
Abstract
The proliferation and differentiation potential of adipose-derived stem cells (ADSCs) decline with aging. Moreover, Alzheimer's disease is associated with progressive decline in cholinergic neurons. The purpose of this study is to enhance the proliferation potential of aged rat ADSCs and their differentiation into cholinergic neurons. The ADSCs were collected from aged male rats cultured and treated with different concentrations of sodium selenite for 3 days or glutathione mono ethyl ester (GSH-MEE) for 1 day. Incubating the ADSCs with 27 nM sodium selenite for 3 days significantly increased the relative cell proliferation, compared with the control, without any change in the telomerase activity, the related telomerase gene expression, and the telomere length, but it does improve differentiation of the aged ADSCs to cholinergic neuron-like cells. GSH-MEE at a concentration of 2 mM for 1 day resulted in increased relative cell proliferation, but it did not change the telomerase activity, the related telomerase gene expression, the telomere length, and differentiation potential. Sodium selenite is more effective than GSH-MEE in improving the aged ADSCs' properties. However, both did not have any effect on telomerase activity.
Introduction
A
The degeneration of cholinergic neurons essentially contributes to the cognitive loss and pathogenesis of AD, proposing that cholinergic neurons might be a feasible type of cells to alleviate the cognitive symptoms related to AD. 5 Thus, production of cholinergic neurons from stromal cells would offer a therapeutic strategy for AD. Producing specific cell types from a patient's own stem cells would have immunologic advantages. Adipose-derived stem cells (ADSCs) are easily available in great amounts with minimal invasiveness. 6 However, the number, proliferation, and differentiation potential of ADSCs decreased with aging. 7 –9 In addition, telomere shortening has been observed in aged ADSCs 10 ; telomere shortening is known as an age-related chromosomal change that occurs in most human tissues, emerging as a potential contributor to the pathogenesis of AD. 10
Impairment of ADSCs' properties with age may cause lower potency of autologous cell therapy in aged patients. Impairment of ADSCs' properties with age may cause lower potency of autologous cell therapy in aged patients. There is need for approaches to optimize the performance of aged ADSCs to use it in autologous transplantation. There are some approaches reported in the literature aiming to improve the proliferation and differentiation capacity of stromal cells. Liu et al. reported that pretreatment with SRT1720 improves the survival of aged bone marrow stromal cells (BMSCs) and enhances their therapeutic efficacy for rat myocardial infarction, 11 while Sun et al. showed that exposing aged BMSC to young extracellular matrix improved replication and osteogenesis of aged mesenchymal stem cells. 12 Others reported that the in vitro culturing of ADSCs with a growth medium low in calcium and supplemented with the antioxidants improved their growth, 13 while Sun et al. documented that exposure of ADSCs treated with N-acetyl-L-cysteine and L-ascorbic acid-2-phosphate promoted their entry into the S phase of the cell cycle resulting in rapid cell proliferation, and the telomere length was increased. 14 We utilized glutathione mono ethyl ester (GSH-MEE) and sodium selenite to improve biological properties of aged ADSCs. These substances have antioxidant properties. Oxidative stress (OS) plays an important role in aging. 15 OS is defined as the condition when the sum of free radicals in a cell exceeds the antioxidant capacity of the cell; 16 nevertheless, it occurs in the natural course of aging, while younger cells can overcome OS by increasing the production of defensive antioxidants. 17 The main endogenous antioxidant, GSH, has been shown to decline with aging 18,19 and this was linked with neurodegenerative diseases. 20 Alteration of GSH which is involved in OS was shown to play an important role in aging and scavenging. 21 Moreover, the activities of glutathione peroxidase (GPx), a group of enzymes that are important for reducing hydrogen peroxide to water, are reduced in old guinea pigs. 22 They are selenocysteine enzymes that use GSH as a reducing agent. 23
Furthermore, previous studies showed that GSH and selenium regulate telomerase activity in 3T3 fibroblasts and in rat hepatocytes. 24,25 Selenium is necessary for neuroepithelial differentiation and survival too. 26 GSH has an important role in improving neuron differentiation in the thalamus of HIV-1Tg rats. 27 Our previous results showed that GSH-MEE improved aged rat BMSC properties. 28 Therefore, this study was carried out to investigate the effect of GSH-MEE and sodium selenite on the proliferation rate, telomerase activity, telomere length, and gene expression levels of several proteins related to telomerase activity during the in vitro expansion of aged rat ADSCs. The effect of these two substances on aged ADSC differentiation to cholinergic neuron-like cells (CNLCs) was also assessed.
