
Abstract
Kaixin-San (KXS), a Chinese formula, was used to treat “amnesia,” a senile dementia in the modern world. This formula was reported to improve behavioral performances in many animal models. This study was designed to explore how KXS has improved amyloid-β (Aβ)-induced memory dysfunction in mice. The mouse models were achieved through unilateral ventricle injection with Aβ42. The effects of KXS on memory improvement were evaluated by the step-down test. The electrophysiological changes induced by KXS were measured by long-term potentiation (LTP) analysis in the hippocampus in vivo. The expression of glutamate receptor 2 (GluR2) was observed through immunohistochemical staining. Behavioral experiment outcome demonstrated reduced avoidance time and increased error time during the step-down test in the mice of Aβ group. This memory impairment, however, was reversed by KXS. Electrophysiological experiment showed no significant difference between Aβ group and KXS group either in the size or the shape of field excitatory postsynaptic potentiation recorded from perforant path to dentate gyrus pathway. However, LTP in this region was reduced by Aβ and recovered by KXS administration. Moreover, immunohistochemical staining showed increased postsynaptic GluR2 expression in DG area in KXS group. These findings suggest that Aβ results in impairment to memory function of the animals, and KXS protects the animal from memory loss by rescuing LTP through postsynaptic mechanism which refers to increasing GluR2 expression.
Introduction
Alzheimer's disease (AD) is a progressive neurodegenerative disorder. Neuropathologically, AD is characterized by amyloid-β (Aβ) protein deposition which forms senile plaques, hyperphosphorylation of the cytoskeletal protein tau which contributes to neurofibrillary tangles, and both synaptic and neuronal loss. 1 Memory and cognition impairment are major clinical manifestations for AD. 2 So far, medicinal plants, including traditional Chinese medicine herbs, have been reported to treat memory disorders for anti-AD. 2 –5
Kaixin-San (KXS), a classic Chinese herbal prescription for dementia, was first recorded to treat “amnesia” in the Chinese ancient medical prescription book Bei Ji Qian Jin Yao Fang <Thousand Formulae for Emergency> in 7th Century. 6 It contains Ginseng Radix (Panax ginseng C. A. Mey.), Poria (Poria cocos (Schw.) Wolf), Radix Polygalae (Polygala tenuifolia Willd.), and Acorus Tatarinowii Rhizoma (Acorus tatarinowii Schott). It is believed that the formula improved learning and memory functions in model animals. 7 –9 Cerebrospinal fluid obtained from rats orally administrated with KXS attenuated β-amyloid (Aβ) peptide induced nervous cell death in vitro, suggesting that KXS protected neurons from Aβ induced cytotoxicity (Our unpublished data). This may give rise to a hopeful candidate drug for Alzheimer's disease (AD). Synaptic long-term potentiation (LTP) is a critical step in memory formation. 10,11 It was reported that the hippocampal LTP was inhibited in AD model animal. 12 Our study confirmed the impairment of hippocampal LTP and memory loss induced by Aβ and found the improving effects of KXS in mice. We have also explored the mechanism by which KXS protected mice from Aβ-induced learning and memory dysfunction using behavioral experiments, electrophysiological recording, as well as immunohistochemical staining analysis.
Materials and Methods
Animals and treatment
ICR mice (25–35 g, 10 weeks old, regardless of sex) were provided by Good Laboratory Practice Centre of Heilongjiang University of Chinese Medicine (Harbin, China). Mice were maintained on a 12-h dark/12-h light cycle at an ambient temperature (24°C ± 2°C) and humidity (55% ± 5%), with access to food and water ad libitum. All animal experiments were performed according to the Guidance for the Care and Use of Laboratory Animals by the Ministry of Science and Technology of China and were approved by the Animal Experiments Ethics Committee of Heilongjiang University of Chinese Medicine (2008001). Animals were divided into three groups randomly, including control group, Aβ group, and KXS group. Seven days before the experiment, 0.2 mL KXS extracts (0.15 g/kg) were given intragastrically to the mice in KXS group once a day, while the same volume of saline was given to the mice in the other two groups.
KXS preparation
The herbs (Ginseng Radix, Poria, Radix Polygalae, and Acorus Tatarinowii Rhizoma) were purchased from Harbin Tongrentang Drug Store (Harbin, China) and authenticated by Dr. Xijun Wang, the professor of the Department of Pharmacognosy of Heilongjiang University of Chinese Medicine, and a voucher specimen of each plant (No. 1-021, No. 8-002, No. 1-028, No. 1-042, respectively) was prepared and deposited in the herbarium of the Department of Pharmacognosy.
