
Abstract
Alzheimer's disease (AD) is a common form of dementia and amyloid-β peptide (Aβ) aggregation is considered to be one of its main causes. Paeoniflorin has been previously shown to attenuate cognitive damage inflicted by exogenous Aβ protein. Using transgenic Caenorhabditis elegans models expressing human Aβ1–42, we demonstrate here that paeoniflorin can delay progressive paralysis caused by endogenous Aβ expression and reduce the amount of toxic Aβ oligomers in vivo, although it has no effect on Aβ aggregation in vitro. Paeoniflorin does not, however, affect the lifespan of either wild-type or AD-like nematodes, implying a mechanism independent of a general antiaging effect. We then demonstrate that paeoniflorin can reduce reactive oxygen species levels in C. elegans AD models, which may contribute to its in vivo suppression of Aβ toxicity. Moreover, paeoniflorin is shown to upregulate the expression of the small heat shock protein HSP-16.2 as it is capable of increasing the hsp-16.2 transcript level in wild-type as well as AD-like nematodes and enhancing the fluorescence intensity in hsp-16.2::GFP nematodes. Taken together, our findings demonstrate the underlying mechanisms of the protective effect of paeoniflorin against age-onset Aβ proteotoxicity, which are, in part, connected with oxidative and heat shock stress responses.
Introduction
A
Although there are various types of Aβ aggregates, recent evidence suggests that soluble Aβ oligomers may be responsible for Aβ proteotoxicity, rather than the fibrillar aggregates found in plaques. 5 For example, oligomers such as the 56-kDa Aβ assembly that reacts with A11 antiserum are shown to cause cognitive impairment in mice. 6 The Caenorhabditis elegans strain CL4176, which is an AD model engineered to express human Aβ1–42 peptide in muscle cells, becomes paralyzed before the appearance of amyloid deposits, suggesting that damage to muscle is caused by soluble Aβ oligomers. 7 Moreover, pathological studies of AD brains have indicated significant oxidative stress attributed to the free radical-associated toxicity of soluble Aβ oligomers. 5 –7 Therefore, there is much interest among investigators in addressing the problem of toxic Aβ oligomeric intermediates. 8 –10
During the long history of the clinical practice of traditional Chinese medicine, many herbs have been used to treat neuronal diseases. The root of Paeonia lactiflora Pall. has been used for centuries in traditional Chinese herbal formulas such as Danggui Shaoyao San, prescribed for treatment of depression-like disorders. 11 Modern studies indicate that paeoniflorin is a major active component in this herb. For instance, paeoniflorin is shown to attenuate cognitive deficit in rats after a stereotactic intrahippocampal bilateral injection of exogenous Aβ. 12 Paeoniflorin is also reported to have an antioxidant activity, 11 –13 and can induce expression of heat shock proteins (HSPs). 14 Meanwhile, a number of signaling pathways of stress response, including oxidative stress and heat shock pathways, are known to be involved in the pathogenesis of AD. For instance, an elevated reactive oxygen species (ROS) level in SK-N-MC cells leads to an increased β-secretase gene/BACE1 expression, which may enhance Aβ production 15 ; and an increased expression of HSP-16.2, a small heat shock inducible chaperone protein, can alleviate proteotoxicity by reducing Aβ oligomers in transgenic C. elegans. 16 Therefore, it would be of interest to investigate whether paeoniflorin can provide protection against proteotoxicity caused by endogenous Aβ oligomers and how it interacts with stress response pathways.
In this study, we first used transgenic C. elegans models expressing human Aβ1–42 to examine the ability of paeoniflorin to alleviate proteotoxicity induced by Aβ aggregation and, in particular, assessed its effect on soluble Aβ oligomers. We then explored the underlying mechanisms of paeoniflorin with regard to key signaling components in aging and stress-related processes such as ROS and HSP levels.
Materials and Methods
Nematode strains and handling procedures
All C. elegans strains were obtained from the Caenorhabditis Genetics Center (University of Minnesota) and maintained on solid nematode growth medium (NGM) seeded with Escherichia coli OP50 as food. The wild-type strain N2 and the transgenic strains CL2006 {dvIs2 [pCL12 (unc-54/A-Beta1-42) + pRF4 (rol-6 (su1006))]} and CL2070 {dvIs70 [pCL25 (hsp-16.2p::GFP) + pRF4 (rol-6 (su1006))]} were grown at 20°C, while the transgenic strain CL4176 {dvIs27 [pAF29 (myo-3/A-Beta 1-42/let UTR) + pAF4 (rol-6 (su1006))]} was grown at 16°C. Synchronization of the nematodes was performed using the standard alkaline hypochlorite method unless stated otherwise.
Paralysis assay
C. elegans strain CL4176 was used for the paralysis assay as described. 16 Before the assay, nematode hermaphrodites were transferred to new NGM plates to lay eggs for 1 hour and the new generation of nematodes was grown to adulthood. The gravid adults were then transferred to 3.5 cm NGM plates (eight nematodes/plate, three plates for each treatment) containing paeoniflorin, which was prepared in H2O and brought to final concentrations as indicated with OP50 broth, and allowed to lay eggs for 1 to 2 hours at 16°C. The adults were removed and the plates each containing ∼100 newly laid eggs were incubated at 16°C. After 36 hours, the temperature was upshifted to 23°C to induce the transgenic protein expression in the nematodes (∼300 for each treatment) until the end of the paralysis assay. Nematodes were scored every 2 hours after they began to show signs of paralysis unless otherwise stated.