Materials and Methods
Cell culture
In this study, for experimentation, consent was given by the Ethics Committee of the Tarbiat Modares University (Tehran, Iran) according to the guidelines of the Helsinki ethical code regarding experiments performed on animals. ADSCs were acquired from three aged (18 months) 29 and three young (3 months) male Sprague-Dawley rats and cultured. The animals were anesthetized under sterile condition, and the adipose tissue was obtained from the pararenal region. For sample of every rat cultured separately, the ADSCs were not pooled. The tissue was processed based on the established method with minor modifications. 30 Briefly, the samples were washed several times in phosphate-buffered saline containing penicillin and streptomycin. Thereafter, the tissue was minced and treated with equal volumes of 0.075% collagenase type1 (Sigma, Belgium) with continuous agitation at 37°C for 30 minutes. Enzyme activity was neutralized with 10% fetal bovine serum (Gibco, United Kingdom) and then centrifuged at 2700 rpm for 10 minutes to obtain a high-density cell pellet. The cell pellet was cultured. The medium was replaced every 2 days. To characterize the isolated ADSCs which were immunostained with mouse primary antibodies, including anti-CD49d, anti-CD29, anti-CD90, anti-CD105, 31 anti-CD34, and anti-CD106, the isolated ADSCs were labeled with rabbit anti-mouse secondary antibody conjugated with FITC (all purchased from Abcam, Cambridge Science Park, Cambridge, United Kingdom) and counterstained with propidium iodide (PI: Sigma-Aldrich, Germany). Finally, they were examined under a fluorescence microscope (Olympus IX71, Olympus, Japan).
Trypan blue exclusion assay
The aged ADSCs (third passage) were seeded at concentration of 2 × 104 cells/well into 12-well plates and treated with different concentrations (0, 1, 2, and 3 mM) of GSH-MEE (Santa Cruz Biotechnology, Texas) for 24 hours, whereas sodium selenite was used at various concentrations (0, 27, 37, 52, 70, 100, 140, 200, 270, and 380 nM) for 3 days. Triplicate wells were used, and each experiment was repeated thrice. The growth rate of ADSCs in different groups was measured by direct cell counting and then calculated by formula that has been explained previously. 32 Cells were counted using trypan blue dye.
The optimum concentration of GSH-MEE and sodium selenite was detected based on the results obtained from the cell viability studies.
Gene expression assessment and absolute telomere length measurement
The reverse transcription quantitative polymerase chain reaction (RT-qPCR) was used in assessing the expression level of the related telomerase activity genes, while the absolute telomere length was evaluated by qPCR, in the ADSCs at all groups. We used a CFX96™ system (Bio-Rad, Germany). The sequences of the primers used in this study were reported in Aminizadeh et al. 28
The ΔΔCt method was used to determine the relative expression levels, which was normalized to β-Actin level. Absolute telomere length was measured as previously described. 33 Each sample was analyzed in duplicate, and all of the measurements were performed in the presence of standard samples and nontemplate controls, in which the DNA was replaced with water. Standard curves were plotted by performing 10-fold serial dilutions of synthesized oligonucleotides containing only TTAGGG repeats for telomere PCR and an oligomer containing 36B4 PCR product. 28
Telomerase activity assay
The telomerase activity of the ADSCs in the protein extracts was assessed by the TeloTAGGG Telomerase PCR-ELISA Kit according to the manufacturer's instructions. The lysis buffer and the heat treated cell extract served as the negative controls. All samples were performed in triplicate.
