
Abstract
Radix paeoniae alba (RPA) and Veratrum nigrum (VN) L. belong to the 18 incompatible medicaments and have been prohibited for thousands of years in China. Previous studies focused on the chemical constituents that induced the toxicological response of the two agents, but this study offers preliminary insight into the pharmacodynamics and mechanism on estrogenic activity, which is responsible for their incompatibility. We undertook a characterization of the interaction on estrogenic activity of RPA and VN using in vivo models of immature and ovariectomized (OVX) mice and in vitro studies focused on estrogen receptor (ER) pathway for further mechanism. VN disturbed the estrogenic efficacy of RPA in promoting development of uterus and vagina in immature mice, and reversing the atrophy of reproductive tissues in OVX mice by decreasing the increase of serum estrogen level and upregulation of ER expression in reproductive tissues by treatment with RPA. Besides, VN antagonized the estrogenic efficacy of RPA in stimulating the binding with ERα and ERβ, increasing ERα/β-estrogen response element (ERE) luciferase reporter gene expression and promoting MCF-7 cell viability. This study provided evidence that VN antagonized the estrogenic efficacy of RPA by decreasing the up-regulations of estrogen biosynthesis in circulation and ERs in target tissues caused by RPA, and through ER-ERE-dependent pathway.
Introduction
Traditional Chinese medicine (TCM) makes significant contributions to the health of Chinese and has become more popular worldwide because of its efficacy and curative effects. Herb-herb combinations have been used in Chinese medicine practice for thousands of years, the traditional theory of “The eighteen incompatible medicaments”—some controversial herb combinations—were strongly prohibited because the interactions may lead to toxicity and unexpected consequences by either strengthening some side effects or diminishing some therapeutic effects and so on. 1 However, the incompatibilities are not absolutely because there are also some records to use some herb combinations listed in the 18 incompatible medicaments to treat serious diseases successfully in clinic. 2 Currently, researchers primarily focus on the chemical constituents that induced the toxicological response. However, the studies on the pharmacodynamics effect, condition (the specific effect of herb, dose range, and physiological and pathological state of body), and mechanism of such incompatibility are less.
Radix paeoniae alba (RPA) and Veratrum nigrum (VN) are 2 herbs belonging to the 18 incompatible medicaments, whose concurrent use has been prohibited. Commonly, VN is used to treat hypertension, stroke, excessive phlegm, and epilepsy, 3 despite its highly poisonous perennial herb native to Asia and Europe. 4 Studies have revealed that VN can decrease blood pressure and heart rate in hypertensive rats, 5 and it affords significant protection against hepatic ischemia/reperfusion injury in rats. 6
RPA is a traditional medicine in oriental countries because of its functions of nourishing blood, protecting liver, and alleviating pain. It has been widely used for over 2000 years in clinical practice for the prevention of cardiac diseases, arthritis, and other inflammation-related disorders based on its pharmacological effects in multiple tissues, 7 –10 particularly in gynecological diseases, such as abnormal menstruation, menstrual pain, and breast tenderness. 11 It was reported that RPA has estrogenic activity, which was indicated by the increase in the immature mice uterus weight and RPA pharmacological serum-inducing estrogen receptor (ER)-positive cell MCF-7 cell proliferation. 12,13
It was reported that the increased toxicity of VN and RPA combination was mainly due to the content increase of the special veratrum alkaloids, and the cytotoxicity of veratridine in SH-SY5Y cells was decreased with the increase in paeoniflorin concentrations. 14 Although we confirmed that RPA exerted the estrogenic effect on immature mice or ovariectomized (OVX) mice 13 and VN was similar to ER antagonist ICI182780, 15 the conditions and mechanism of incompatibility of the combination of VN and RPA by affecting the estrogenic effects are still kept untouched. So in this study, a further research was made on estrogenic effects of the combination use of RPA and VN by using in vivo models of immature and OVX mice along with in vitro studies to investigate its mechanism by ER-estrogen response element (ER-ERE) pathway. This study is part of an ongoing effort to provide insight into the nature of VN and RPA incompatibility.
Materials and Methods
Preparation of RPA and VN
RPA and VN were purchased from Beijing Huamiao Medicinal Herbs Co. Ltd. (Anhui, China) and Anhui Tongling Medicinal Herbs Co. Ltd. (Anhui, China), respectively. RPA was cut into small pieces and boiled twice with distilled water for 1 hour under reflux. The aqueous extracts were collected and filtered. The filtrates were then concentrated under reduced pressure at 60°C to a concentration of 100 and 200 g/L. VN was extracted by the same method and to a concentration of 2.25 g/L.
The samples of RPA and VN mixture (RPA:VN = 1:1) for cell culture assays were prepared by extracting the small pieces with distilled water (thrice, for 1 hour each). The extracts were concentrated in vacuo and dissolved in dimethyl sulfoxide (DMSO) (0.001 g/L).
In vivo studies
Animals and experimental design
The experimental protocol was approved by Institute of Chinese Materia Medica, China Academy of Chinese Medical Sciences, and all methods were carried out in accordance with the approved guidelines.
Immature mice model, 21-day-old female Kunming mice (12 ± 2 g) were purchased from Experimental Animal Center of Academy of Military Medical Sciences (Certificate No. SCXK [Jing] 2012-0004). The immature mice were randomly assigned to 10 groups: control group (Con), estradiol (E2, 0.1g/kg), E2 plus estrogen receptor antagonist (ICI, 0.005g/kg), E2 plus VN (VN, 0.045g/kg), RPA (RPA 2, 4 g/kg), RPA plus ICI, RPA plus VN for consecutive 7d. ICI182780 group was given intraperitoneal injection, untreated control mice received distilled water only, and for the rest of the groups, all test samples were orally administrated once a day for 7 days.
OVX mice model, 4-week-old female Kunming mice maintained normal 5-day estrous cycles, as confirmed by daily vaginal epithelium cell smear testing until ovariectomy was performed. Dorsal ovariectomy was performed under general anesthesia using 0.3 mg/kg of chloral hydrate. All OVX mice were checked by daily vaginal epithelium cell smear analysis, in which five consecutive days of leukocytes were indicative of constant diestrus and successful ovariectomy. In sham-operated negative controls, fat near the ovary was removed. The mice were randomly assigned to 11 groups: sham operated (Sham), OVX without treatment, and the rest of the 9 groups were the same with the immature mice, with 10 mice in each group. The same administration as immature mice lasted for 4 weeks.
Dose calculations followed guidelines correlating the dose equivalents between humans and laboratory animals based on ratios of body surface area. All animals were maintained on a 12-hour light/12-hour dark cycle under constant temperature (24°C ± 2°C) and humidity (55% ± 5%), and allowed free access to food and water.