Toxic Aβ oligomer assay
The assay of toxic Aβ oligomers was performed using dot blot analysis as described. 17 After synchronization, eggs of C. elegans strain CL2006 were transferred to 7.5 cm NGM plates containing OP50 with or without paeoniflorin prepared as above. The plates (three for each group) were incubated at 16°C for 36 hours and then at 23°C for another 36 hours. After that, the nematodes were washed off the plates using the M9 buffer, resuspended in lysis buffer with protease inhibitor mixture (Beyotime, Shanghai, China), and homogenized. After centrifugation, the supernatant was precipitated overnight with methanol (1:4, v:v) at −20°C, and the proteins were resuspended in a loading buffer. Equal amount (2 μg) of proteins was spotted onto nitrocellulose membranes. After blocking with 5% skimmed milk, the membranes were incubated with rabbit anti-oligomer antibody (A11; Invitrogen, Carlsbad, CA; 1:1000) for 1 hour at room temperature. Peroxidase-conjugated goat anti-rabbit IgG (Earthox, San Francisco, CA; 1:5000) was used as secondary antibody. Densities of immunoreactive spots were analyzed using ImageJ Software (NIH, Bethesda, MD) as previously described. 17
Thioflavin T fluorescence assay
The content of toxic β-sheet amyloid was determined using thioflavin T (ThT) assay as described. 18 Aβ1–42 (Anaspec, Fremont, CA) was dissolved in 1,1,1,3,3,3-hexafluoro-2-propanol (1 mg/mL), sonicated in a water bath for 10 minutes, and stored at 4°C. Before use, fractions of Aβ were air-dried, dissolved in dimethylsulfoxide (1 mg/mL), and diluted to 40 μM in phosphate-buffered saline (PBS). Equal volumes of 40 μM Aβ solution and 1 mM paeoniflorin solution were mixed and incubated at 37°C. At indicated time points, 30 μL of the Aβ-paeoniflorin reaction was taken and mixed with 70 μL of ThT solution (final ThT concentration 5 μM). Fluorescence intensity was monitored at an excitation wavelength of 450 nm and an emission wavelength of 480 nm using Fluoroskan Ascent FL microplate reader (Thermo, Waltham, MA).
Lifespan analysis
Lifespan assay of N2 nematodes was conducted at 20°C in 96-well plates using liquid culture with or without 500 μM paeoniflorin as previously described. 19 The CL2006 nematodes always undergo paralysis, making it difficult to judge whether they are alive in liquid when they grow old. Therefore, lifespan of CL2006 nematodes was determined on solid NGM plates. Gravid CL2006 nematodes were allowed to lay eggs at 20°C on 7.5 cm NGM plates for 1 to 2 hours to produce an age-synchronized population. Once the population reached adulthood, 40 nematodes were transferred to fresh 3.5 cm NGM plates containing OP50 with or without paeoniflorin; three parallel plates were used for each treatment. The adult nematodes were transferred onto fresh plates every day during the egg-laying period, and the fraction of nematodes alive was scored every other day until all were dead.
Determination of ROS level
The ROS level in C. elegans models was determined using 2’,7’-dichlorofluorescin diacetate (DCFH-DA; Sigma, St. Louis, MO) essentially as described previously. 20 Briefly, the CL4176 nematodes (80 in each group) treated with or without paeoniflorin as above in “Paralysis assay” section were collected 36 hours after temperature upshift from 16°C to 23°C and homogenized on ice in 100 μL of PBS with 0.1% Tween-20. After adding 100 μL of 0.05% DCFH-DA, which can be directly oxidized to the fluorescent 2’,7’-dichlorofluorescein (DCF) by ROS, the samples were transferred into 96-well black plates and placed in the Fluoroskan Ascent FL microplate reader (Thermo) for 160 minutes with readings at 10-minute intervals (excitation, 485 nm; emission, 538 nm). Relative ROS levels were indicated by the DCF fluorescence intensity.
The ROS level was also analyzed in whole living nematodes as described previously. 21 Briefly, both CL4176 and N2 nematodes were treated as above and dispensed into 96-well black plates by a COPAS Biosort system (100 animals in 80 μL per well; 10 wells for each treatment). Then Tween-20 and DCFH-DA in PBS (120 μL) were added to each well to make final concentrations of 0.01% and 50 μM, respectively. DCF fluorescence intensity was monitored using the microplate reader every 30 minutes for 3 hours at 37°C. The data were obtained by subtracting the initial readings from the final readings and normalized as percentages to that of the control nematodes. To further examine the changes of ROS levels in the nematodes, fluorescent images were taken by a fluorescence microscope ( × 100) with a fluorescein isothiocyanate (FITC) excitation and emission filter setup using the nematodes from the plates after microplate reading.
Real-time PCR analysis
After adult nematodes were treated with alkaline hypochlorite, about 500 eggs were collected and transferred to NGM plates containing OP50 with or without 500 μM paeoniflorin. The plates were incubated at 16°C for 36 hours and then at 23°C for another 36 hours. Total RNA was extracted with TRIzol® reagent (Invitrogen). After reverse transcription, relative quantification of complementary DNA by real-time PCR was performed as described previously using SYBR Green. 19 Primers used are listed in Table 1.