ADSCs' differentiation to CNLCs
The ADSCs at third passage were plated on nonadherent plastic flask at a density of 3 × 105 cells/cm2 in neurosphere formation medium (DMEM/F12 medium with 2% B27 [Invitrogen, Germany], 20 ng/mL basic fibroblast growth factor [bFGF: Sigma-Aldrich, Germany], 20 ng/mL epidermal growth factor [EGF: Sigma-Aldrich, Germany], 100 U/mL penicillin, and 100 mg/mL streptomycin [GIBCO, Germany]). Neurosphere-like structures were observed after 2 days, which were cultured for 7 days. Seven days after plating, the neurospheres were harvested with 0.25% trypsin/1 mM EDTA and plated in six-well culture plates in DMEM/F12 medium supplemented with 5% fetal bovine serum, 10 ng/mL EGF, 10 ng/mL bFGF, 2% B27, 100 U/mL penicillin, and 100 mg/mL streptomycin. The cells were maintained for 5 days, immunostained for nestin and neurofilament 68(NF 68) neural markers. The nerve growth factor (NGF: Sigma-Aldrich, Steinheim, Germany) was utilized as cholinergic neuron inducer. NGF concentration in differentiation medium was increased by 10 ng/mL, both EGF and bFGF were reduced by 1 ng/mL, and B27 was reduced by 0.1% (the final concentrations of NGF, EGF, bFGF, and B27 were 100, 0, 0 ng/mL, and 0%, respectively). After 14 days, immune-staining was carried out for ChAT, NF200, NF160, synapsin, and MAP2. All of the immune cytochemical experiments were performed in triplicate; three fields per well were randomly chosen and examined by fluorescence microscope. The total number of cells per field was determined under fluorescence microscope by counting the total nuclear number, followed by counting the number of immunoreactive cells. The average percentage of the immunoreactive cells was determined from all fields.
Statistical analysis
The statistical analysis was performed using SPSS software release 16. The results are expressed as the mean ± standard error. The normality of the data was assessed by the Kolmogorov–Smirnov test (K-S test). The data of the cell growth were analyzed by ANOVA and Tukey's post hoc test. Comparison was done by Student's t- test. The level of statistical significance was set at p < 0.05.
Results
Surface markers of ADSCs
ICC results show that ADSCs were immunoreactive to CD49d, CD29, CD90, and CD105. They were negatively immunostained with antibodies against CD34 and CD106 (Fig. 1). The percentages of immunoreactive cells (PIC) were 98.12% ± 0.56%, 96% ± 0.37%, 95.18% ± 0.42%, and 92.12% ± 0.34%, respectively. They were negatively immunostained with antibodies against CD34 and CD106 and the PIC were trivial.

Represents the characterization of the adipose-derived stem cells (ADSCs) isolated from aged rats using immunostaining of the cells by the following mouse monoclonal primary antibodies: Anti-CD49d, anti-CD29, anti-CD90, anti-CD105, anti-CD34, and anti-CD106 mouse antibodies. Nucleus counterstained with propidium iodide (PI). (Scale bar = 200 μm, all). The percentages of immunoreactive cells (PIC) were 98.12% ± 0.56%, 96% ± 0.37%, 95.18% ± 0.42%, and 92.12% ± 0.34%, respectively. They were negatively immunostained with antibodies against CD34 and CD106 and the PIC were trivial.
Effect of sodium selenite and GSH-MEE on the ADSC growth rate
The effect of sodium selenite on the ADSCs' growth rate is shown in Figure 2A. The highest rate of growth was observed in the cells treated with 27 mM sodium selenite, which was significantly high compared to the controls. Sodium selenite at concentration of 138 mM significantly decreased the cell growth.

The effect of GSH-MEE on the ADSCs' growth rate is shown in Figure 2B. GSH-MEE in 2 mM concentration significantly improved the growth rate in comparison with those of the control, but 1 and 3 mM concentration of GSH-MEE did not change the growth rate.