Analysis of target tissue and serum
Blood was collected by removing the eyeball for analysis of E2, follicle-stimulating hormone (FSH), and luteinizing hormone (LH) levels was performed by enzyme-linked immunosorbent assay (ELISA) (Beijing Xinfangcheng Biotechnology, China). 13,15,16 The sensitivities of the three ELISA assays were 1.0 pg/mL, 1.0 m IU/mL, and 1.0 ng/mL, respectively, and not soluble structural analogues with other cross-reactions, and all the intra-assay and inter-assay variation of each hormonal assay were less than 9% and 15%. All animals were sacrificed by decapitation after collecting blood. The uterus of immature mice and OVX mice and adrenal gland of OVX mice were removed and weighed. The left horns of the uterus and the upper portion of the vagina were stored at −80°C for analysis by Western blot and real-time quantitative polymerase chain reaction (RT-PCR). The right horns of the uterus and the rest of the vagina were fixed with 4% polyoxymethylene for 24 hours for staining with hematoxylin and eosin (H&E) and immunohistochemistry. The right samples were embedded in paraffin and prepared for cross-sections; 4-μm-thick sections were cut, mounted, and stained with H&E for microscopy. 17
Immunohistochemistry
Tissue sections, 4-μm thick, of uterus and vagina were mounted on polylysine-coated sides. The paraffin sections were dewaxed by a routine method and incubated for 10 minutes with 3% hydrogen peroxide (H2O2). Each section was incubated with blocking serum (Boster Biological Technology Co., Ltd.) at room temperature for 30 minutes and then with primary rabbit anti-ER-α antibody (dilution 1/20; Santa Cruz Biotechnology) and a rabbit anti-ER-β (dilution 1/30; Santa Cruz Biotechnology), respectively, overnight at 4°C. Sections incubated in phosphate-buffered saline without antibody served as negative controls. After incubation with biotinylated secondary antibody, sections were incubated with an avidin-biotin complex reagent containing horseradish peroxidase for 30 minutes. The sections were then stained with 3, 3′-diaminobenzidine (Boster Biological Technology Co., Ltd.). 18 The Image-Pro plus 6.0 System image analysis system was used for quantitative analysis.
Western blot
Uterus and vagina were resuspended in lysis buffer (50 mM Tris, pH 8.0, 150 mM NaCl, 5 mM EDTA, 0.1% sodium dodecyl sulfate, and 0.5% NP-40) containing 10 Mm phenylmethylsulfonyl fluoride and 2 mg/mL aprotinin. The protein was obtained to detect the levels of ERα and ERβ in target tissue by Western blot. 19 The antibody of rabbit anti-ERα polyclonal antibody (dilution 1/200; Santa Cruz Biotechnology) or mouse anti-ERβ monoclonal antibody (dilution 1/1000; Abcam Biotechnology) was used. All experiments were done in triplicate. The relative quantity of each antibody was measured by Alpha Ease FC (Fluorchem FC2) software. The density ratio of protein to glyceraldehyde 3-phosphate dehydrogenase (dilution 1/1000; Cell Signaling Technology) was calculated from the band density.
Real-time quantitative polymerase chain reaction
The RT-PCR protocol and quantitative analysis were carried out following the protocol of our previous study. 16 Gene-specific primers were used for ERα (forward: CGCCTTCTACAGGTC TAAT; reverse: GGTTCTTGTCAATGGTGC), ERβ (forward: CT GTGAGGTAGGAATGCGAA AC; reverse: GGTCTGGGTGATTGCGAAGA), and β-action (forward: CCTCTATGCCAACA CAGTGC; reverse: CTGTAGAACGGTGTGGTC ATC). The mRNA levels of ERα and ERβ were normalized to the β-action mRNA level. PCR was performed as 95°C for 10 min, followed by 40 cycles of 95°C for 30 s and 60°C for 1 min. The quantification data were analyzed with ABI Prism analysis software. The relative mRNA expression was calculated with the comparative Ct method. 20
In vitro studies
MTT assay of MCF-7 cell viability
The MCF-7 cell line was purchased from Xiehe Cell Research Institute of Peking Union Medical College (from the American Type Culture Collection). Three days before testing, the cells in logarithmic phase were digested by trypsin enzyme (0.05%) and cultured in Dulbecco's Modified Eagle Medium containing 1% sodium pyruvate (vol/vol), 1% nonessential amino acids (NEAA) (vol/vol), 1% GlutaMAX-1 (vol/vol) (Invitrogen, Carlsbad, CA), and 10% heat-inactivated FBS (vol/vol) without phenol red for purpose of depleting the intracellular estrogen. Then, they were inoculated to the 96-well plate at 180 μL per well. After 24 hours, test samples of 20 μL RPA extract (0.1, 1, 10, 100 μg/mL), 17β-estradiol (E2 0.01 μM), RPA or RPA as well as the 0.1 μM/L ICI were also added into the plate, test samples with E2 + VN (10-9 μM + 1 μg/mL), RPA and VN mixture (at the ratio of 1:1) at the above different concentrations, and 0.1% DMSO solvent blank (the same final concentration of DMSO in test sample solutions) were added and incubated at 37°C and 48 h. Proliferation was determined by the MTT (3-[4,5-dim ethylthiazol-2-yl]-2,5-diphenyltetrazolium) bromide assay at 490 nm. Percent growth induction was calculated as a percentage of the average response of the DMSO control samples. Results reported are the mean ± standard deviation (SD) of four replicate determinations from a representative assay. 18
Transfection and reporter assay of ER-subtype selectivity
HEK 293 cells were stably transfected with human estrogen receptor α/β (hER α/β) and the ERE plasmid (kindly provided by Professor Yung-Chi Cheng, Yale University), and the luciferase reporter assay system from Promega (WI) was used to evaluate the formation of functional ER α/β -ERE complexes. The cells were maintained and primed to minimize the effects of endogenous estrogens as described above and then seeded (1 × 105 cells/100 μL/well) in 96-well plates. The same test samples with the MTT assay of MCF-7 cell proliferation were added to three replicate wells, and were incubated for 24 h. The luciferase activity was measured as described in our previous reports. 21 Activity of the luciferase reporter gene was expressed relative to the DMSO control. Results reported are the mean ± SD of three replicate determinations from a representative assay.
ERs competitive ligand-binding assay
To confirm if RPA could bind to ERs, LanthaScreen Time Resolved-Fluorescence Resonance Energy Transfer (TR-FRET) ERα and ERβ competitive binding assay kits were used (Life Cat NO:A15883 and A15890). Briefly, serial dilutions of RPA and VN mixture (0.004, 0.013, 0.04, 0.12, 0.37, 1.1, 3.3, and 10 μg/mL) were combined with Fluormone™ GS1 Green for binding with terbium-labeled ER-ligand binding domains (LBD) on a 384-well plate. One hour later, after incubation at room temperature, the fluorescence intensity was detected on a microplate reader (Excitation: 340 nm; Fluorescein emission: 535 nm; Terbium emission: 485 nm; EnvisionTM, PerkinElmer). The final data were shown by normalizing the signal of fluorescein to that of terbium.