Quantification of hsp-16.2::GFP expression
Fluorescence assay of hsp-16.2::GFP expression was performed using C. elegans CL2070 as described previously. 22 Synchronized L1 larvae were placed on solid NGM plates with or without 500 μM paeoniflorin and incubated at 20°C for 3 days. The nematodes were then harvested and immobilized with 20 mM sodium azide on 2% agarose pad, and HSP-16.2::GFP was quantified by fluorescence microscopy as described. 23 Briefly, the fluorescence images in the pharynx region, where GFP signal is most prominent, were taken by an Olympus IX51 inverted fluorescence microscope ( × 400). Fifteen animals were randomly selected in each group for quantifying the average GFP fluorescence intensity by ImageJ software (NIH, Bethesda, MD).
Statistical analysis
The effect of paeoniflorin on C. elegans was analyzed using an independent Student's t-test or a two-way analysis of variance test. A p-value of <0.05 was considered statistically significant.
Results
Paeoniflorin delays paralysis caused by Aβ expressed in muscle cells of CL4176 nematodes
The C. elegans strain CL4176 expresses human Aβ1–42 in muscle cells in a temperature-inducible manner.
24
Because Aβ peptides are toxic, the nematodes become paralyzed within 40 hours after the culture temperature being upshifted to 23°C from 16°C at the L3 larval stage. Using this transgenic model, we preliminarily tested the effect of paeoniflorin against Aβ-induced proteotoxicity at the concentration range of 100–1000 μM. The nematodes were treated with paeoniflorin at 16°C for 36 hours and then at 23°C, and the paralyzed nematodes were scored at designated time points corresponding approximately to 10–20%, 50–60%, and 80–90% paralysis, which were referred to as early (mild), middle (moderate), and late (severe) stage of paralysis, respectively (Fig. 1A). At 100 μM, paeoniflorin offered only limited protection against the toxicity at early and middle stages of paralysis, while at 200 and 500 μM, it was effective at all three stages to prevent paralysis of the nematodes. Noticeably, paeoniflorin showed almost no protective effect at the time points tested when the concentration was increased to 1000 μM. On the other hand, neither time of flight (equivalent to nematode length) nor extinction (EXT; an indicator of optical density and internal structure of nematodes) as determined by a COPAS Biosort system
25
was affected in CL4176 and N2 nematodes by paeoniflorin at <1000 μM, but EXT was slightly reduced at 1000 μM (Supplementary Fig. S1; Supplementary Data are available online at
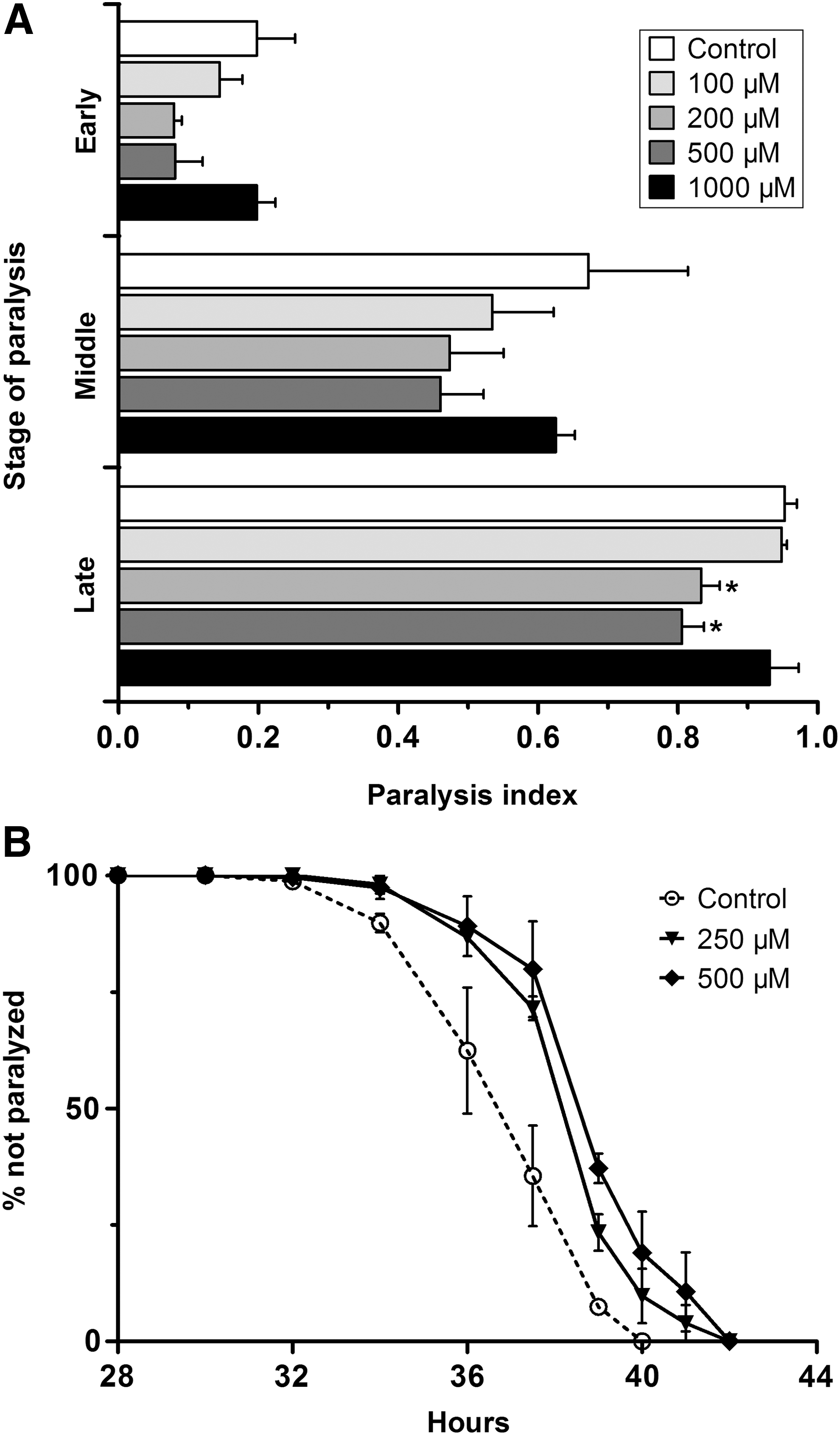
Effect of paeoniflorin on progressive paralysis in Aβ-transgenic C. elegans.
