
Abstract
Normal arterial aging processes involve vascular cell dysfunction associated with wall stiffening, the latter being due to progressive elastin and elastic fiber degradation, and elastin and collagen cross-linking by advanced glycation end products (AGEs). These processes progressively lead to cardiovascular dysfunction during aging. Elastin is only synthesized during late gestation and childhood, and further degradation occurring throughout adulthood cannot be physiologically compensated by replacement of altered material. However, the ATP-dependent K+ channel opener minoxidil has been shown to stimulate elastin expression in vitro and in vivo in the aorta of young adult rats. Therefore, we have studied the effect of a 10-week chronic oral treatment with minoxidil (120 mg/L in drinking water) on the aortic structure and function in aged 24-month-old mice. Minoxidil treatment increased tropoelastin, fibulin-5, and lysyl-oxidase messenger RNA levels, reinduced a moderate expression of elastin, and lowered the levels of AGE-related molecules. This was accompanied by the formation of newly synthesized elastic fibers, which had diverse orientations in the wall. A decrease in the glycation capacity of aortic elastin was also produced by minoxidil treatment. The ascending aorta also underwent a minoxidil-induced increase in diameter and decrease in wall thickness, which partly reversed the age-associated thickening and returned the wall thickness value and strain-stress relation closer to those of younger adult animals. In conclusion, our results suggest that minoxidil presents an interesting potential for arterial remodeling in an antiaging perspective, even when treating already aged animals.
Introduction
L
Elastin and collagens are the major ECM proteins that make up the framework of the elastic artery structure. These two fibrous proteins are key structural components of arterial walls, and they provide the strength and resilience needed by the aorta to accommodate the pressure and volume variations during each heartbeat. Elastin, constituting more than 50% of the dry weight of the proximal parts of the aorta, assembles with microfibrils into elastic fibers, which are responsible for the elastic properties of the large arteries. Microfibrils directly contribute to the mechanical properties of elastic fibers, 2 while collagen stiffens the vascular wall and limits its extensibility. 3 Other ECM or microfibrillar components must properly interact to allow for a correct assembly of elastic fibers, in particular the cross-linking enzymes lysyl-oxidase (LOX) and lysyl-oxidase-like-1 (LOXL-1), and fibrillin-1—the major constituent of microfibrils—and fibulin-5. In case of genetic deficiency or improper arrangement of some elastic fiber component(s), abnormal organization and function of the arterial wall occurs, such as in Williams or Marfan syndromes. 4 –6
Elastin is essentially synthesized during development and childhood. 7 During arterial development, elastin synthesized by vascular smooth muscle cells (VSMCs) forms elastic fibers that are arranged into concentric elastic lamellae around the lumen. Each elastic lamella alternates with a VSMC layer, which forms lamellar units allowing the artery to comply with the increased hemodynamic stress induced by systolic blood ejection and to maintain arterial blood pressure and flow during diastole. 8
During aging, the aorta diameter and stiffness increase because of degenerative changes in elastin, resulting in a transfer of stress to less extensible collagenous components of the aorta. Age-related large vessel stiffening is an important process underlying the development of isolated systolic hypertension in humans, 9 although not or very moderately in mice. 10,11 Conversely, epidemiological studies have shown that preexistent hypertension could induce large artery stiffening. This led to the concept of and debate about a bidirectional interaction between hypertension and arterial stiffness. 9 Although there is no consensus or complete understanding about causality, a strong link exists between the increased prevalence of systolic hypertension with age, 12 stiffness of conduit vessels, diastolic hypotension, heart remodeling, and cardiovascular disease morbidity and mortality, including aging-heart failure. 13 –16
Elastin production ends at the end of childhood, adults then live with an elastin stock. Elastin and elastic fibers are progressively degraded by enzymatic processes involving an age-related imbalance between anti-proteases and proteases. Elastin-degrading enzymes, that is, elastases, include several matrix metalloproteases (MMP), such as MMP-2 and MMP-9. 17 However, increases in the aortic stiffness due to processes other than enzymatic degradation of elastin also contribute to the appearance of age-associated cardiovascular dysfunction or diseases. These mechanisms mainly include the following: (1) accumulation of aortic elastic fiber ruptures mechanically induced by systolo-diastolic cyclic stretch throughout life; (2) age-dependent increase in the collagen content; (3) change in the intrinsic VSMC contribution to tissue mechanical properties, especially cell number and cytoskeletal—at least α-smooth muscle actin—remodeling 18 ; (4) shifts in the ratios of the different collagen types, in particular types I, III (promoting elasticity), and VIII 19 ; and (5) non-enzymatic glycation of elastin and collagens, leading to the formation of deleterious advanced glycation end products (AGEs) and related molecular cross-links that modify the tissue mechanical properties. 20,21
Since quality and possibly quantity of elastic fibers decrease during aging, 22,23 the identification of molecules capable of enhancing elastin synthesis in mature animals would be of major importance to counteract the age-associated arterial stiffening of large arteries. Minoxidil, an ATP-dependent potassium channel (KATP) opener initially developed as an antihypertensive molecule, 24 was shown to elevate tropoelastin—the monomeric elastin precursor—mRNA (messenger RNA) levels in cultured VSMCs 25 and elastin content in several arteries, including the aorta, in chronically treated rats, 26,27 through a calcium-dependent mechanism. 28 Further supporting the impact of potassium metabolism on elastin synthesis is the fact that a chronic oral treatment of young adult rats with another potassium channel opener, nicorandil, also increased the aortic elastin to total protein ratio. 29 However, no demonstration of pharmacologically induced increase in the arterial elastin or elastic fiber content has ever been done in aged animals. Here, we have chosen to investigate the effect of minoxidil on the elastin and AGE metabolisms and contents, in addition to structure and function of the aorta in aged (24-month-old) mice. Our results show for the first time that minoxidil triggers an increase in aortic tropoelastin mRNA, elastin, and elastic fiber contents, and a decrease in AGE levels, impacting the arterial function of aged animals.
Materials and Methods
Animals
All C57B1/6J male mice were purchased from Janvier (Le Genest-Saint-Isle, France). Thirty-two animals were studied at 24–25 months of age (aged mice). In addition, eight untreated animals aged 6 months (young adult mice) were used in the aorta mechanics and reactivity experiments. All housing and surgical procedures were in accordance with institutional guidelines.
Minoxidil treatment
Minoxidil treatment was given in tap water ad libitum at the concentration of 120 mg/L, as described.