Statistics Analysis
The software SPSS software version 11.0 for Windows (SPSS, Inc, Chicago, IL) was used for statistical analysis. All data are expressed as mean ± SD and were analyzed by one-way analysis of variance followed by least significant difference or Dunnett's T3 test. Differences were considered statistically significant when p was less than 0.05.
Results
Effect of incompatibility of VN and RPA on body, uterine, and adrenal gland weight
RPA or E2 had significant stimulatory effects on the uterine weights of immature and OVX mice (p < 0.05, 0.01, or 0.001) as shown in Figure 1A, B. Compared with RPA or E2 treatment alone, RPA + ICI182780 or RPA + VN induced a lower uterus index in immature and OVX mice (p < 0.05, 0.01 or 0.001). The results suggest that VN could inhibit the estrogenic efficacy of RPA on increasing the uterine weight of immature and OVX mice.
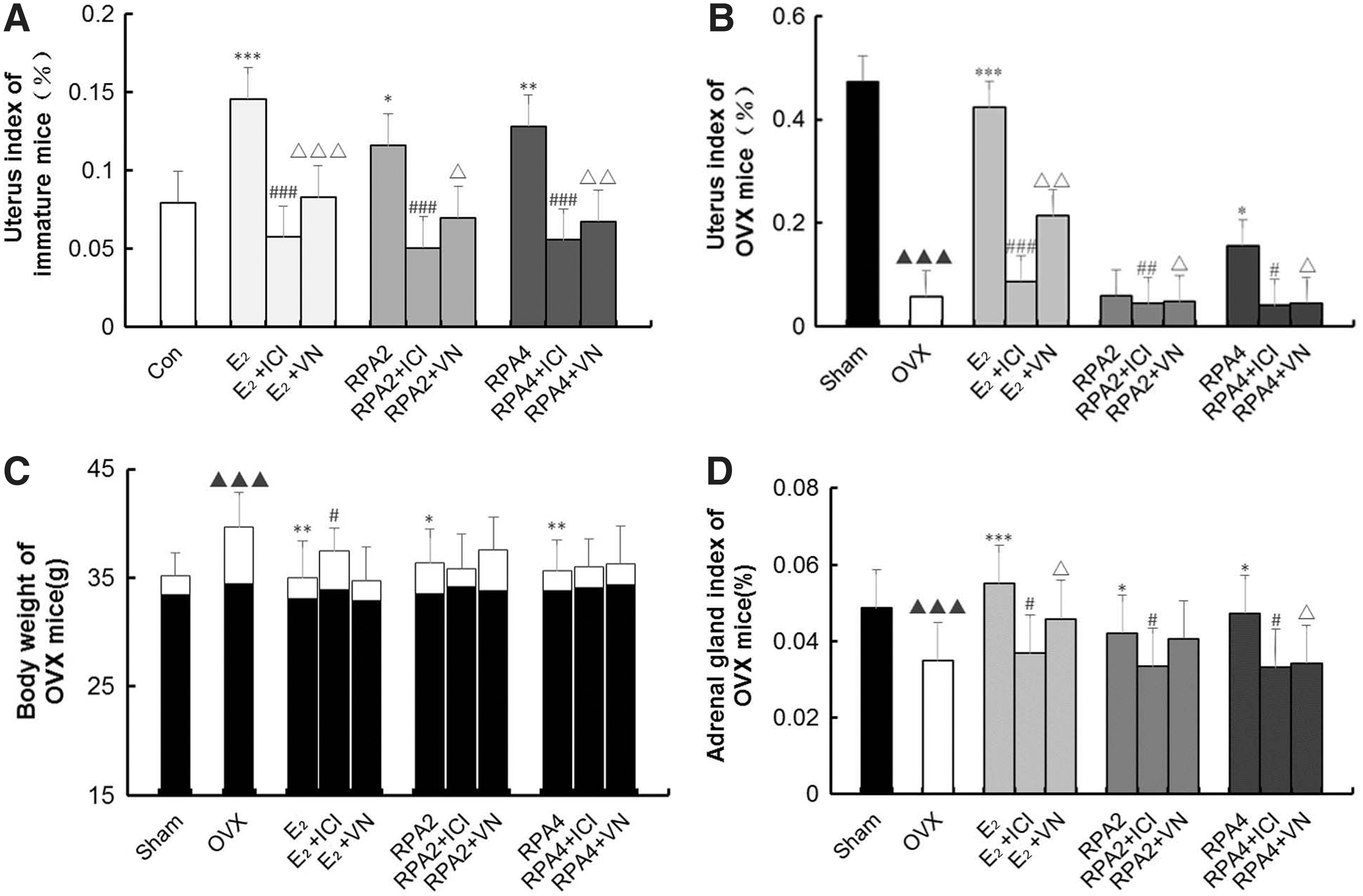
The effects of incompatibility of VN and RPA on uterine, adrenal gland, and body weights.
Figure 1C showed that treatment with RPA or E2 completely prevented the increase in body weight of OVX mice associated with E2 deficiency. The estrogen antagonist ICI182780 and VN could lower the estrogenic efficacy of E2 or RPA on preventing the increase in body weight of OVX mice, but VN combination treatment-induced differences were not significant.
Adrenal gland weight showed similar results with uterine weight. ICI182780 and VN could attenuate the increases in adrenal gland weight induced by RPA or E2 treatment (p < 0.05) (Fig. 1D).
Effect of incompatibility of VN and RPA on levels of serum E2, LH, and FSH
Figure 2 showed that E2 or RPA treatment significantly raised levels of circulating serum E2 compared to those of untreated immature and OVX mice (p < 0.05, 0.01 or 0.001). Meanwhile, RPA treatment significantly downregulated LH and FSH content in immature and OVX mice as E2 treatment group (p < 0.05, 0.01 or 0.001). VN + RPA groups significantly attenuated the increase of serum E2 and the decrease of LH and FSH in immature and OVX mice compared with RPA groups (p < 0.05, 0.01, or 0.001). This effect was comparable to that of the ER antagonist ICI182780 combination treatment (p < 0.05, 0.01 or 0.001).