To further investigate the effect of paeoniflorin on the process of progressive paralysis, we then performed a temporal profiling of paralysis at 2-hour intervals from the beginning of paralysis until all nematodes were paralyzed using 250 and 500 μM paeoniflorin. As shown in Figure 1B, when treated with either 250 or 500 μM of paeoniflorin, the paralysis caused by Aβ expression was significantly deferred compared to that in the untreated CL4176 nematodes. A beneficial effect of paeoniflorin on paralysis was also observed in C. elegans strain CL2006 (data not shown), another nematode AD model that constitutively expresses human Aβ1–42 in muscle cells and shows an age-dependent paralysis phenotype. Together, these data demonstrate the protective activity of paeoniflorin in nematode AD models.
Paeoniflorin reduces soluble Aβ oligomer levels in CL2006 nematodes, but does not prevent in vitro Aβ oligomer formation
Soluble Aβ oligomers are thought to be responsible for the toxicity of Aβ aggregates in many animal models. To determine whether inhibition of Aβ oligomerization by paeoniflorin underpins its beneficial effect in animals, we used the nematode CL2006, an Aβ model of slow paralysis with accumulations of both oligomers and fibrils, 8 to test the level of toxic Aβ oligomers (A11 antiserum reactive) by dot blotting. To be consistent with the above experiments, the nematodes were incubated with 500 μM of paeoniflorin at 16°C for 36 hours and then at 23°C for another 36 hours. As shown in Figure 2A, the ratio of Aβ oligomers was clearly reduced, demonstrating a decreased level of toxic oligomeric Aβ form in the nematodes. In addition, the fibrillar Aβ deposits were also decreased in CL2006 nematodes after paeoniflorin treatment as observed by ThT staining (data not shown), suggesting that paeoniflorin was able to reduce the levels of both oligomeric and fibrillar Aβ forms.

Effect of paeoniflorin on Aβ aggregation.
In some cases, A11-positive β-sheet-rich toxic oligomers can be remodeled by small compounds such as epigallocatechin-3-gallate (EGCG), which thereby reduce oligomer levels. 26 We therefore asked whether paeoniflorin can remodel Aβ oligomers in vitro by using an assay involving fluorescent ThT, which is an indicator dye widely used to visualize and quantify the presence of amyloid fiber formation. 27 As shown in Figure 2B, however, the ThT fluorescence intensities of aggregating Aβ peptides were the same whether or not paeoniflorin was present, suggesting that the in vitro aggregation process of Aβ was unaffected by paeoniflorin.
Paeoniflorin does not affect the lifespan of either N2 or CL2006 nematodes
Aging plays an important role in the process of late-onset neurodegeneration, and the paralysis phenotype is also age related in transgenic Aβ nematodes such as CL4176 and CL2006. Previously, we showed that aging was involved in the counter-proteotoxicity effect of some compounds such as astragalan, a polysaccharide isolated from Astragalus membranaceus. 19 Therefore, we determined the lifespan of both wild-type (N2) and Aβ (CL2006) nematodes to investigate whether the effect of paeoniflorin against Aβ-induced toxicity could be part of an antiaging effect. As shown in Figure 3, however, the lifespans of both N2 and CL2006 strains were not affected by treatment with 500 μM of paeoniflorin, suggesting that the protective activity of paeoniflorin is independent of a general antiaging effect.
