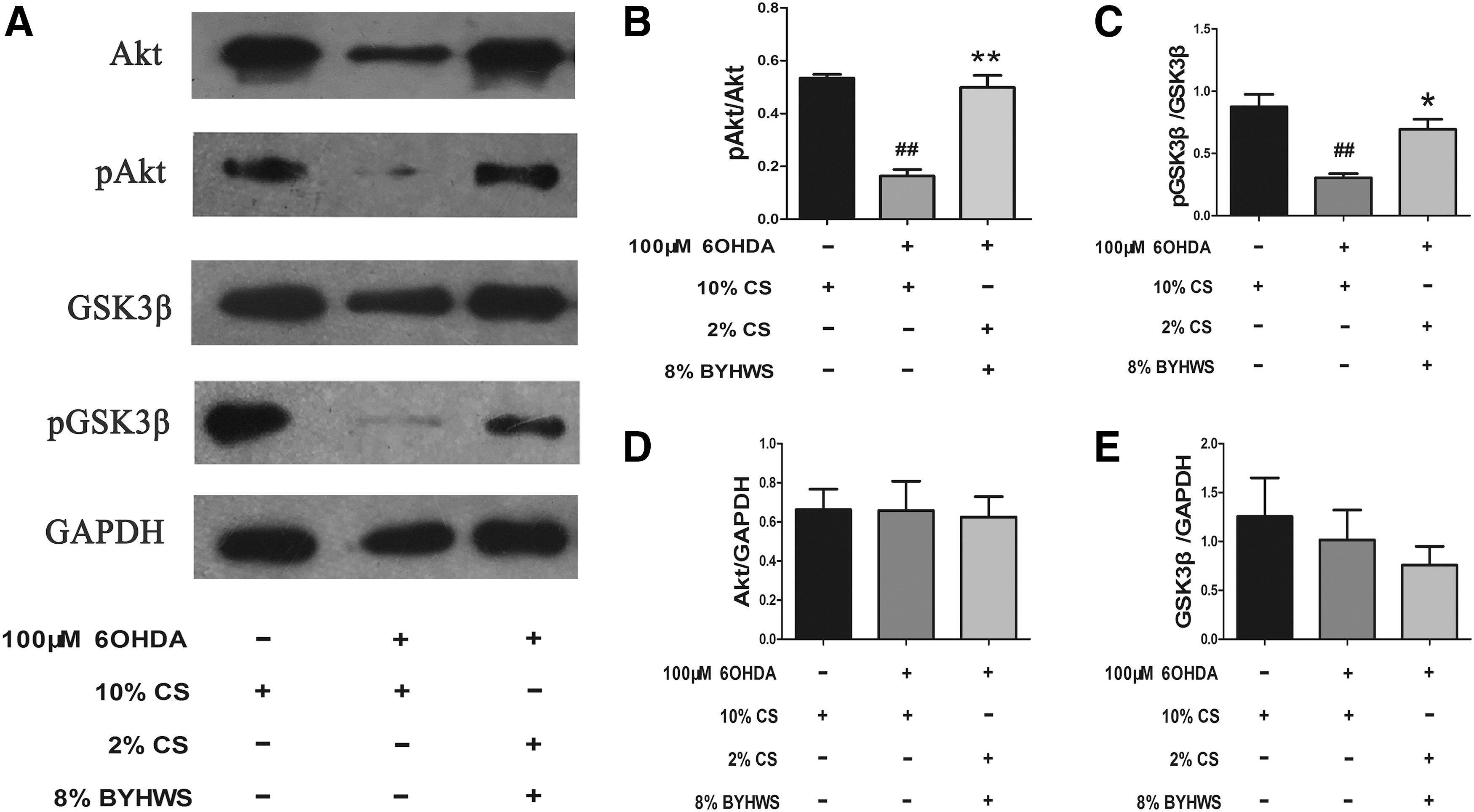
BYHWS suppresses the dephosphorylation of Akt/GSK3β pathway. After treating with 100 μM 6-OHDA plus 10% mixed serum for 24 hours, the proteins were extracted and prepared for western blotting. GAPDH was selected as loading control.
Akt inhibitor LY294002 partly attenuates neuroprotective effects of BYHWS
To further confirm the involvement of Akt/GSK3β pathway in neuroprotective effects of BYHWS, LY294002, a specific Akt inhibitor, was coincubated with BYHWS and 6-OHDA. The MTT assay and Hoechst 33342/PI staining were performed simultaneously after drug interventions. Results indicated that the viabilities of PC12 cells were obviously reduced in the BYHWS protection group (Fig. 7, p < 0.05 vs. BYHWS protection group) when LY294002 was applied. Moreover, cell apoptotic rate was attenuated, at least partly, in the BYHWS protection group (p < 0.05) after LY294002 exposure.

LY294002 partly attenuates neuroprotective effects of BYHWS. PC12 cells exposed to 100 μM 6-OHDA were cocultured with 10% mixed serum in the presence of 10 μM LY294002 for 24 hours.
Discussion
In the present study, neurotoxic model in vitro was established to investigate the neuroprotective effects of BYHWS on damages caused by 6-OHDA insult. Much to our surprise, BYHWS almost completely rescued damaged cells against 6-OHDA neurotoxicity. Serum pharmacology methodology was the cornerstone to perform quality control. 24 Innovative experimental grouping method was adopted to eliminate biases of different serum concentrations. Furthermore, we explored the underlying molecular mechanism and found that ROS-mediated Akt/GSK3β pathway was involved in the process of BYHWS neuroprotection.