The quality of these crude drugs from KXS was controlled as described in the Chinese Pharmacopoeia (2015). Briefly, the four ingredients, Ginseng Radix (60 g), Poria (60 g), Radix Polygalae (40 g), and Acorus Tatarinowii Rhizoma (40 g), were mixed in a weight ratio of 3:3:2:2. KXS was prepared as described previously 13 according to standard procedures. In brief, the four dried crude drugs were mixed and decocted/extracted by refluxing for 1.5 hours in 2,000 mL boiling 60% ethanol (1:10, w/v). The extracts were passed through filter paper, dried under a vacuum, and stored at −80°C. The extracts were mainly characterized by 75 chemical constituents, identified by ultra-performance liquid chromatography tandem mass spectrometry.
Other materials
Aβ42 was purchased from Sigma-Aldrich Co. LLC. (Sigma-Aldrich) and dissolved in sterile normal saline (500 μM), diluted with saline before use. The rabbit polyclonal anti-glutamate receptor 2 (GluR2) antibody and SABC Kit were purchased from Wuhan Boster Biological Technology, Ltd. (China).
Lateral ventricle injection
The mice were anesthetized with 45 mg/kg sodium pentobarbital by intraperitoneal administration and placed on a stereotaxic device. After incising the head skin, the skull was exposed and the small hole was drilled at the site (1 mm posterior to bregma, 1.75 mm lateral to midline). Then the microsyringe was inserted straight into the brain surface (1.8 mm depth to surface). Subsequently, for Aβ and KXS groups, incubated Aβ42 (concentration 1 μM) was injected into the lateral ventricle on right side for 5 μL lasting 10 minutes and retained in the site for another 10 minutes to avoid liquid pouring out, while the control group was given injection of equal volume of 0.9% saline solution. The body temperature was maintained at 36°C using a heating pad and monitored with a thermo probe placed in the rectum of the mice.
Behavioral experiment
Forty-eight hours after Aβ injection into the hippocampus, the evaluation of the learning and memory functions was carried out through behavioral tests, which were performed by a step-down test device. The device contains five isolated compartments. Each compartment is a 12 × 11 × 33 cm room with black plastic boards in the back and side and a visible plastic board in the front. The floor of the device consists of parallel metal bars, a rubber platform with 3.2 cm of diameter, and 3.3 cm of height placed in the corner of the box.
The mice were first trained for 5 minutes to leave for the safe area to avoid the electrical current stimuli, during which they were gently placed onto the floor of the test box, and then the stainless steel floor was loaded a 36 V electric voltage. The mice would escape from the electric stimuli and finally jumped on the platform. When the mice stepped down from the platform they would receive electric shock again. The memory attaining ability was confirmed 24 hours after the first training. The mice were gently placed on the platform; the avoidance time of the mice on the safe platform and the number of error (mice step down on the electrical area) during 5 minutes were counted. More than two paws step down was measured as one error.
Electrophysiological recording
Perforant path-dentate gyrus (PP-DG) pathway was used for LTP recording. Stimulating and recording electrodes were, respectively, positioned at the perforant path branch and the molecular layer of DG area of hippocampus. Two small holes were drilled at the sites of skull where stimulating and recording electrodes were positioned (for stimulating electrode: 4.5 mm posterior to bregma, 3.0 mm lateral to midline, 1.5–2.0 mm depth to brain surface and for recording electrode: 2.1 mm posterior to bregma, 1.5 mm lateral to midline, 1.8–2.2 mm depth to brain surface). Coaxial bipolar stimulating electrode was made of platinum with tip diameter of 25 μm. Glass recording electrode was filled by saline with a less than three megohms tip resistance. A reference electrode was attached to the head skin of the mice. To maintain a constant depth of anesthetization, the mice were given sodium pentobarbital (8 mg/kg) intramuscularly every 30 minutes after the first injection.
The field excitatory postsynaptic potentiation (fEPSP) was induced with a stimulating parameter (wave width of 0.6 ms and frequency of 0.067 Hz). After the baseline recording for 15 minutes, LTP was induced by high frequency stimuli (HFS, 100 trains, 1 second). Then the fEPSPs were monitored for 90 minutes continuously and analyzed using a computer-based data acquisition system. The magnitude of potentiation was expressed by the percentage of baseline of the population spike amplitudes of fEPSPs.
Immunohistochemistry
Immediately after the electrophysiological recording, the mice were decapitated, and the brains were taken out after artery perfusion with normal saline and subsequently with 4% paraformaldehyde (PFA) solution, and the brains were placed in PFA overnight. Then the brains were dehydrated and embedded into paraffin block, and the sections for histological stain were made with 4 μm thickness.