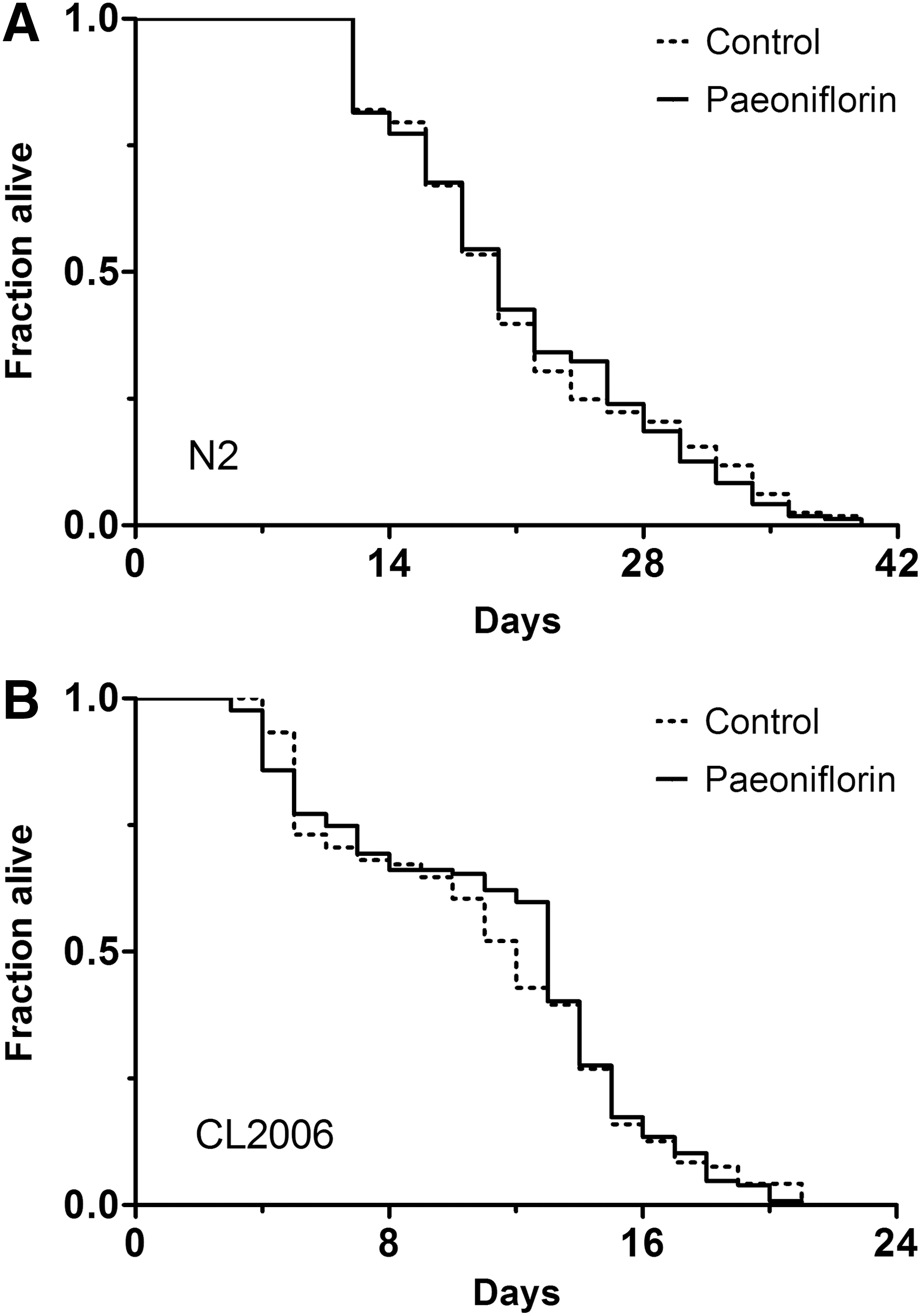
Effect of paeoniflorin on the lifespan of wild-type and Aβ-transgenic C. elegans. The adult lifespan was determined for N2
Paeoniflorin reduces ROS levels in CL4176 nematodes
It is known that Aβ peptides in the form of soluble oligomers can enhance free radical formation, which causes oxidative stress and results in cell death. 10 Since paeoniflorin can reduce the ratio of toxic Aβ oligomers in vivo as demonstrated above, we went on to investigate its antioxidant action using the transgenic Aβ nematode CL4176. The nematodes were treated with 500 μM of paeoniflorin at 16°C for 36 hours and then at 23°C for another 36 hours before determination of ROS level using the DCFH-DA methods. As shown in Figure 4A, the DCF fluorescence intensity was persistently lower in the homogenates of nematodes treated with paeoniflorin compared to that of the control. DCF fluorescence intensity was also shown to be significantly reduced in whole animals treated with paeoniflorin as analyzed by direct microplate reading (Fig. 4B) and by fluorescence microscopy (Fig. 4C) of the CL4176 nematodes compared to the untreated controls. These data suggest that paeoniflorin can reduce ROS level in the Aβ nematode model CL4176 after temperature upshift. Interestingly, the DCF fluorescence intensity was also lower in N2 nematodes after paeoniflorin treatment (Fig. 4B, C), demonstrating that the ROS level of wild-type nematodes was reduced by paeoniflorin. Taken together, these findings suggest that the ROS-reducing capability of paeoniflorin may contribute to its protection against Aβ proteotoxicity.
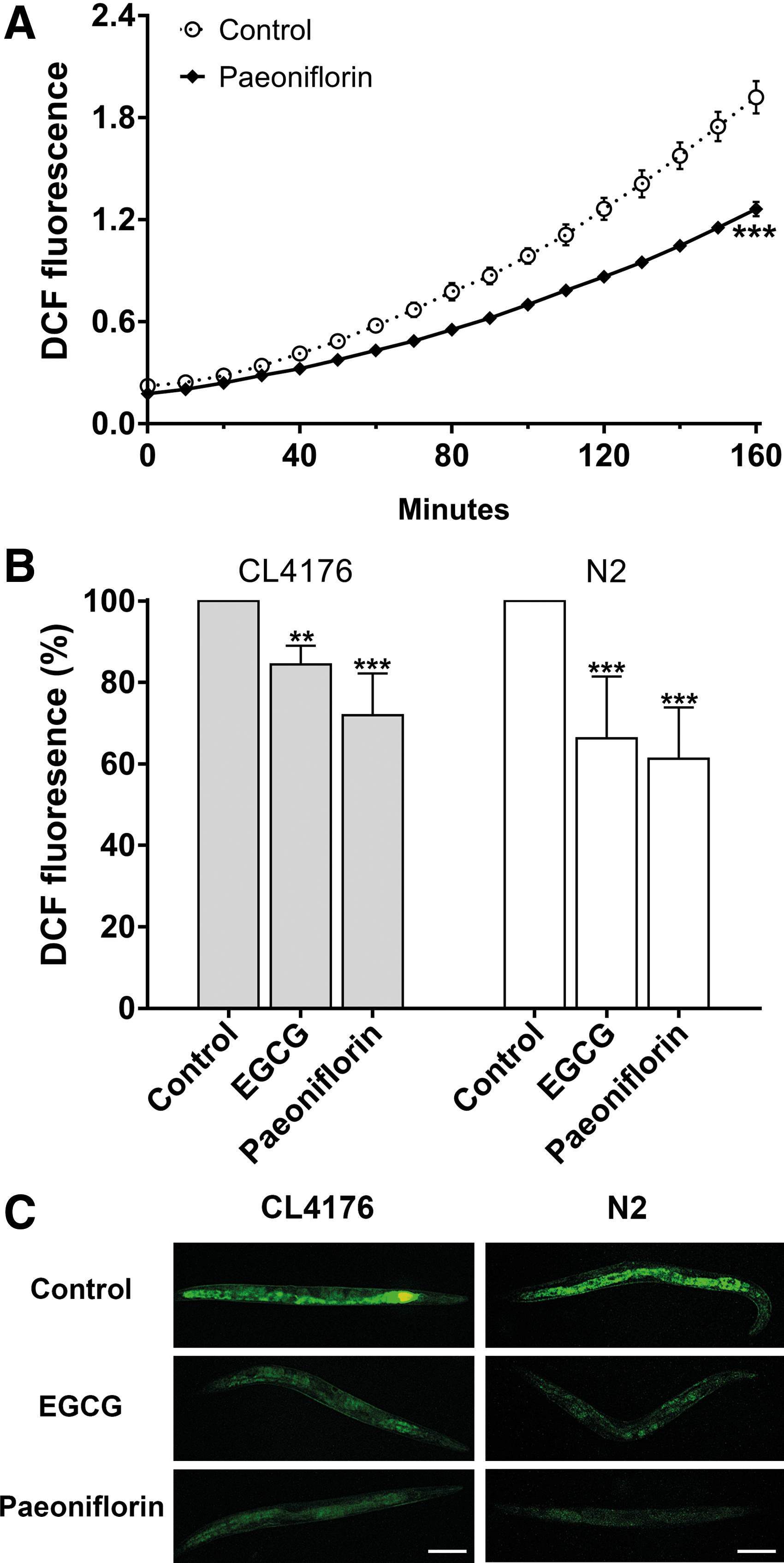