26,27,30
A group of aged mice was treated for 10 weeks before the analyses were performed. The control mice were given water without minoxidil ad libitum. Minoxidil was purchased from Molekula (London, UK). In the treated group, mice (∼33 g) drink ∼5 mL per day (our observations, consistent with general data, see
Blood pressure
At the end of treatment by minoxidil, blood pressure was measured in the tail artery of awake animals placed in a restrainer by using the tail-cuff method (Blood pressure analyzer, MODEL-179; IITC, San Diego, CA). To avoid blood pressure elevation due to the stress induced by restrainment, measurements were repeated during 3 days, three times a day for animal habituation. The values obtained at days 2 and 3 only were averaged for each animal.
Heart weight
Mice were anesthetized by intraperitoneal injection of pentobarbital (60 mg/kg). Hearts were collected, washed, and weighed (wet weight). Left ventricle, right ventricle, and septum were then dissected, washed, and weighed (wet). Ratios of total heart, left ventricle, septum, and right ventricle weights to body weight were calculated to eliminate any influence of interindividual body weight variations.
Surgical procedure and mechanical studies
Animals were anesthetized by intraperitoneal injection of pentobarbital (60 mg/kg). Before heart collection, a segment of the ascending aorta was quickly excised and placed in physiological buffer, and the vessel was then cleaned of connective tissue and fat, cannulated and mounted onto a pressure arteriograph for video-microscopy monitoring, as described previously. 10,31 Below 125 mmHg, the inner diameter (ID) was calculated as described. 10,31,32 Circumferential midwall strain ( = relative increase in diameter), circumferential wall stress (forces that are circumferentially applied on each small portion of the vessel wall), and incremental elastic modulus (Einc, wall stiffness) were calculated according to Gibbons and Shadwick's formulas. 33 Aortic distensibility (relative change in vessel volume in a 25 mmHg pressure increment) was also calculated. 10,31 Arterial reactivity was evaluated at 75 mmHg by measuring the aorta diameter decrease (%) in response to the SMC-dependent vasoconstrictor phenylephrine (5 μM), then the percent diameter restoration of the PE-constricted vessel in response to the endothelial cell-dependent vasodilator acetylcholine (5 μM). 10
Histology
Mice ascending aortae were fixed in 4% paraformaldehyde for one night at 4°C and embedded in paraffin. Five-micrometer sections were deparaffinized with xylene, hydrated in alcohol, and then rinsed in water. Three standard stainings were used: hematoxylin-eosin for cytoplasm and nuclei, picrosirius red for collagen, 34 and Weigert (resorcin-fuschin) staining for elastic fibers. Picrosirius red-stained sections were examined under bright-field illumination for general collagen, or polarized light for fibrillar collagen observations 35 in which collagen I appearing in red-orange and collagen III in green-yellow. 36 In Weigert-stained cross sections of the ascending aortae (1) the number of radially oriented elastic fibers was counted in 90 areas located between the elastic lamellae in each group (6 control and 8 minoxidil-treated animals), and (2) the elastic lamella thickness was measured in 142 different points in each group (5 animals per group).
Electron microscopy
For transmission electron microscopy, abdominal aorta segments were fixed for 2 hours in 3% glutaraldehyde, stored in Milloning buffer (pH 7.3) and postfixed in 2% osmium tetroxide. Once dehydrated in a graded series of acetone, the specimens were embedded in Araldite to obtain thin cuts. These sections were counterstained with lead citrate and examined using a Zeiss-109 transmission electron microscope (Zeiss, Jena, Germany).
RNA analyses
Total RNA was extracted from thoracic descending aorta with the E.Z.N.A.® total RNA Kit I (Omega Biotek, Inc., Norcross, GA) and genomic DNA was digested with DNase I. Gene expression levels were evaluated by real-time PCR using a CFX96® real-time system with the IQ™ SYBR® Green supermix (Bio-Rad, Marnes-la-Coquette, France) after oligodT-primed reverse transcription of 200 ng of total RNA. Expression of mRNAs were normalized to the housekeeping gene hypoxanthine guanine phosphoribosyl transferase (HPRT) mRNA level. Amplification primers
10
are provided in Supplementary Data (Supplementary Data are available online at
Quantification of elastin in the thoracic aorta
After dissection of the descending thoracic aorta and cleaning of blood and surrounding adipose tissue, delipidation, drying (and weighing of the dry tissue), cell protein extraction, and removal of non-elastin extracellular proteins by alkali hydrolysis were performed, as described. 37 Elastin was then quantified by determining the dry weight of the residue as micrograms per millimeter aorta wall and concentration as micrograms per milligram aorta dry weight. 22 A detailed protocol is provided in Supplementary Data.
Quantification of collagen and total protein in the thoracic aorta
Contents in hydroxyproline—representative of collagen—and total amino acids—whose sum represents total proteins—were determined by amino acid analysis of thoracic aorta segments using high-pressure ion-exchange chromatography 10 on a Biochrom 30 amino acid analyzer (Biochrom, Cambridge, UK). Hydroxyproline concentration was also calculated as nanomoles per milligram total protein. Norleucin was used as an internal standard.
Enzyme-linked immunosorbent assay for determination of circulating anti-elastin and anti-tropoelastin antibodies
Alpha-elastin was obtained from aortas of ICR mice as published. 38 Porcine aortic tropoelastin was courteously provided by Prof. Sandberg, Loma Linda University (Loma Linda, CA).
Anti-elastin (AEAb) and anti-tropoelastin antibodies (ATEAb) were assessed by direct Enzyme-linked immunosorbent assay (ELISA), as described. 39,40 A detailed protocol is provided in Supplementary Data. All the samples were analyzed in triplicates and the average was calculated.
Direct determination of AGEs formed in vivo in the thoracic aorta
Soluble α-elastin was obtained from the descending thoracic aorta by the method of Partridge. 38 Insoluble elastin was hydrolyzed 5 times in 0.25 M oxalic acid (OA) in boiling water bath, then dialyzed against phosphate buffered saline (PBS) or distilled water (according needs). OA allows the solubilization of unaltered AGE 39,41,42 and non-AGE 43,44 cross-linked proteins. Although OA potentially interacts with sugars at the early reversible stages of glycation, 42,45 it does not interact with the irreversible and stable advanced glycation products that are studied in our experiments. The presence of nondamaged AGE structures in isolated soluble elastin fragments is proved by their fluorescence at their specific excitation/emission wavelengths. 39,41
Protein content of each sample was determined by measurement of the absorption of ultraviolet light at 280 nm wavelength and calculated according to a standard curve, constructed on the basis of different dilutions of mouse or human α-elastin.