After pretreatment with 3% H2O2 for 15 minutes to quench endogenous peroxidase activity, 0.1% trypsin was used for repairing the GluR2 antigen. Then the sections were blocked by 5% bull serum albumin (BSA) for 20 minutes, followed by the incubation with the primary antibody (1:50 dilution) overnight at 4°C. Then the slices were incubated with a secondary antibody for 30 minutes at 37°C followed by StreptAvidin-Biotin Complex (SABC) distribution droplets. Between each step, sections were rinsed with phosphate buffered saline (PBS) thrice with 5 minutes each time. The sections were developed in the diaminobenzidine and made by hematoxylin counterstaining. Brain slices were then washed, dehydrated, cleared, and cover slipped. The positive cells with yellow-brown color were counted under a microscope. Sections for negative control were incubated with PBS not containing the primary antibody.
The images of sections were acquired by Nikon NIS-Elements F 3.0 software. Before taking photos we need to check at blank site with auto white. Then the immunoreactive (IR) cells in every slice were countered at the same area. The data were shown as percentage of the IR cells of the total cells in DG area in each vision.
Statistical analysis
Statistical analysis was analyzed by SPSS 17.0 software, and all data are expressed as mean ± SD. Comparisons of all data among the groups were performed by the analysis of variance (ANOVA); the comparison between two groups was carried out by two-sample t-test. p Values less than 0.05 were considered to be statistically significant.
Results
KXS ameliorated Aβ induced learning and memory impairment
Twenty-four hours after the first training, the step-down test was carried out. In the control group, the average avoidance time of the mice was 277.71 ± 58.96 seconds (n = 7), and the average error time was 0.29 ± 0.49 (n = 7). Compared with the control mice, the average avoidance time of the mice in Aβ group was significantly shortened (57.88 ± 48.59 seconds, n = 8, p < 0.01), and the average error time was significantly increased (1.75 ± 0.71, n = 8, p < 0.01). These Aβ induced memory dysfunctions, however, were reversed by KXS administration. The average avoidance time of the mice in the KXS group was dramatically increased (182.00 ± 123.19 seconds, n = 9, p < 0.05), and the average error time was also reduced dramatically (0.78 ± 0.83, n = 9, p < 0.05) compared with Aβ group. The results indicated that Aβ impaired the memory formation and KXS reversed the memory dysfunction induced by Aβ. (Fig. 1).
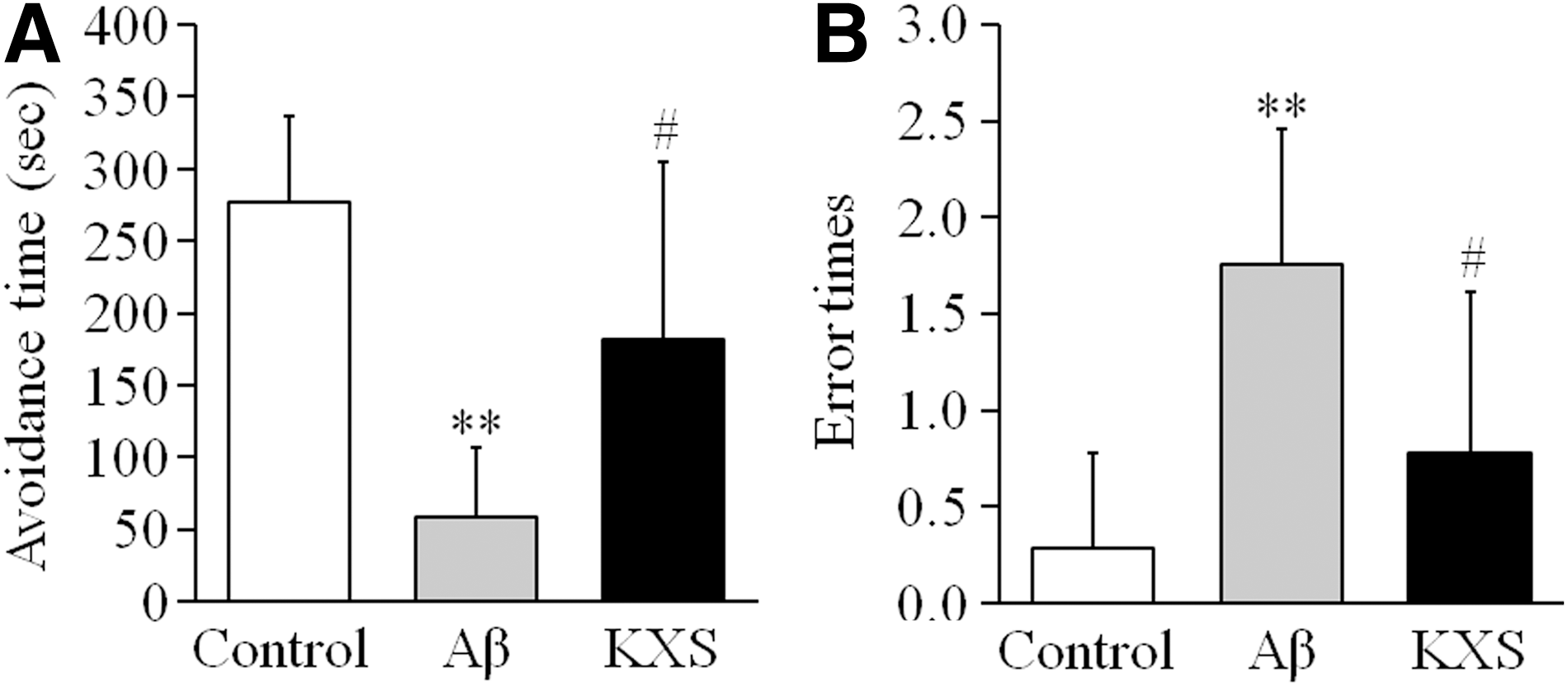
KXS reversed the memory dysfunction induced by Aβ in mice in the step-down test. Twenty-four hours after the first training, the step-down test was carried out. Both the avoidance time and error times were counted for evaluation of learning and memory functions.
KXS improved the Aβ induced inhibition of hippocampal LTP in mice