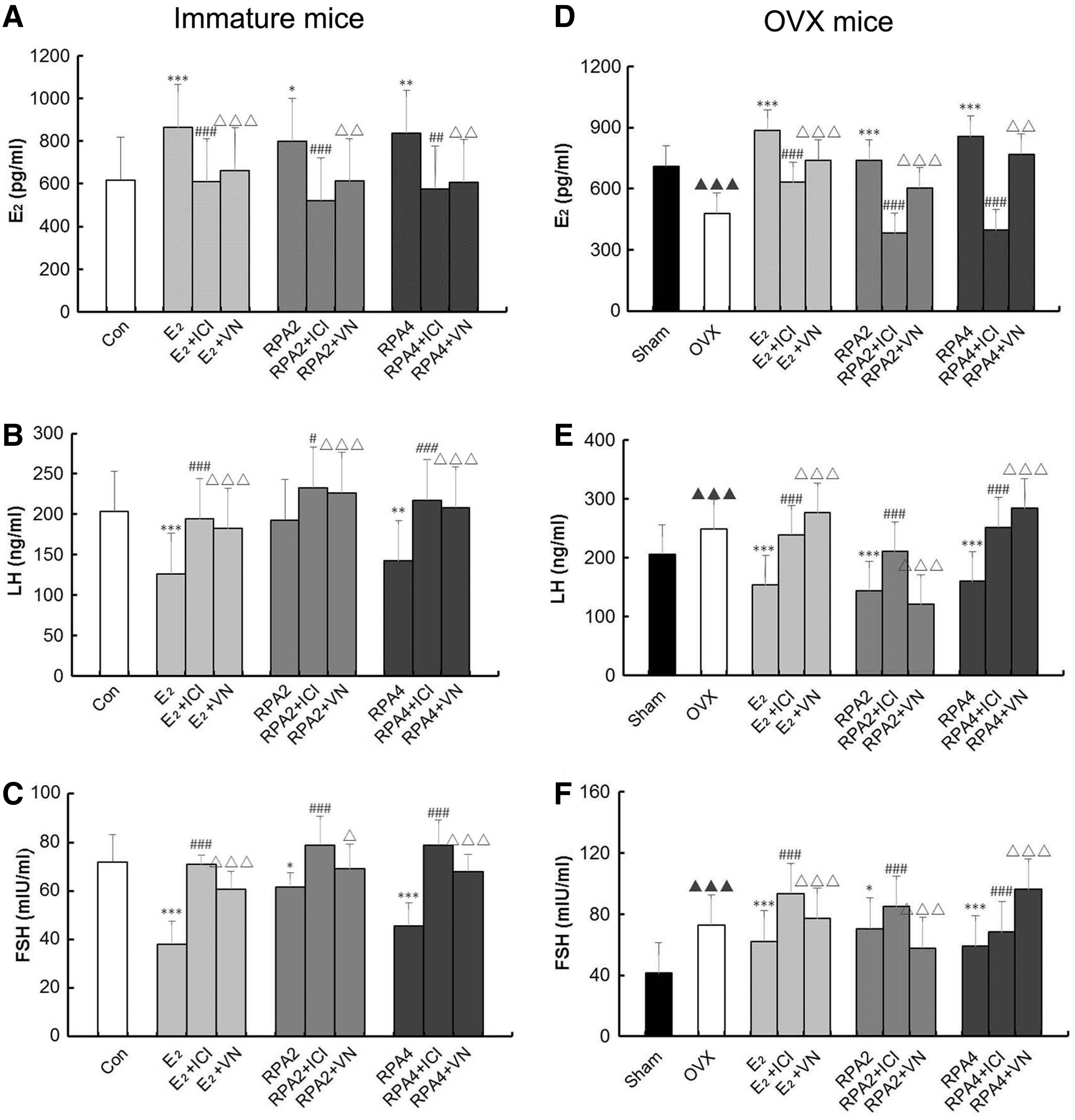
The effects of incompatibility of VN and RPA on serum estradiol (E2), LH, and FSH in immature and OVX mice. Serum levels of E2
Effect of incompatibility of VN and RPA on histology of the uterus and vagina
Compared with untreated controls, histological analysis of uterine sections revealed that treatment with E2 or RPA substantially induced the growth and development of the uterus in immature mice and restored the atrophy of the uterus in OVX mice. These results in treated uterus samples were indicated by the thickening of the uterine endometrium, an increased number of glands, and more extended glandular cavities compared with untreated samples. In RPA + VN or ICI182780 groups, the uterine morphology was atrophic, as indicated by the thinner uterine endometrium, the decreased number of glands, and fewer extended glandular cavities compared with samples treated with RPA-alone treatment. The results depicted that VN reduced the estrogenic efficacy of RPA in promoting the growth and development of the uterus in immature mice and restoring the atrophy of the uterus in OVX mice (Fig. 3A, C and Tables 1 and 2).
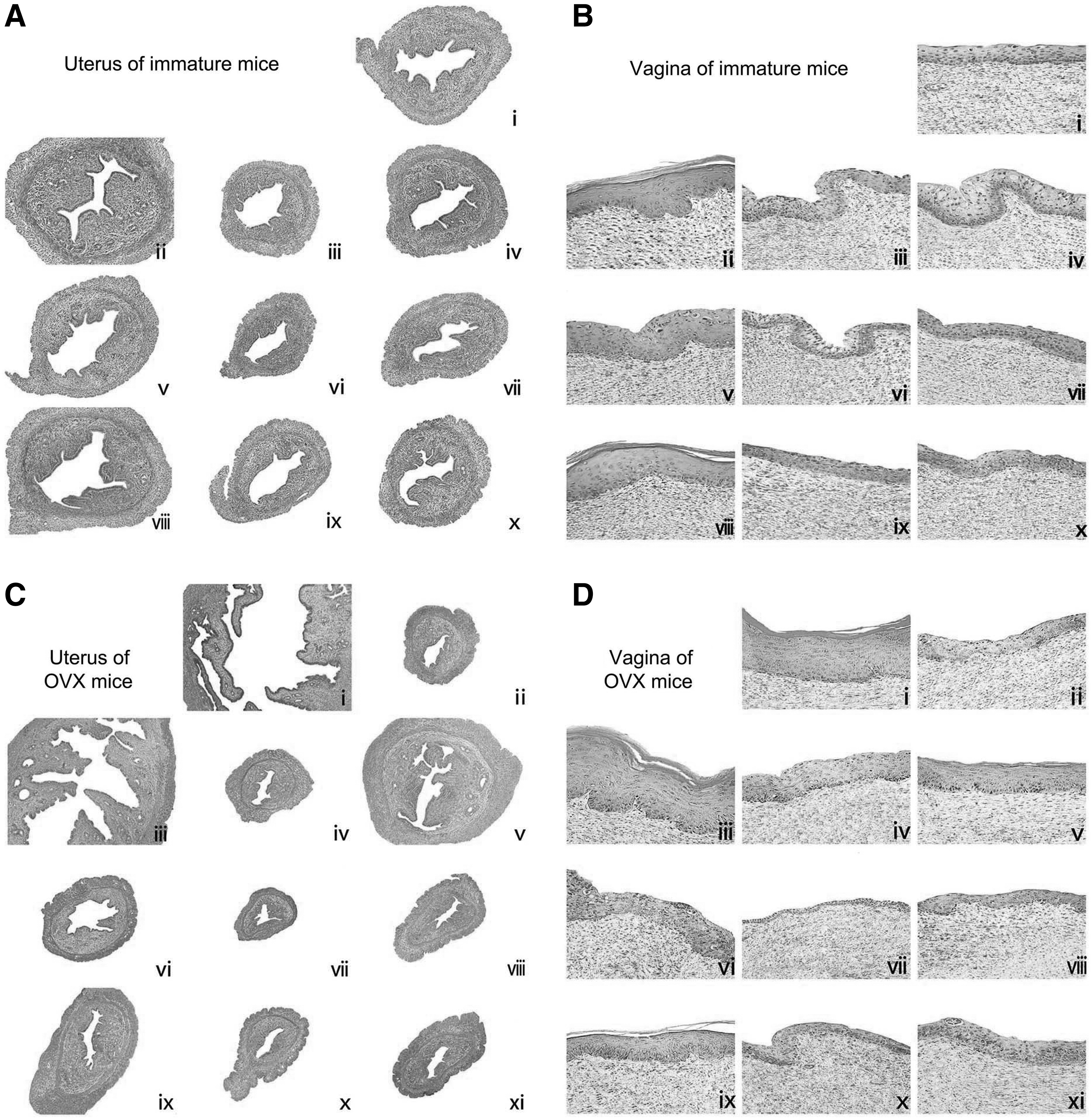
The effects of incompatibility of VN and RPA on the histology of the uterus and vagina in immature and OVX mice. Representative photomicrographs taken at 200- × magnification of uterine in immature mice; 100- × magnification of uterine in OVX mice, and 400- × magnification of vaginal sections.