Effect of sodium selenite and GSH-MEE on the relative expression of telomerase activity genes and the telomerase activity and the telomere length
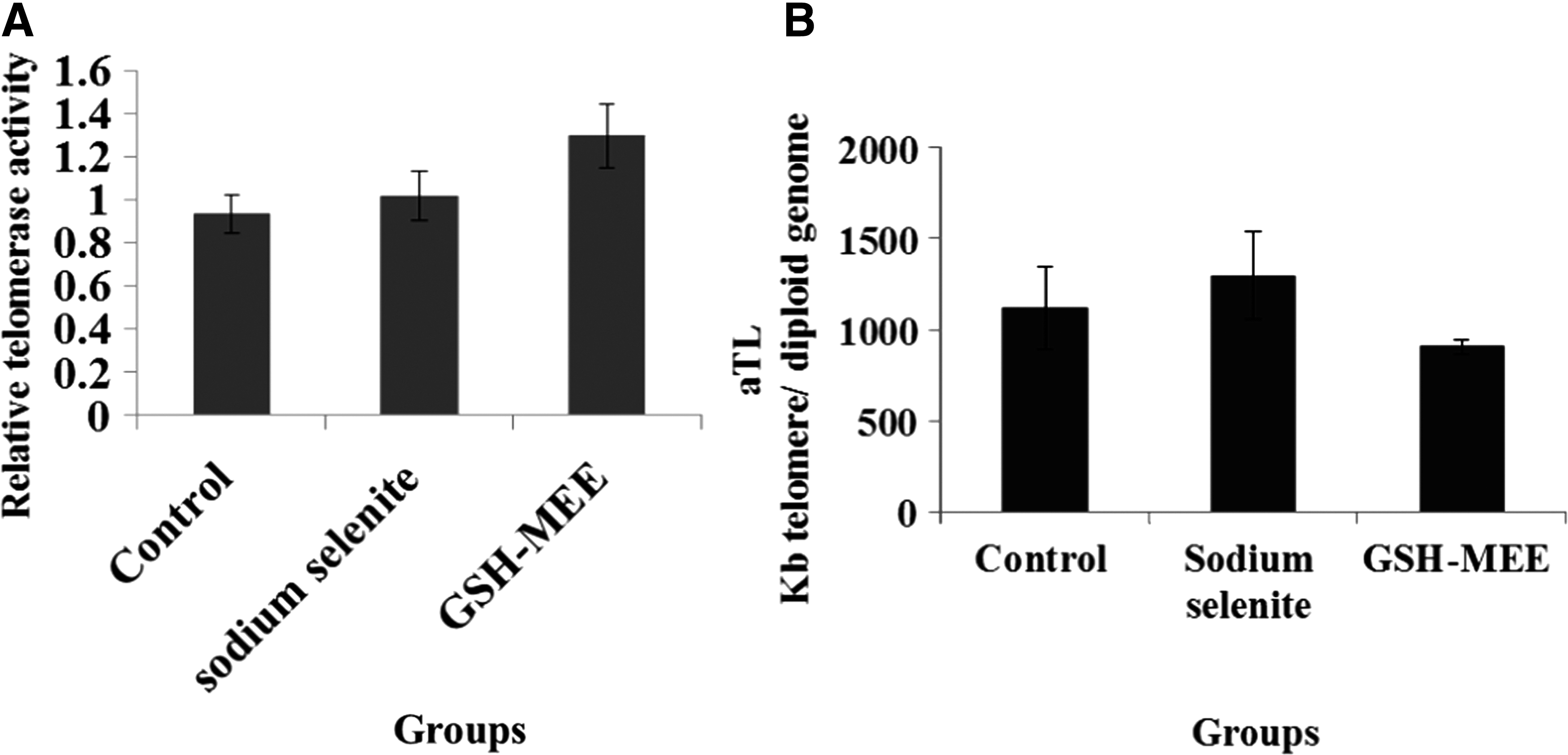
Telomerase activity genes include shelterin complex (TERF1, TERF2, POT1, TPP1, TIN2, RAP1), CTS complex [CTC1, OBFC1 (STN1), TEN1], glyceraldehyde-3-phosphate dehydrogenase (GAPDH), and components of telomerase enzyme (TERT, TERC). Sodium selenite and GSH-MEE did not significantly change the expression level of these genes (Fig. 3); also they did not alter telomerase activity (Fig. 4A) and telomere length (Fig. 4B) in aged ADSCs.

Represents the expression level of the related telomerase activity genes in the ADSCs at all groups. Sodium selenite and GSH-MEE did not significantly change the expression level of these genes. Data represent as the mean ± SE.
Expression of neural markers in ADSC-derived NSCs and CNLCs
NSCs demonstrated positive reaction to anti nestin and NF68 antibodies (Fig. 5A). There is a significant difference between these protein expressions in young and aged rat ADSC-derived NSCs. Sodium selenite increased expression of both nestin and NF68 in NSCs generated of aged ADSCs (Fig. 5B).