Effect of paeoniflorin on ROS levels in C. elegans. The nematodes were treated with or without 500 μM of paeoniflorin and then collected for ROS analysis.
Paeoniflorin induces the expression of hsp-16.2
HSPs are important in the clearance of unfolded and misfolded proteins in C. elegans, 28,29 and paeoniflorin is known to induce HSP in human neuroblast cells. 14 We therefore hypothesized that the ability of paeoniflorin to reduce toxic Aβ oligomers in the nematodes might be related to its ability to induce HSPs. To test this, we determined the gene expression levels of heat shock factor 1 (HSF-1) and two representative HSPs (HSP-16 and HSP-70) using quantitative real-time PCR analysis. The nematodes were treated with or without 500 μM paeoniflorin at 16°C for 36 hours and at 23°C for another 36 hours, and then used for the subsequent real-time PCR assay. As shown in Figure 5, expression of hsp-16.2 (HSP-16) was upregulated in both N2 and CL4176 nematodes, while expression of hsf-1 and hsp-1 (HSP-70) was not affected. To further confirm the upregulation of hsp-16.2 by paeoniflorin, we then used the transgenic C. elegans strain CL2070, which carries GFP as a reporter to facilitate visualization of hsp-16.2 expression, 23 to investigate the effect of paeoniflorin on HSP-16 expression in living animals. The CL2070 nematodes were treated with 500 μM paeoniflorin from L1 for 3 days at 20°C and then subjected to analysis of HSP-16.2::GFP by fluorescence microscopy. As shown in Figure 6, the relative GFP fluorescence in paeoniflorin-treated nematodes was higher compared with the control, demonstrating that paeoniflorin was indeed able to upregulate the expression of hsp-16.2 in the nematodes.