Maillard reaction-related fluorescence, representative of AGEs formed in vivo, was measured by spectrofluorimetry (excitation: 360 nm; emission: 450 nm) 39,41 with a Corning-EEL fluorimeter. Quinine sulfate 1 μM in 0.1 N H2SO4 was used as a standard. The AGE levels were expressed as arbitrary fluorescence units per milligram protein.
In vitro glycation capacity of mouse aortic elastin to form AGEs
Descending thoracic aorta elastin was incubated with 100 mM glucose in 0.2 M PBS, pH 7.8, containing 0.04% NaN3, at 37°C for 4 weeks. 46 Control samples were studied in the same conditions in the absence of glucose. The capacity of elastin to form AGEs in vitro was evaluated by measurement of the Maillard reaction-related fluorescence (FC) of the samples and controls, as described above, from day 1 to day 28 of incubation. The FC difference (sample FC–control FC) was calculated for each day.
Measurement of circulating anti-AGE antibodies
Glycation of keyhole limpets hemocyanin for antigen
Keyhole limpets hemocyanin (KLH) (Sigma-Aldrich, St Louis, MO)—20 mg/mL was glycated in vitro with 3.33 M glucose in 0.4 M PBS, pH 7.5 with preservative 0.04% sodium azide, at 37°C, for 12 weeks. The formation of AGE of KLH (AGE-KLH) was determined via measuring the fluorescence at 360/440 nm excitation/emission. The obtained AGE-KLH was used as antigen in homemade ELISA.
ELISA for determination of circulating anti-AGE antibodies
AGE antibodies (AGEAb) were assessed by homemade ELISA, as described. 46,47 A detailed protocol is provided in Supplementary Data. All the samples were analyzed in triplicates.
Statistics
Comparisons of treated and control mice body weights, heart weight/body weight ratios, mRNA dosages, protein levels, in addition to vessel diameter (outer diameter [OD] and inner diameter [ID]), wall thickness, or incremental elastic modulus (Einc) in response to transmural pressure or vasoactive agonists were assessed using multivariate analysis of variance (MANOVA), or one- or two-way analysis of variance (ANOVA) followed when necessary by Fisher's least significant difference test for paired value comparisons. The results are presented as mean values ± standard error of the mean, and p-values ≤0.05 were considered as statistically significant.
Results
Body and heart weights, blood pressure
Body and heart masses were compared as a function of treatment (one-way ANOVAs). Minoxidil induced a trend toward elevation of body weight (34.5 ± 0.6 vs. 33.3 ± 0.7 g, p = 0.2) and a statistically significant increase in total heart weight (132 ± 2 vs. 121 ± 2 mg) (Fig. 1A, B). Total heart weight to body weight ratio only presented a nonsignificant trend toward elevation following minoxidil treatment (one-way ANOVA, p = 0.24, Fig. 1C). This mitigated observation was explained by the different effects of minoxidil on the different cardiac parts. While left ventricle/body weight and right ventricle/body weight ratios were increased by 6%–8% by minoxidil treatment (Fig. 1D), no significant effect of minoxidil on the septum/body weight ratio could be detected (one-way ANOVA, p = 0.4, Fig. 1E). This suggested that chronic minoxidil treatment in aged mice presents detrimental side effects on the cardiac physiology that are milder than those observed in other rodents or humans, which include weight gain and cardiac hypertrophy. 48 –55
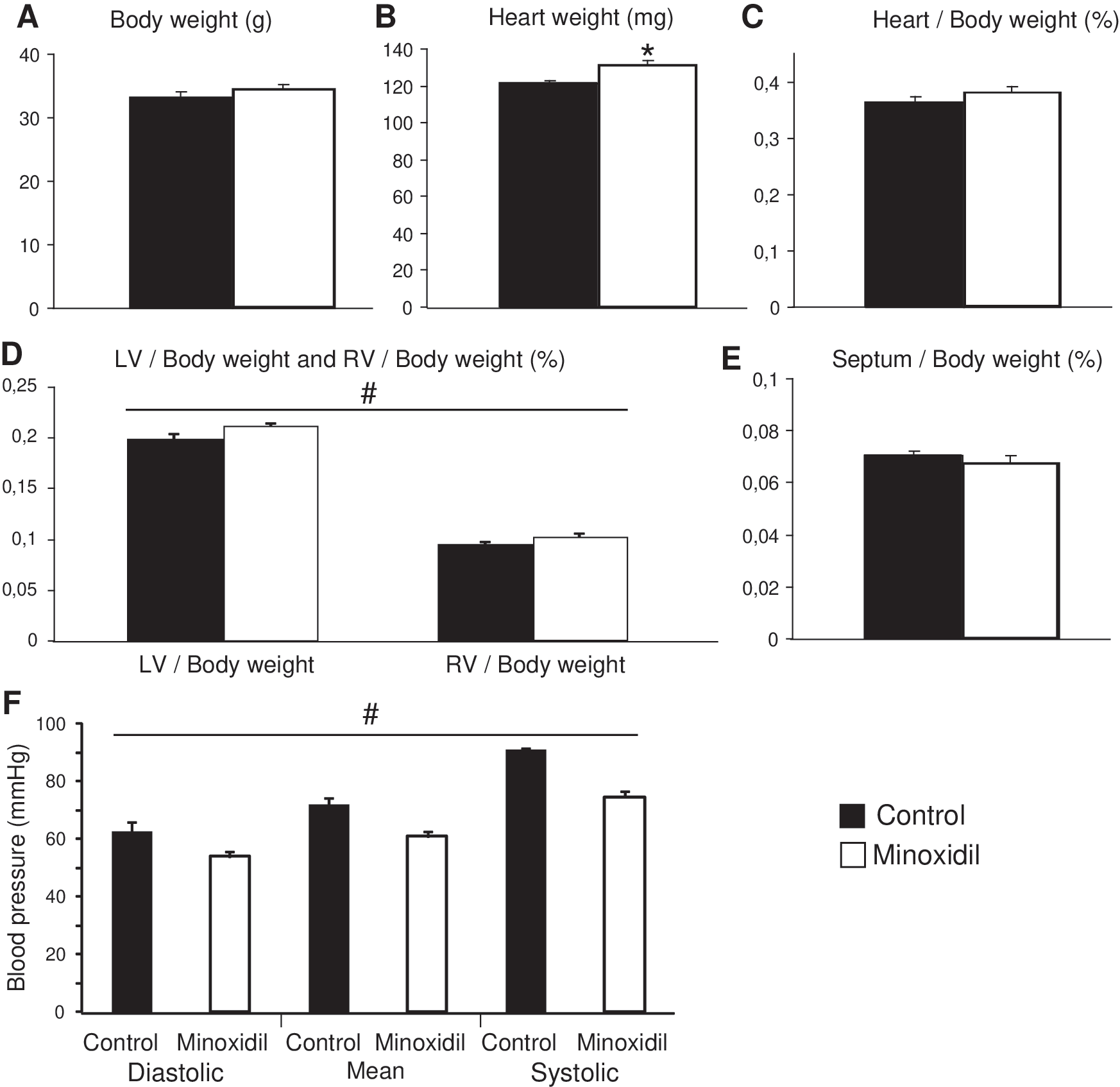
Effect of minoxidil on body weight, heart weight, and blood pressure. Body and heart weights (
As expected and in accordance with previous results obtained in humans and young rats and mice, 24,26,56 minoxidil treatment also significantly decreased blood pressure of aged mice by 14%–18% (Fig. 1F).