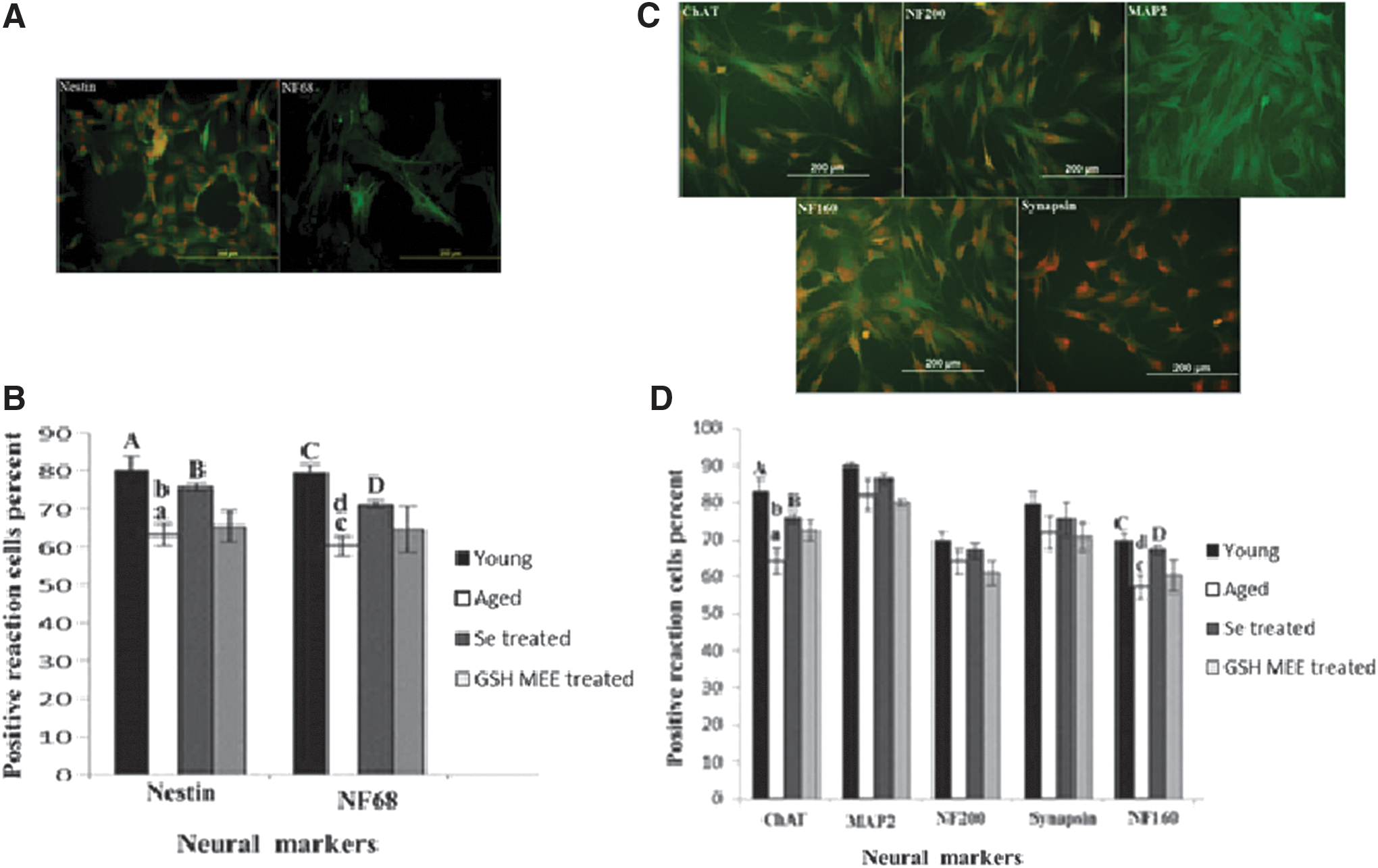
CNLCs have positive reaction to ChAT, NF160, NF200, MAP2, and synapsin (Fig. 5C). Positive reaction cells to ChAT and NF160 were significantly lower in aged cells in comparison to young cells; sodium selenite significantly increased expression of ChAT and NF160 in CNLCs generated of aged ADSCs (Fig. 5D). GSH-MEE did not change expression of neural markers in aged ADSC-derived NSCs and CNLCs.