Quantitative Date of Histological Feature in Uterus and Vagina
(***) p < 0.001, (**) p < 0.01, and (*) p < 0.05, compared with the Con group; (###) p < 0.001, (##) p < 0.01, and (#) p < 0.05, compared with the RPA group or E2 group; (ΔΔΔ) p < 0.001, (ΔΔ) p < 0.01, and (Δ) p < 0.05, compared with the RPA group or E2 group.
Con, control group; VN, Veratrum nigrum.
Quantitative Date of Histological Feature in Uterus and Vagina
p < 0.001, ** p < 0.01, and (*) p < 0.05, compared with the OVX group; (###) p < 0.001, (##) p < 0.01, and (#) p < 0.05, compared with the RPA group or E2 group; (ΔΔΔ) p < 0.001, (ΔΔ) p < 0.01, and (Δ) p < 0.05, compared with the RPA group or E2 group; (▴▴▴) p < 0.001, compared with the Sham group.
OVX, ovariectomized.
Figure 3B and D and Tables 1 and 2 also showed that treatment with RPA at any dose or E2 increased epithelial thickness and the number of cell layers in both mouse models, and the E2 group displayed typical squamous multilayered epithelium with cornification in immature/OVX mice. Cotreatment of RPA with VN or ICI182780 resulted in vaginal epithelium of immature and OVX mice that were composed of fewer cell layers and flattened cells with less cornification when compared with RPA treatment alone. In RPA + VN group, the effect of reversing the atrophy of uterus and vagina was antagonized, with a similar effect to RPA + ICI182780 group.
Taken together, these studies provide evidence that VN decreased the estrogenic efficacy of RPA in promoting the growth and development of the uterus and vagina in immature mice. These data prompted further studies to elucidate the molecular basis of incompatibility of VN and RPA.
Effect of incompatibility of VN and RPA on the ER subtype expression in uterus and vagina
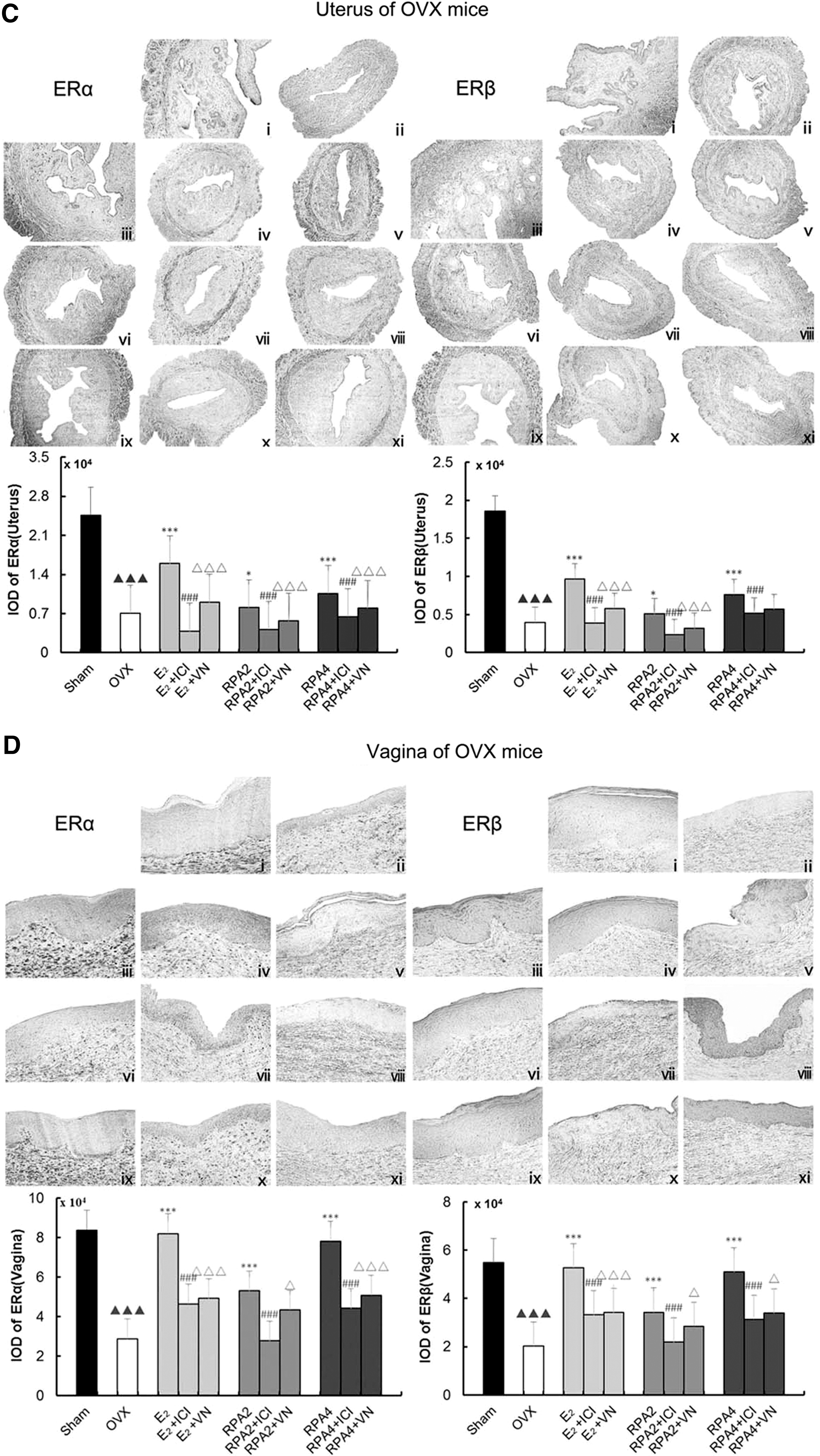
Figure 4A–D showed representative sections of the expressions of ERα and ERβ in the uterus and vagina from each group and quantitative analysis in immature and OVX mice. Mainly, ERs in the uterus were expressed in the epithelial cells of the endometrium, interstitial cells, and smooth muscle cells. ERs in the vagina were expressed in the vaginal epithelium cells of squamous and smooth muscle cells. The number of positive-expressing cells was counted and expressed as a percentage of the total number of cells. Treatment with either E2 or RPA at any dose induced a significant upregulation of ERα and ERβ in the uterus and vagina compared with the untreated group in immature (p < 0.05, 0.01, or 0.001) or OVX mice (p < 0.05 or 0.001), respectively. In RPA + VN or E2 + VN treatment group, the expressions of ERα and ERβ in the uterus and vagina decreased, with a similar effect to RPA+ICI182780 or E2 + ICI182780 group. These data indicated that VN decreased the estrogenic efficacy of RPA through ERs in vivo.