Ascending aorta histology
The overall morphology of the ascending aorta was unchanged by minoxidil (Fig. 2 A, B). The media contained seven concentric elastic lamellae, sometimes disrupted, in both minoxidil-treated and control mice. Nevertheless, many additional radially oriented elastic fibers, often bridging the circumferential elastic lamellae, appeared in the aorta wall of minoxidil-treated mice, compared to controls: 7.92 ± 0.66 versus 2.25 ± 0.32 radial elastic fibers/1000 μm2 inter-lamellar space (one-way ANOVA, p < 0.000001, Fig. 2C, D). Minoxidil also increased the average elastic lamella thickness by 13% (Fig. 2C, D, I), suggesting that some of the neosynthesized elastic fibers are also deposited onto preexisting fibers/lamellae. No clear change in the overall collagen (Fig. 2E, F) or fibrillar collagen (Fig. 2G, H) distributions could be observed in response to minoxidil treatment. Nevertheless, less fibrillar collagen type I and more fibrillar collagen type III were detected in the ascending aorta wall from minoxidil-treated mice, compared to controls (Fig. 2G, H). These results indicated for the first time that chronic minoxidil is able to induce neosynthesis of elastic fibers in aged animals.
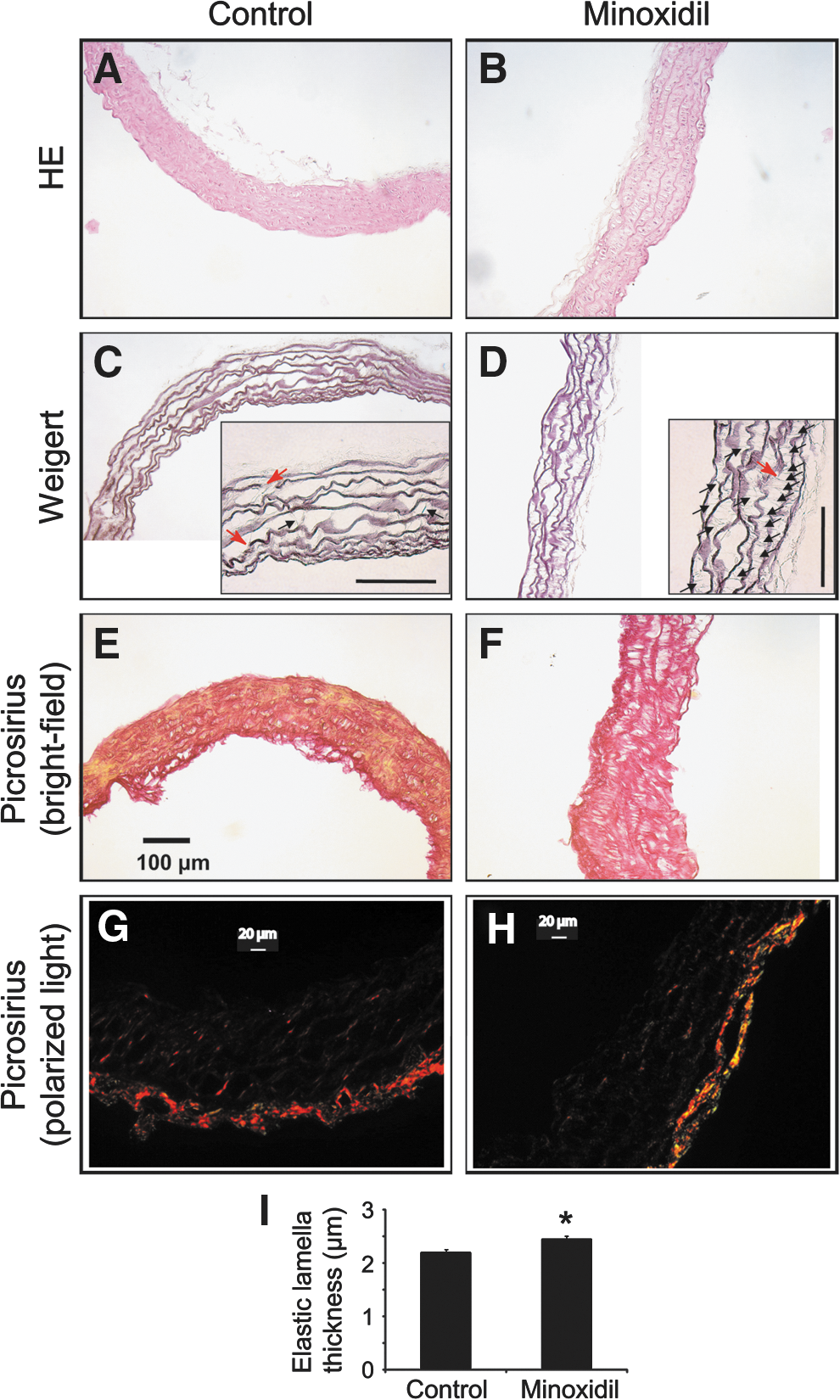
Histology of the ascending aorta. Cross sections of the vessels from aged mice were stained with HE
Aorta ultrastructure
Electron microscopy confirmed the presence of additional neosynthesized thin elastic fibers in the aorta of minoxidil-treated mice. While these elastic fibers were virtually not observed in control animals (Fig. 3A), additional thin elastic fibers were observed after minoxidil treatment, either radially oriented (i.e., perpendicular to elastic lamellae), coiled or crossing elastic lamellae or aligned to the circumferential elastic lamellae (Fig. 3B). The existence of elastic fibers aligned to or crossing the elastic lamellae fits with the histological finding of thickened elastic lamellae after minoxidil treatment. This supports the idea that, besides inter-lamellae radially oriented neofibers, an appreciable part of the neosynthesized elastic fibers are more properly oriented and aggregated to preexisting elastic lamellae.