The formation of hippocampal LTP is a critical step for memory formation. The results above in the step-down test indicated that unilateral ventricle injection of Aβ42 led to memory loss in mice, while KXS improved the memory dysfunction induced by Aβ42. This suggested that KXS could improve the inhibition of LTP in neuronal synaptic transmission. Therefore, we have recorded fEPSP in mice in vivo. The results showed that there was no difference in fEPSP waveforms induced by basic stimulus among the control group, Aβ group, and KXS group. However, after HFS were delivered to the perforant path branch, LTP generation with the feature of amplitude and rising slope of fEPSP increasing were observed in all the mice of the three groups. Eighty minutes after HFS, the average of magnitude of amplitude of fEPSP population spike in mice of the control group became 137.30% ± 21.49% (n = 20) higher than baseline (before HFS). This value was only 21.08% ± 12.02% (n = 6), higher than baseline in Aβ group, which is about one-sixth (1/6) of the control group. The statistical analysis revealed significant difference between these two groups (p < 0.01). These results suggested that Aβ42 impaired synaptic plasticity in hippocampal neurons and caused mice learning and memory dysfunction. Interestingly, the average of amplitude of fEPSP population spike in mice in KXS group was increased by 93.93% ± 28.01% (n = 7), 4.5 times higher than the value of Aβ group (p < 0.05). This indicated that KXS tended to rescue the impaired LTP induced by Aβ42 in hippocampal neurons and consequently improved the memory function in mice (Fig. 2.).

KXS improved Aβ induced hippocampal LTP impairment in vivo. After recording stable fEPSP for 15 minutes as baseline, HFS were given to induce LTP and then LTP was continuously recorded for 80 minutes.
KXS reversed the low expression of GluR2 induced by Aβ in DG area
As shown in Figure 3, GluR2 IR cells were marked with brown color. The cells were almost zonally distributed in the site of DG area where recording electrode was positioned, suggesting that the artificial electrical stimulation could increase the postsynaptic GluR2 expression. The immunoreactivity was rather weak in the Aβ group mice than those in both control group and KXS group. The IR cells of GluR2 (GluR2 IR cells) were counted in a small square. The average number of GluR2 IR cells in Aβ group was 10.30 ± 4.56 (n = 15), which was significantly decreased compared with the control group (69.47 ± 9.86, n = 10, p < 0.01). However, an increasing number of GluR2 IR cells were observed in KXS group (36.32 ± 9.28, n = 18, p < 0.01) compared with Aβ group.

KXS upregulated the expression of GluR2 on postsynaptic membrane.
Discussion
The present study strongly supports the hypothesis in traditional Chinese medicine that KXS ameliorates memory impairment. We have provided convincing evidence in vivo that KXS enhances the LTP which is inhibited by Aβ. Moreover, our findings indicated that the increasing expression of GluR2 on postsynaptic membrane has played an important part in the KXS effect on improving LTP inhibition.