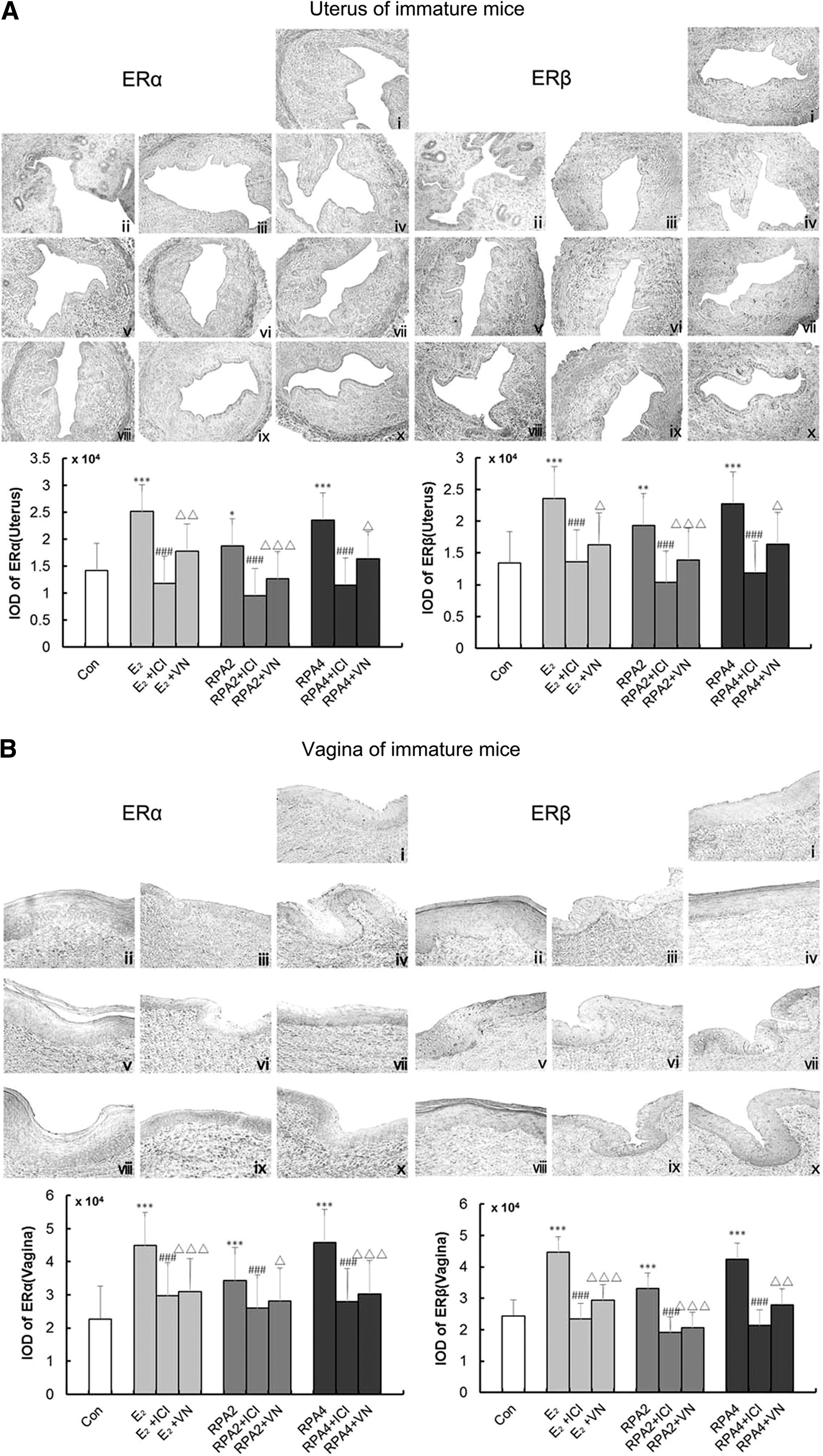
The effects of incompatibility of VN and RPA on the expressions of ER α and β in the uterus and vagina. ER expressions were assessed by immunohistochemistry. Representative photomicrographs taken at 200- × magnification of uterine and 400- × magnification of vaginal sections.
Effect of incompatibility of VN and RPA on protein and gene levels of ER subtype in uterus and vagina
Western blot and real-time quantitative PCR are used to explore further evidence for the compatibility of VN and RPA on the effects on ER subtype expressions at protein and mRNA levels in target tissues. Figure 5A–D showed that treatment with E2 or RPA at any dose induced significant upregulation of protein levels of ERα and ERβ in the reproductive tissues of immature or OVX mice (p < 0.05, 0.01 or 0.001), and also upregulation of gene expression of ERα and ERβ in immature and OVX mice (p < 0.05, 0.01 or 0.001) compared with the untreated group. RPA + VN or RPA+ICI182780 in combination induced clear and comparable downregulation of ERα and ERβ on protein and gene expression in the reproductive tissues of immature and OVX mice when compared with RPA treatment alone (p < 0.05, 0.01, or 0.001). The data provide further evidence that VN antagonism of the estrogenic efficacy of RPA in vivo through ERs was comparable to that of the estrogen antagonist, ICI182780.