Ultrastructure of the abdominal aorta wall. Transmission electron microscopy images of vessels from control
Levels of collagen and elastic fiber component-related mRNAs
To estimate the impact of minoxidil on gene expressions, the mRNAs of the major contributors to elastic and collagen fibers were quantified in the thoracic aorta at different time points: 1, 4, and 10 weeks of minoxidil treatment. Interestingly, tropoelastin, LOX, and fibulin-5 mRNA levels presented a time-dependent elevation during the treatment, with a significant peak (2x–3x) at 4 weeks, before returning to control levels at the end of treatment (Fig. 4A–C). No significant effect of minoxidil on fibrillin-1, LOXL-1, and collagen-I mRNA levels could be detected, compared to controls (one-way ANOVAs, at 1-, 4-, and 10-weeks of minoxidil, respectively: fibrillin-1: p = 0.772, 0.552, 0.886; LOXL-1: p = 0.794, 0.281, 0.784; collagen-I: 0.137, 0.085, 0.693; Fig. 4D–F). Fibrillin-2 mRNAs were undetectable in the aorta of these old animals.
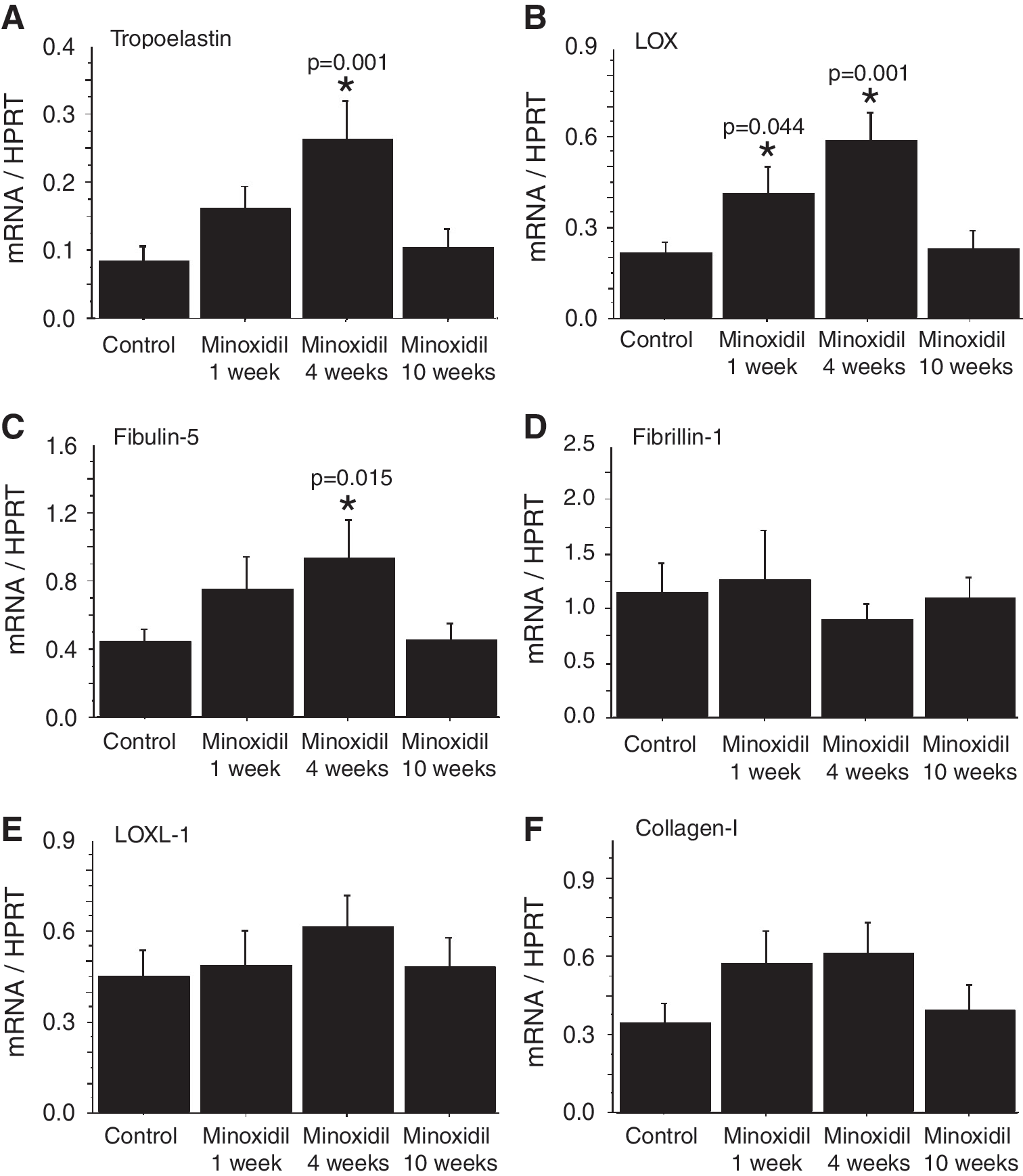
Extracellular matrix-related mRNAs in the descending thoracic aorta of aged mice. The mRNA levels of major constituents of elastic and collagen fibers, and cross-linking enzymes involved in the assembly of elastic and collagen fibers, were measured in the aorta at three different time points: 1, 4, and 10-weeks of minoxidil treatment. The values were normalized to the housekeeping gene HPRT mRNA levels. Tropoelastin
Elastin, collagen, and AGE metabolism in the thoracic aorta
Aortic elastin content and levels of circulating antibodies to tropoelastin—representative of ongoing elastin synthesis processes—or AGEs 40,46 were compared as a function of treatment by MANOVA. Minoxidil treatment had a general significant effect on the joint distribution of these three variables, that is a moderate increase in aortic elastin content (+7%) and circulating ATEAb levels (+13%), and a decrease in circulating anti-AGEAb (−39%) (Fig. 5A–C). Minoxidil treatment did not modify aortic wall hydroxyproline content, indicative of collagen (one-way ANOVA, p = 0.884, Fig. 5D), and the levels of circulating antibodies to elastin, markers of ongoing elastin degradation, were not significantly modified (data not shown). Minoxidil also induced a significant decrease in elastin concentration (Fig. 5E), a strong trend toward reduction of hydroxyproline (i.e., collagen) concentration (one-way ANOVA, p = 0.122, Fig. 5F), and an elevation of the aorta dry weight per length unit (99.6 ± 4.1 vs. 148.9 ± 5.0 μg dry aorta per millimeter length for control and minoxidil-treated, respectively, one-way ANOVA, p < 0.00001). Since collagen and elastin concentrations were dropping, the increase in aortic tissue amount apparently relies on non-elastin non-collagen tissue production, that is, secretion of other ECM components and/or cellular hypertrophy. A slight trend toward minoxidil-induced decrease in aortic elastin-associated AGE content was also observed (one-way ANOVA, p = 0.625, Fig. 5G). Finally, the in vitro glycation capacity of aortic elastin was investigated. Minoxidil treatment substantially decreased by ≈20% the capacity of elastin to form AGEs in the presence of glucose over time (Fig. 5H). This protective effect of minoxidil is of particular interest, given the important detrimental processing of macromolecules by non-enzymatic glycation occurring during aging.