Discussion
In the present study, the data have shown that sodium selenite significantly increased the growth rate of aged rat-derived ADSCs. This result was consistent with those of other studies. Alves et al. showed that selenium increased the growth rate of BMSCs 34 ; moreover, Park et al. 35 reported that selenium stimulated the proliferation of 3T3-L1 preadipocytes although Ebert et al. did not find a significant change in doubling time of the human selenium-supplemented BMSCs compared to those of the untreated controls. 36 In addition, in this study, GSH-MEE significantly increased the growth rate of aged rat-derived ADSCs. Previous studies showed that GSH concentrations increased at the beginning of the logarithmic growth phase in IEC-6 intestinal epithelial cells. 37 In addition, we previously showed that GSH-MEE could increase the relative cell proliferation of aged rat-derived BMSCs. 28
In this study, despite the increase in the growth rate of the aged rat-derived ADSCs by sodium selenite and GSH-MEE, unexpectedly the telomerase activity and telomere length remained unchanged. This can be explained on the basis of the findings of others, which showed that mesenchymal stem cells (MSCs) were able to proliferate quickly without increasing their telomerase activities. 38 Other investigators' reports confirmed that selenium upregulated the telomerase activity of young mouse lymphocytes; 39 however, Murillo-Ortiz et al. showed that aged cells are different from young cells about telomere length response, so that Concanavalin A increased the telomere length in BMSCs obtained from young men, but it did not change the telomere length of the aged cells. 40 In addition, Son et al. observed that the capability for induced telomerase expression in T and B cells is diminished with age. 41
Our results showed that the differentiation potential of aged rat ADSCs to CNLCs was significantly lower than the ADSCs of young rats. There are some conflicting reports about the effects of aging on the differentiation potential of ADSCs. Zhu et al. demonstrated that the osteogenic potential of ADSCs was significantly reduced by aging, 42 although Khan et al. did not find any significant difference between aged and young ADSCs' osteogenic differentiation potential. 43 Furthermore, aged ADSCs secrete fewer proangiogenic factors compared to the young cells, and, as a result, their angiogenic potential declines. 44 To the best of our knowledge, there is no report about differentiation of aged ADSCs to CNLCs, and this study is the first such report. Further investigation is needed to confirm the data in geriatric human samples.
In addition, results of this study showed that sodium selenite increased the differentiation potential of aged rat ADSCs to CNLCs. Other reports showed that selenium promotes the differentiation and development of NSCs of the newborn rat. 45 OS plays a major role in the physiological decline of major body systems during aging. 46 Maintenance of intracellular redox homeostasis is dependent on antioxidant factors. 47 Selenium is an essential trace element with great capacity for regulating antioxidant defense. 48 The biological effects of selenium are chiefly due to its integrating into the selenoproteins. 49 Studies have shown that treatment with selenite could reduce basal OS, 50 upregulate the expression of several cell cycle-related genes, 47 enhance gene expression and activity of GPx, reverse the decreased total antioxidant capacity, reduce GSH, and suppress reactive oxygen species production. 50 It is likely that the cellular mechanisms described in other cell types may also occur in aged ADSCs.
GSH-MEE did not change expression levels of the neural markers in NSCs and CNLCs generated of treated aged ADSCs. In this study, GSH-MEE was used because it is more lipophilic than GSH and is effectively transported inside many types of cells. 51 A previous study showed that GSH-MEE increased intracellular levels of GSH in rat mesencephalic cells in vitro. 52 Sagara and Makino showed that although the extracellular GSH could induce the neuronal differentiation in rat bone marrow stromal cells, the intracellular GSH had no effect in this respect. 53
Conclusion
Data presented in this study showed that sodium selenite and GSH-MEE increased the growth rate of aged rat ADSCs. Cell therapy requires a sufficient number of cells, so that these substances could potentially be used to prepare adequate cell numbers for autologous transplantation in aged people. In addition, sodium selenite increased expression of neural markers in NSCs and CNLCs generated of aged ADSCs, but GSH-MEE did not have any effects on aged ADSC differentiation capability. So, although these substances are both antioxidants, sodium selenite is more effective to improve performance of aged ADSCs.
Footnotes
Acknowledgment
The authors are grateful for the support of the Faculty of Medical Sciences, Tarbiat Modares University, Tehran, Iran.
The project was funded by Shefa Neuroscience Research Center, Khatam Alanbia Hospital, Tehran, Iran (Grant # 86-N-105).
Author Disclosure Statement
No competing financial interests exist.