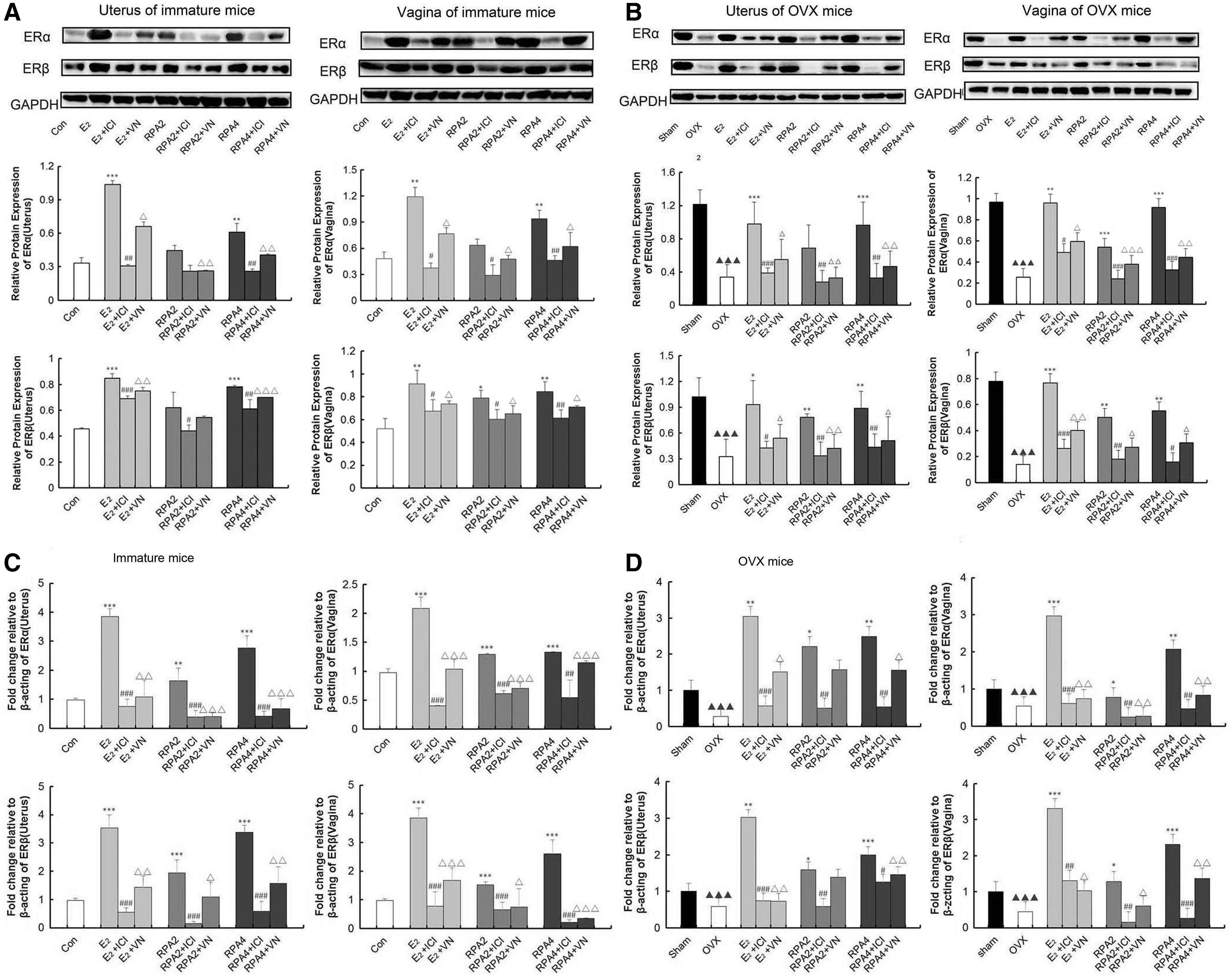
The effects of incompatibility of VN and RPA on the protein or gene expression of ER α and ERβ in the uterus and vagina of mice.
Effect of incompatibility of VN and RPA on viability of MCF-7 cell
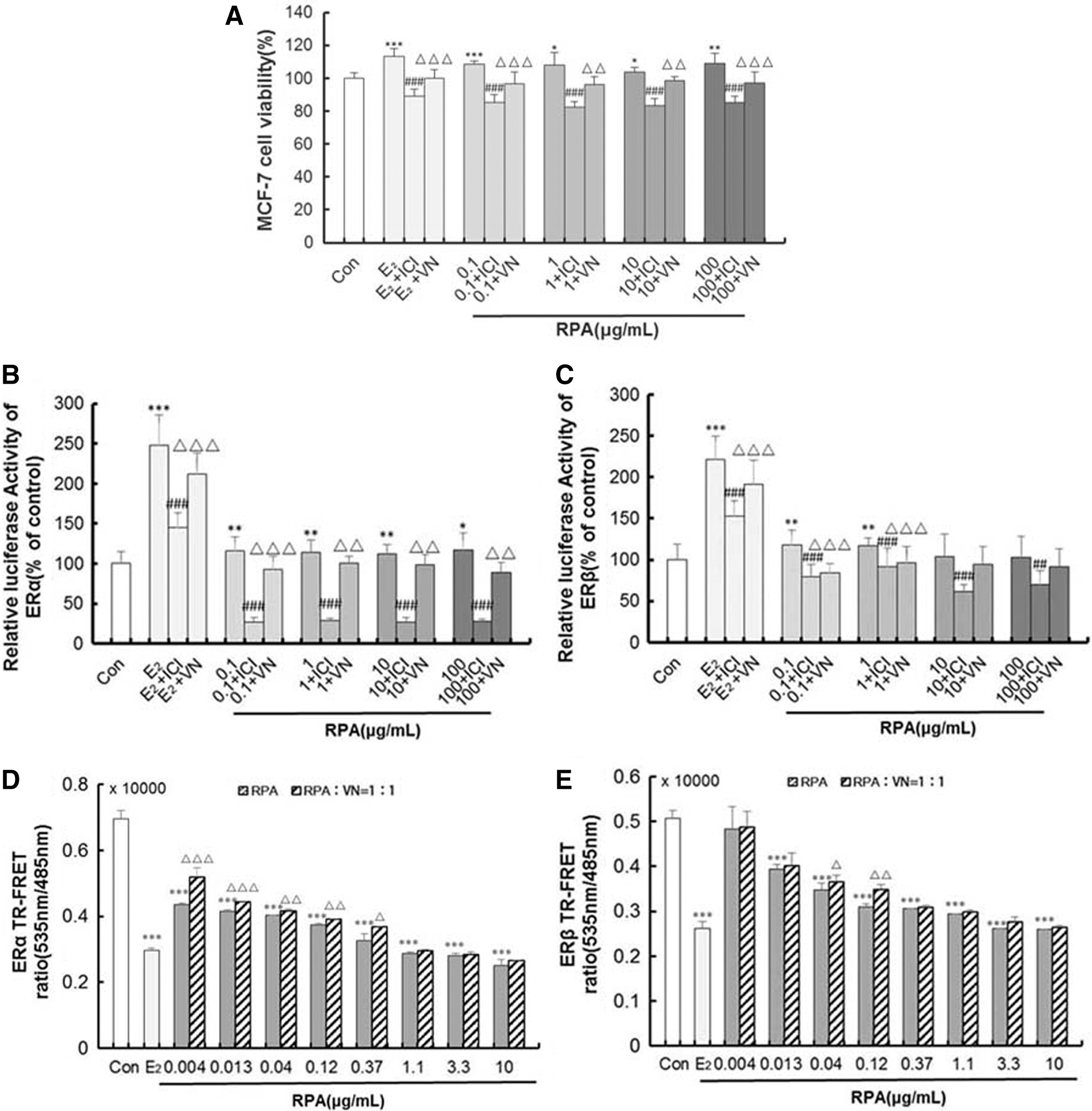
To investigate the effect of incompatibility of VN and RPA in more detail, we used MCF-7 human breast cancer cells as a model because they are dependent on estrogen for growth in monolayer culture. Figure 6A showed treatment with RPA at dose levels of 0.1–100 μg/mL and 0.01 μM E2; both stimulated proliferation of MCF-7 cell, demonstrating estrogenic activity in the RPA extracts. Compared with RPA or E2 treatment alone, the RPA + VN mixture or E2 + VN inhibited the proliferation of MCF-7 cell. The result is comparable to that observed with RPA + ICI182780 or E2 + ICI182780.