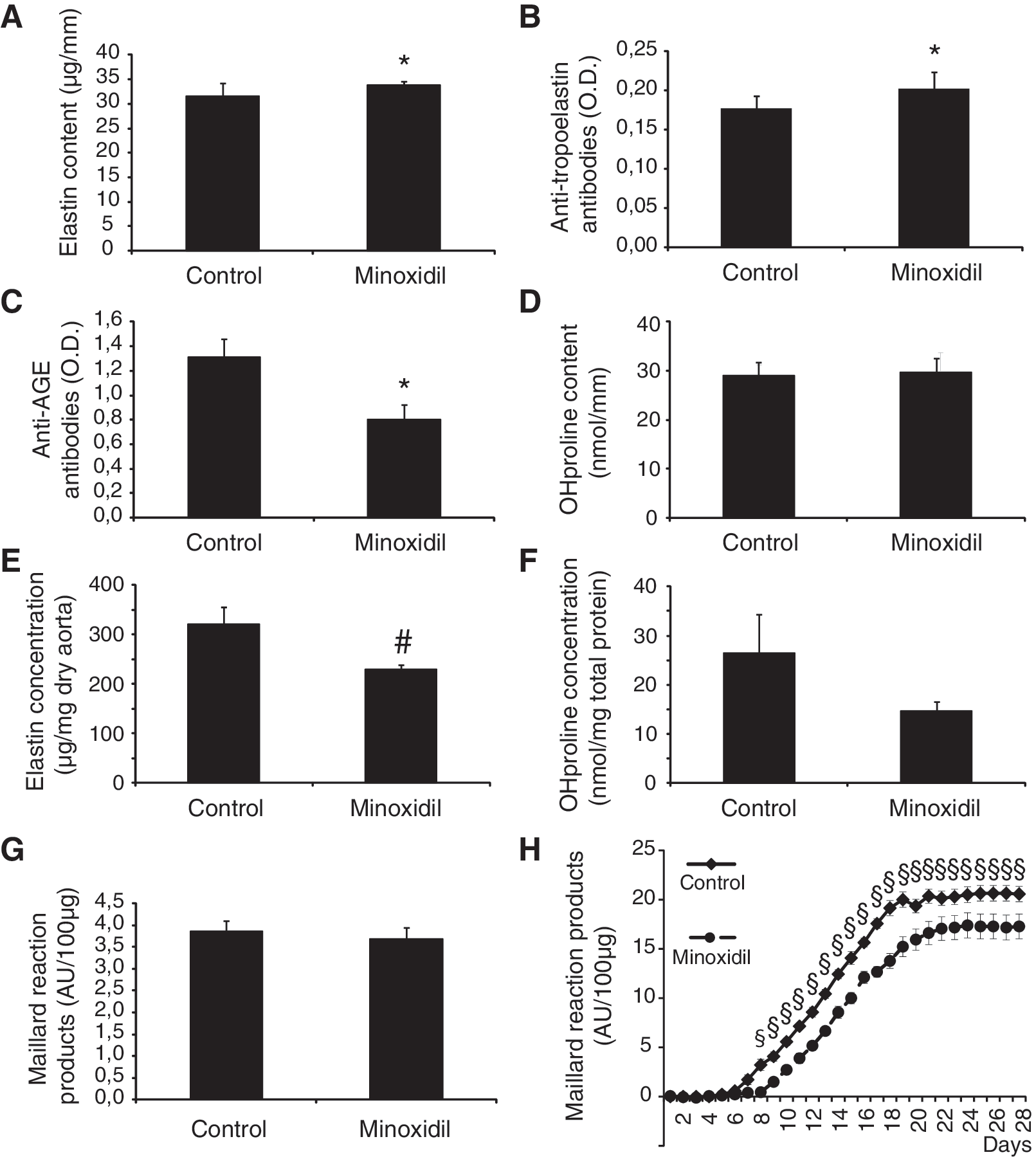
Elastin, collagen, and AGEs in the descending thoracic aorta, and related circulating antibodies. Aortic elastin content
Mechanics of the ascending aorta
Since the above described structural changes in the aorta may have important functional consequences, we addressed the impact of minoxidil on the ascending aorta mechanics and responsiveness. During normal aging, the diameter and wall thickness generally increase in the ascending aorta. 10,57 Here, in aged mice after minoxidil treatment, the OD and ID of the ascending aorta were even larger than those of both young and aged control mice (Fig. 6A, B), while the wall thickness thinned and returned closer to the value measured in young untreated animals (Fig. 6C).