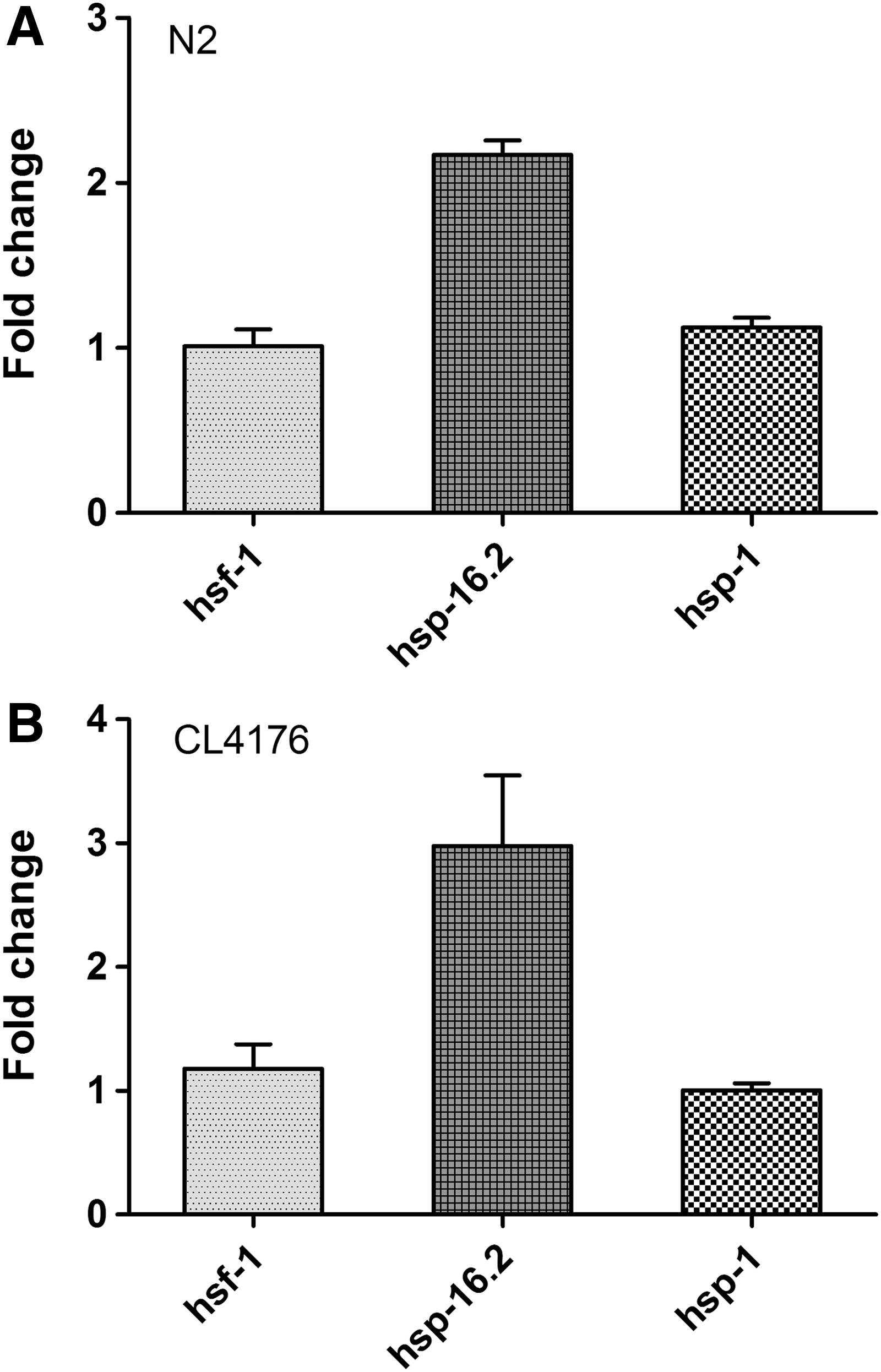
Effect of paeoniflorin on the expression levels of HSPs in C. elegans. The expression levels of hsf-1, hsp-16.2, and hsp-1 were determined by quantitative real-time PCR assay in N2
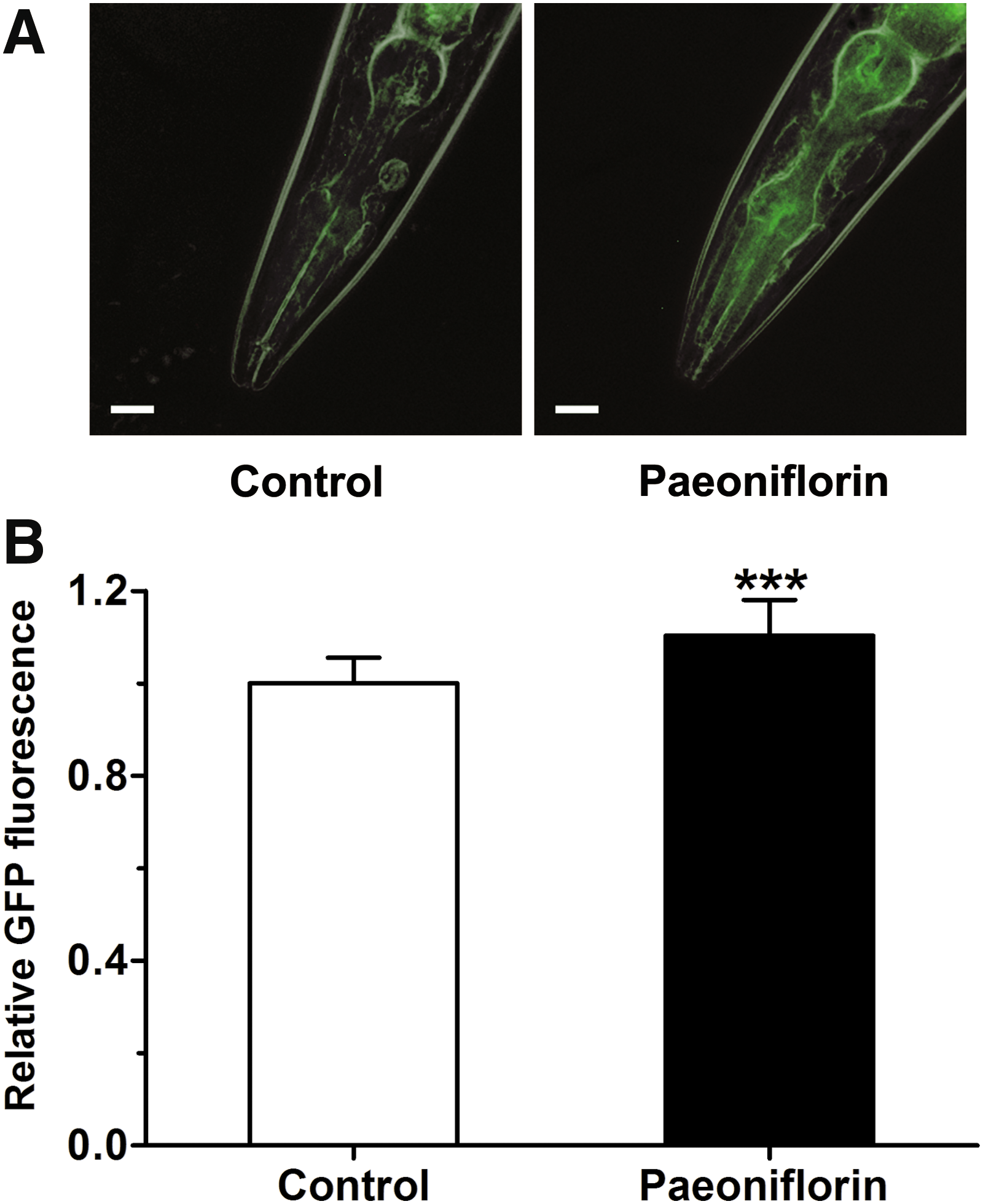
Effect of paeoniflorin on hsp-16.2 expression in C. elegans CL2070. The nematodes were treated with or without 500 μM of paeoniflorin and subjected to visualization of HSP-16.2::GFP by fluorescence microscopy.