Effect of incompatibility of VN and RPA on activity of ERα and ERβ transcriptional activity
HEK 293 cells that had been stably transfected with the hERα/β-ERE-luciferase plasmid were used to measure the formation of functional hERα/β-ERE complexes in response to treatment with RPA and E2. Results are expressed relative to expression in DMSO-treated cells. RPA at the concentration of 0.1∼100 μg/mL significantly induced ERa-ERE luciferase activity, and induced ERb-ERE luciferase activity at the concentration of 0.1∼1 μg/mL compared with the Con, respectively. 0.01 μM E2 significantly induced both ERa and ERb-ERE luciferase activity compared with the Con. (Fig. 6B, C). These effects were ablated when treatments were administered in the presence of ICI182780, and the RPA + VN or E2 + VN combination treatment had the similar effect with RPA/E2 + ICI182780. These data indicate that VN inhibits the estrogenic activity of RPA that is mediated through the activation of ER-ERE.
Effect of incompatibility of VN and RPA competing with ER combining ability
The estrogenic activities of RPA were significantly inhibited by VN in vivo and in vitro. We next examined VN influence on RPA binding to ER using a TR-FRET ER competitive assay. As shown in Figure 6D, E, RPA could bind to human ERα and ERβ ligand binding domain (LBD) at a certain dose range. As the concentration increased, the combination was enhanced. RPA and VN combination treatment at the ratio of 1:1 inhibited the effect of binding to ERα (0.004–0.37 μg/mL) and ERβ (0.004–0.12 μg/mL) compared with RPA treatment alone (p < 0.05, 0.01 or 0.001).
Discussion
TCM theory recorded that the 18 incompatible medicaments were not to be used in combination as they may cause fatal consequences. Up to now, little evidence links interference effects with VN and RPA in combination use. In our previous studies, we have addressed the effects of RPA on estrogen target tissues, hot flashes, and weight gain in immature mice after short-term administration or in OVX mice after a relatively long duration of application. 13 To confirm the pharmacodynamics effect, condition, and mechanism of the incompatibility, in vivo and in vitro assays were used and were based on the estrogen-like effect of RPA. As a gold standard of estrogenic activity in vivo study, the uterine weights of mice treated with RPA + VN were much lower than RPA alone, and it is similar to the ER antagonist ICI182780. Moreover, VN has a potent effect of ER antagonist that antagonized the estrogenic efficacy of RPA in promoting development of the uterus and vagina in immature mice, and reversing the atrophy of the uterus and vagina in OVX mice.
Under physiological conditions, the biological effects of estrogen depend on not only the level of estrogen in serum but also on the distribution and expression levels of the corresponding ERs in the target cells, ERα and ERβ. 22 –24 The results showed that VN decreased the estrogenic activity of RPA on uterus and vagina by attenuating the increase of serum estradiol and the up-regulation of ERs expression in reproductive tissues caused by RPA, which suggests that VN inhibits RPA exerting estrogenic effects might be related to regulation of estrogen secretion and ER expression.
Estrogen mediates its actions by binding to the ER and inducing a major conformational change, which allows the estrogen-ER complex to relocate to the nucleus, to bind to its cognate DNA response element (ERE) located in the promoter/enhancer regions of target genes and regulate gene transcription. 25 Our in vitro results showed the RPA + VN mixture inhibited MCF-7 cell proliferation, increased the binding effect of ERα and ERβ, and increased ERα/β-ERE luciferase reporter gene expression by using RPA treatment alone. These results showed RPA and VN decreased the effects on estrogenic activity maybe relate to ER-ERE pathway.
VN at the dose of 0.045 g/kg decreased the estrogenic activity of 2 or 4 g/kg RPA in a physiological model of immature mice after short-term administration and in a pathological model of OVX mice after long-term administration. Two gram per kilogram RPA is the clinical equivalent dosage of the high recommended dose of 0.2 g/kg in humans according to Chinese pharmacopeia. The lower dose of VN of 0.045 g/kg, which is based on a low recommended dose of 0.005 g/kg in humans according to “Zhong Hua Ben Cao,” induced the more profound response on decreasing the uterine weights and serum E2 of immature mice. However, 0.09 g/kg VN is the clinical equivalent dosage of the high recommended dose of 0.01 g/kg, which did not induce the estrogen response change. This study provides the reference of administration dose and body state for the clinical use of RPA-VN in combination or alone.
In recent researches, it is reported that the LD50 of aqueous extracts of VN and RPA were 2.566 and 160 g/kg, respectively, after intragastric administration. 26 When the dose of VN is higher than or equal to RPA, acute toxicity of VN and RPA mixture increased. 14 It suggested that the increased toxicity of the VN and RPA combination was mainly due to content increase of the special veratrum alkaloids, which is one of the toxic components of VN. 27,28 In addition, besides toxicity, the inhibition of pharmacodynamics in combination use were involved in incompatibility. The study that VN appears to suppress the estrogenic effects of RPA is one of evidences of the incompatibility. VN is a potential antagonist of ERs. Although the exact effective components remain unknown, this finding encourages us to do further research on VN.
In conclusion, this is the first report of estrogenic activity on the combination of VN and RPA. VN interferes with the estrogenic efficacy of RPA by restraining the increase of serum estradiol and decreasing ER expression in target tissues and through the ER-ERE-dependent pathway. The findings of this incompatibility study on the estrogenic effects in different conditions and physiological and pathological states of body will not only provide theory reference but also increase the effects of clinical use of RPA.
Footnotes
Acknowledgments and Funding
This work was supported by the National Basic Research Program of China (973 Program) (2011CB505300, 2011CB505305).
Author Disclosure Statement
No competing financial interests exist.