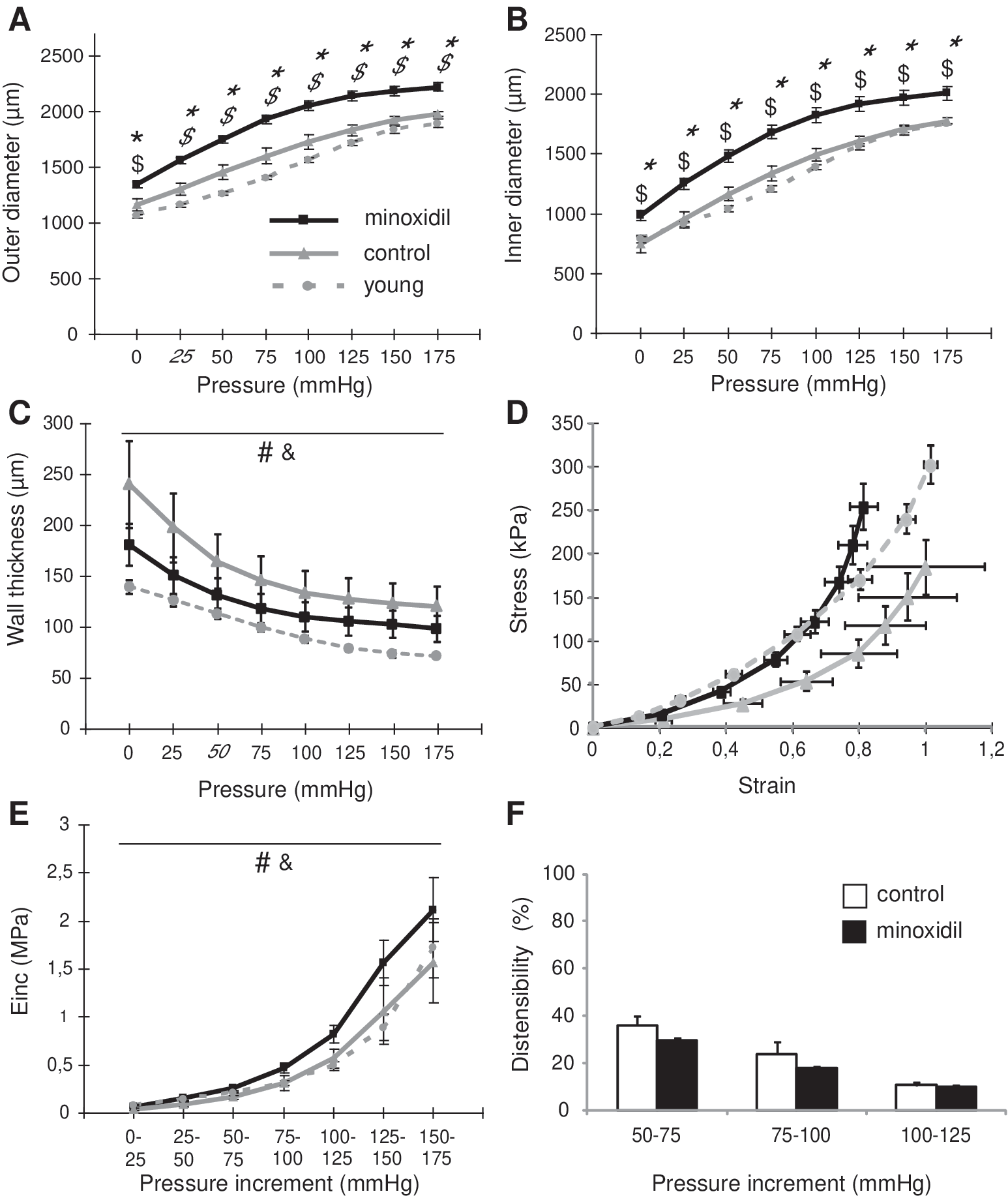
Diameters, wall thickness, and mechanical parameters of the aorta. The OD
The circumferential stress values were increased at any given strain, due to the minoxidil-decreased aortic wall thickness in these aged animals. This shifted the strain-stress relation in minoxidil-treated aged animals closer to that found in younger adult mice, the two curves being virtually superimposed for most of the tested pressures, in particular in the physiological pressure range (Fig. 6D). Unexpectedly, vessel stiffness (Einc) was generally found higher in minoxidil-treated mice than in young and aged controls (Fig. 6E). However, in the physiological blood pressure range (50–125 mmHg), Einc values were close and the aortic distensibility in the physiological blood pressure range was not significantly modified by minoxidil (one-way ANOVAs, p = 0.073, 0.118, and 0.665 for the 50–75, 75–100, and 100–125 mmHg ranges, respectively, Fig. 6F).
The effects of the VSMC-dependent vasoconstrictor phenylephrine (PE) and the endothelium-dependent vasodilator acetylcholine (Ach) on the ascending aorta reactivity were investigated. Minoxidil treatment of aged mice significantly limited the PE-induced vasoconstriction and Ach-induced vasodilation (Fig. 7).
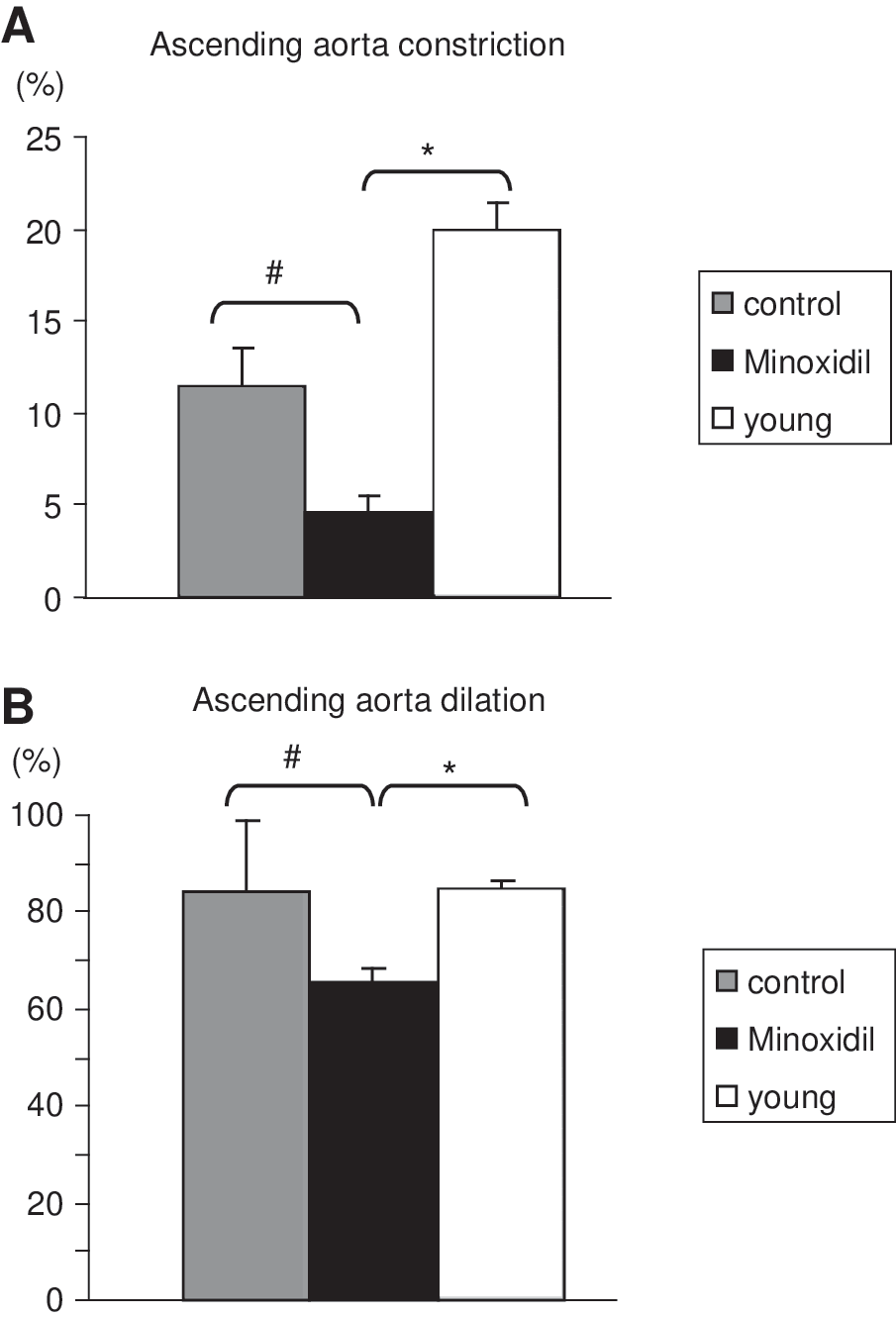
Reactivity of the ascending aorta. Because the initial diameters were different in all the animal groups, the responses to the vasoactive agents are presented in percentage of the initial diameter at 75 mmHg: 5 μM phenylephrine-induced vasoconstriction
Discussion
Minoxidil activates elastin expression in VSMCs in vitro and in vivo in the aorta after chronic administration in young rats, through the involvement of calcium- and ERK1/2-dependent mechanisms, and an increase in tropoelastin transcription and mRNA stability. 25 –28,58 We therefore postulated that chronic minoxidil treatment could also trigger neosynthesis of elastin and elastic fibers in the arteries of aged animals, in which elastic fibers are substantially degraded and elastin production is virtually abolished. 10,59 Such effect may lead to an improvement of the structure/function of aging arteries. Our present investigation showed that minoxidil treatment was able to reactivate elastin synthesis in aged mice, as shown by the observation of increases in tropoelastin mRNA level, aortic elastin content, and circulating ATEAb. 40 This was supported by the transient elevation of mRNA levels of key contributors to elastic fiber assembly—LOX and fibulin-5—during the course of minoxidil treatment. Since nicorandil, another KATP channel opener, was previously shown to increase the relative elastin content of the rat aorta only in young adults, not in aged animals, 29 the present results are of particular importance.
Emphasizing these findings is the fact that numerous additional neosynthesized elastic fibers—close to four times more—were observed in the aorta wall of aged minoxidil-treated mice. This shows that minoxidil not only induces an increase in elastin production but also triggers the complex multimolecular processes that are necessary for the assembly of elastic fibers. 60 While histological observation suggested that new elastic fibers were radially oriented, bridging consecutive elastic lamellae, electron microscopy studies showed that the neosynthesized fibers could have diverse orientations, including alignment along the circumferential elastic lamellae. This unusual disposition of some newly synthesized elastic fibers could partly counteract a stiffness decrease favored by the increase in elastin content induced by minoxidil. This, together with the minoxidil-induced enlargement and increase in tissue amount of the aorta—leading to decreased elastin and collagen concentrations—could explain the slight increase in aortic stiffness at physiological blood pressures without significant change in aortic distensibility after minoxidil.