Aβ plays a main role in the pathogenesis of AD. 14 Recent studies have found that the cognitive function was impaired in the early stage of AD without the neuron loss and senile plaque. 15 It has been confirmed that Aβ acted as a neurotoxin combined with neuron synapses leading to their dysfunction and finally induced memory loss and other neuronal injuries. 16,17 Electrophysiological studies found that Aβ inhibits neuronal synaptic transmission plasticity, which is a critical step of memory formation. 18,19 Our findings are in accordance with these previous studies that the inhibition of LTP induced by Aβ is responsible for learning and memory dysfunction. 20,21
As main glutamate receptor, AMPA receptor (AMPAR) plays a key role in the LTP formation. 22 Aβ induced AMPAR dysfunction leading to the impairment of glutamatergic neuron synaptic transmission. 15 In vitro, the expression of AMPAR on postsynaptic membrane decreased in the cultured APP transgenic neurons, as well as Aβ added cultured wild-type neurons. 23 It can be considered that this is due to an Aβ-induced decrease in the circulating levels of GluR2 onto the dendrite cell surface in the postsynaptic neurons. 24
Previous research has found that KXS improved cognitive function of AD model animals by enhancing cholinergic nervous system function in the brain. In addition, KXS could improve the ability of antioxidation and stress in animals by increasing the activity of SOD and clearing the superoxide radicals. Although these observations of KXS effects on neuron degenerations were demonstrated, yet the neuroelectrophysiological mechanism that has contributed to KXS improving the memory dysfunction induced in the early step of AD still remains unclear.
In the present study, we have first confirmed that KXS improved the Aβ induced memory dysfunction by enhancing the hippocampal LTP in vivo in mice. We then further explored the reasons as why LTP was inhibited by Aβ or enhanced by KXS. Reassuringly, one point we can ensure is that LTP maintenance is associated with mechanism of postsynaptic membrane. We have noticed that KXS has increased the expression of GluR2 contributing to relieve LTP inhibition, and consequently, failure of memory in mice was improved. This is consistent with the findings that spatial learning and memory were impaired in GluR2-cKO mice 25 and suggest that in terms of electrophysiology, the neurophysiological mechanism of KXS improving memory in AD model animals is preventing Aβ induced inhibition of synaptic transmission LTP by increasing the expression of GluR2 on the postsynaptic membrane. The presence of GluR2 preventing excitatory neurotoxicity may also be responsible for our discoveries. 24
The chemical ingredients of traditional Chinese medicine prescription are complex. As a traditional formula, KXS may act through multiple mechanisms, including postsynaptic, presynaptic, and other mechanisms, because traditional Chinese herbs have the feature of multifactor and multitarget action on diseases or pathogens. 26 It will be a huge project to explore the relevant mechanisms in detail. In the present study, we have focused on the postsynaptic mechanism of KXS improving the memory function by enhancing the hippocampal LTP. And we found that the trafficking of GluR2 was inhibited by Aβ and recovered by KXS in vivo. These are related to the cognitive changes. GluR2 as an important member of AMPARs is a key mediator of synaptic plasticity on postsynaptic membrane 27 for the formation and stabilization of LTP. 28 The balance of externalization, anchoring, and internalization of GluR2 is important for LTP and is affected by many factors through different mechanisms. 29 KXS may act on these trafficking steps for LTP enhancement. However, further studies are needed for us to explain the mechanisms in detail.
In conclusion, KXS improves memory dysfunction by enhancing GluR2 expression on the postsynaptic membrane by which synaptic LTP recovers from the impairment by Aβ administration. KXS would be a beneficial agent for AD therapy.
Footnotes
Acknowledgments
This work was supported by National Natural Science Foundation of China (No. 81603321, No. 81303248), Scientific Research Foundation of Heilongjiang University of Chinese Medicine (No. 2015bs04, No. 2012RCQ62), Natural Science Foundation of Heilongjiang Province (No. H2016072, No. LC2008C26), and University Nursing Program for Young Scholars with Creative Talents in Heilongjiang Province (No. UNPYSCT-2017216).
Authors' Contributions
Bo Zhang participated in all experimental work; Yan Li and Jiawei Liu were involved in the step-down test and immunohistochemical staining; Xuewei Liu and Wei Wen contributed to electrophysiological experiment; and Yu Cui and Shuming Huang designed the experimental protocols and prepared the article.
Author Disclosure Statement
The authors have no competing or conflicting interests to declare.