Discussion
Although there is still some debate, Aβ aggregation is thought to be the main etiological factor in AD. 30,31 Aβ oligomers provide a potential explanation for the toxicity of Aβ peptides and link the deposition of Aβ plaques with neuronal loss. 2,10 Consequently, there is considerable interest in finding candidate drugs that may reduce levels of toxic Aβ oligomeric intermediates. 25,32 Natural extracts derived from medicinal plants, such as EGb 761 from ginkgo leaves, 20 or traditional Chinese medicines, such as Liuwei Dihuang, 33 are known to protect against Aβ toxicity in C. elegans AD models. We report here that paeoniflorin, a natural compound from the root of P. lactiflora, also has a beneficial effect in the nematode AD model CL4176, at a comparable level with the known antiparalysis compounds curcumin 24 and resveratrol 34 (Supplementary Fig. S2). Injury of cerebral vascular smooth muscle cells by Aβ aggregation is one of the characteristics of AD and the related condition, vascular dementia, 1,35,36 and therefore paeoniflorin may have potentials in the prevention and treatment of these forms of dementia. Further studies using mammalian models and human subjects may provide important insights into its therapeutic applications.
Because it is hypothesized that Aβ oligomers, and not Aβ fibrils, are responsible for neurotoxicity in AD and related disorders, 5 many studies have attempted to uncover the process by which Aβ monomers aggregate into toxic oligomers and find ways to reduce oligomer concentration. Molecules that target and neutralize specific toxic aggregated conformers have been reported. For example, resveratrol, a polyphenol extracted from the skin of red grapes, was found to be able to selectively remodel three types of Aβ conformers (soluble oligomers, fibrillar intermediates, and amyloid fibrils) into an alternative aggregated, but nontoxic, species. 37 In C. elegans AD model CL2006, RNA interference of the daf-2 gene also reduced toxic Aβ oligomers: induced HSF-1 was associated with the disaggregation of toxic Aβ oligomers, while active DAF-16 was associated with the process whereby oligomers are converted into less toxic aggregates. 38 We show in this report that paeoniflorin also reduced the level of toxic Aβ oligomers (i.e., A11-reactive species) in C. elegans strain CL2006, although it had no effect on the aggregation of Aβ monomers in vitro, and this therefore represents a potential mechanism for its protective activity in nematodes.
AD is an age-related neurodegenerative disease and toxic forms of Aβ peptides increase with aging, and therefore lifespan extension may delay Aβ proteotoxicity. 38 Although paeoniflorin was able to protect nematode models from Aβ toxicity, it did not, however, affect the lifespan of either AD model or wild-type nematodes, indicating a mechanism not directly related to the aging process per se. On the other hand, aggregated Aβ oligomers are known to be associated with high oxidative stress levels in C. elegans AD models, 7,39 which is consistent with the inflammation and high oxidative stress characteristic of the brain tissue of AD patients. 9,10 Also, Aβ-initiated free radicals react rapidly with several components of the plasma membrane, causing membrane protein oxidation and lipid peroxidation. Such oxidized species may migrate to different parts of the neuron and alter cellular function, resulting in cell death. 8 In paeoniflorin-treated nematodes, we found a significant decrease in ROS levels, which may relate to the reduction in toxic A11-reactive oligomers.
An important element of protein homeostasis involves highly conserved stress responses that monitor the cellular environment and prevent protein mismanagement. 38,40 Partly, this is achieved by the heat shock response, which leads to the upregulation of a series of HSPs. In nematode AD models, hsp-16.2 encodes HSP-16, which interacts directly with Aβ peptide and alters its oligomerization pathways, leading to reduced formation of toxic species. 29 Consistent with this, we found that hsp-16.2 expression levels were upregulated by paeoniflorin in both N2 and CL4176 nematodes, suggesting an important role for this HSP in the protective effect of paeoniflorin. Additional experiments using techniques such as RNA interference would further reveal the involvement of hsp-16.2/hsf-1 and other pathways in the effect of paeoniflorin against Aβ-induced paralysis in nematode AD models.
Footnotes
Acknowledgments
This work was supported by the Special Funds of the Central Finance to Support the Development of Local Universities and Colleges, Guangdong Province Department of Education (Grant Nos. 2015KGJHZ022 and 2016KQNCX084), the National High-Tech R & D Program of China (863 Program; Grant No. 2014AA022001), and the Basic Research Project (Grant No. 2015JB007) from the Institute of Forensic Science.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.