The presence of diversely oriented neosynthesized elastic fibers could result from the loss during aging of the physiological mechanisms (mechanical stress, chemical signals, normal VSMC orientation/shape, or inteaction with ECM) circumferentially orienting the normal assembly of small elastic fibers and their normal aggregation into large circumferential elastic lamellae. However, as an alternative hypothesis to such a random side effect, production of small radially oriented elastic fibers could be a beneficial way to compensate for the age-dependent alteration of the mechanical properties of the arterial wall. Supporting this hypothesis is the observation of similar radially oriented thin elastic fibers in the aorta of mice genetically deficient for protocadherin-12, an intercellular junction protein, in which arterial structure and mechanics are also altered. 61
Additionally, minoxidil reversed in part the age-dependent wall thickening observed between 6- and 24-months of age. This shifted the ascending aorta strain-stress relation of minoxidil-treated aged mice close to that of untreated 6-month-old animals, confirming at the mechanical level an antiaging action of minoxidil treatment.
Unexpectedly, another particularly interesting effect of minoxidil treatment was also uncovered regarding the metabolism of AGEs, known for their deleterious effects on blood vessels. 62 –64 Minoxidil treatment decreased circulating anti-AGEAb, possibly slightly decreased the aortic content of AGEs, and limited the capacity of elastin to form AGEs. The latter finding could be explained by the known activation by minoxidil of LOX, the enzyme responsible for the formation of elastin cross-links between lysine residues, 65 as confirmed at the mRNA level in our experiments. It is known that increased LOX activity leads to the formation of more ECM cross-links 66,67 and uncross-linked lysine residues are present in mature elastin. 68 Therefore, minoxidil-induced increase in LOX activity could induce the formation of more desmosin cross-links, leave elastin with less free lysine residues—important targets for elastin glycation 69 —available for non-enzymatic glycation, and therefore limit the capacity of elastin to form AGEs.
Regarding the cellular part of the aorta wall function, minoxidil treatment generally decreased the cell response to vasoactive agents, possibly because of an inhibitory effect of the permanent K+ channel stimulation imposed to the vascular cells by minoxidil over the 10 weeks of treatment. This desensitization hypothesis is supported by the previous finding that, in rats, 2-week chronic oral administration of another ATP-dependent K+ channel opener, levcromakalim, attenuated the vasorelaxant effects of a NO-donor. 70
Our study definitively shows that, in aged animals, minoxidil is capable of reinducing aortic elastin and elastic fiber production, protecting elastin against glycation, and reversing some deleterious evolution of vascular mechanics (see synthesis schematic in Supplementary Data). Such reversal of several adverse effects of arterial aging suggests that minoxidil could be seen as a multitarget antiaging molecule. The next step will be to find a way to assemble more properly oriented neo-elastic fibers, possibly though manipulating some of the major contributors to this mechanism (e.g., LOX, LOXL, and fibulin-5).
Footnotes
Acknowledgments
The authors thank the financial support of the European Commission (contract ELAST-AGE, 6th PCRD, number LSHM-CT-2005-018960), the Association Française contre les Myopathies, and the Association “Autour des Williams.”
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.