Traditional grouping method could not perform consecutive multiple comparisons. Considering its limitation, we elaborated a novel grouping method that considered the proportion of BYHWS as a variable under the condition of 10% total serum concentration. This grouping method eliminated biases that stem from different serum concentrations and was adopted in the following experiments. Survival rate is the most direct indicator of cell status. In our study, the MTT assay was performed to investigate the survival rate and growth of PC12 cells. Surprisingly, BYHWS vigorously rescued cells against neurotoxicity caused by 6-OHDA, which strongly elevated cell viability from 0.49 to 1.25 (Fig. 2I). Then, we improved the grouping method and increased the concentration of 6-OHDA to 150 μM, which was a lethal concentration. It turned out that BYHWS possessed vigorous neuroprotective effects, which show an obvious concentration-dependent manner even under the condition of 150 μM 6-OHDA.
Interesting in this unexpected consequence, we speculated that neuroprotective pattern of BYHWS might be extracellular neutralization and elaborated another experiment to explore this phenomenon. As explained before in the “Results” section, our experiments found that cell viabilities of 6-OHDA-maintained groups were lower than those of 6-OHDA-removed groups, and they were statistically significant, and there was a remarkable time-dependent decrease in cell viability in groups with different secondary intervention times. Moreover, considering the fact that the BYHWS protection group exceeded the normal control group in cell viability, we inferred that a growth-promoting effect may have existed and went a step further to investigate BYHWS effects on cell growth. The results corresponded with our speculation. BYHWS incubation strikingly stimulated cell growth. However, growth-promoting effect cannot explain the vigorous anti-6-OHDA effects under a lethal concentration of 150 μM, and morphological alterations in all assays demonstrated the existence of neuroprotective effects. In summary, three aspects of evidences were recapitulated to certify cellular protection pattern of BYHWS: concentration-dependent protective effects of BYHWS under different concentrations of 6-OHDA as the indirect evidence, specific experiment as the direct evidence, and growth-promoting effect partly involved in the rescue effect of BYHWS.
Growing empirical evidences indicated that oxidative stress may be a key step in the mechanism of action of 6-OHDA neurotoxicity 6,36 and triggered a series of waterfall cascades, such as calcium overload, mitochondrial dysfunction, and biomacromolecule degradation. 37 Mitochondrial dysfunction precedes the morphological alteration of early apoptosis. ROS disrupts MMP, which in turn elevates the level of intracellular ROS level. Vicious circle forms to initiate apoptotic pathway. 37,38 Our results showed that apoptotic rate decreased significantly in the BYHWS protection group. Accordingly, BYHWS attenuated ROS fluorescence and strongly elevated MMP. Our investigation conclusively indicated that ROS-mediated apoptotic pathway was involved in the rescue effect of BYHWS.
Akt/GSK3β pathway played an important part in cellular apoptosis and pathogenesis of schizophrenia. 28,39 It has been reported that Akt/GSK3β pathway was involved in the embryonic brain development. 40 Besides, the activation of Akt/GSK3β pathway downregulated dopamine transporter in the nucleus accumbens and attenuated behavior sensitization. 41 Moreover, previous studies have indicated that dysregulation of Akt signaling pathway not only involved in the PD pathology 42 but also became the novel target of PD treatment. 43 Interestingly, neurotrophic factors (including brain-derived neurotrophic factor [BDNF] and glial cell line-derived neurotrophic factor [GDNF]) and Akt had a relationship of mutual promotion. For one thing, GDNF was upregulated by Akt activation, and for another thing, Akt mediated the neuroprotective effects of BDNF and GDNF. 44,45 Our results demonstrated that BYHWS inhibited the downregulation of pAkt and pGSK3β (Ser 9) induced by 6-OHDA exposure, which was in line with previous studies showing 6-OHDA dephosphorylation of Akt/GSK3β pathway. However, we did not investigate another phosphorylation site of GSK3β (Tyr216) because it did not show upstream and downstream relationships with Akt. 46 Remarkably, the molecular changes in the BYHWS protection group were so drastic that the phosphorylation of Akt/GSK3β almost reached to the normal control group. The results matched to the rescue effect and vigorous morphological alterations. Therefore, we proposed that BYHWS exerted its neuroprotective effects via Akt/GSK3β pathway. Furthermore, the neuroprotective effects were significantly attenuated by coincubation with Akt inhibitor, LY294002, which substantiated our original inference.
However, present studies focus on TCM limit to the compound medicine without going further to the level of their constituents. They were all inadequate to the systematic comprehensive study of TCM. In the formula of BYHWD, which possesses seven individual herbs, there must be certain explicit components responsible for the vigorous neuroprotective effects, and interactions between different herbs may be synergy or reinforcement to its effects, which is beyond the range of our study. Thus, further researches remain to be unfolded.
In summary, we investigated the neuroprotective effects of BYHWS on 6-OHDA-induced neurotoxicity in PC12 cells and validated the involvement of ROS-mediated Akt/GSK3β pathway. Our study highlighted the striking rescue effect of BYHWS, suggesting BYHWD might be the potential treatment for neurodegenerative diseases and providing preliminarily data for further animal and clinical experiments.
Footnotes
Acknowledgments
This study was supported by grants from the National Natural Science Foundation of China (81371471) and the Shandong Provincial Natural Science Foundation (ZR2012HQ018) and by the research project for the Doctoral Program of Ministry of Education of China (20120131110039).
Author Disclosure Statement
No competing financial interests exist.