
Abstract
The efficient use of stem cells for transplantation is often limited by the relatively low number of stem cells collected. The ex vivo expansion of human stem cells for clinical use is a potentially valuable approach to increase stem cell number. Currently, most of the procedures used to expand stem cells are carried out using a 21% oxygen concentration, which is about 4- to 10-fold greater than the concentration characteristic of their natural niches. Hyperoxia might cause oxidative stress with a deleterious effect on the physiology of cultured stem cells. In this review, we investigate and critically examine the available information on the ability of natural compounds to counteract hyperoxia-induced damage in different types of stem cells ex vivo. In particular, we focused on proliferation and stemness maintenance in an attempt to draw up useful indications to define new culture media with a promoting activity on cell expansion in vitro.
Introduction
Human stem cells are being investigated for the treatment of many different diseases, like cardiovascular and autoimmune diseases and traumatic lesions. 1 Among stem cells, particular interest has been focused on human mesenchymal stem cells (hMSC) that can be derived from many tissues but are often insufficient to reach a relevant number of cells adequate for clinical use. Moreover, patient age, gender, genetic background, and clinical history influence their quantity and quality.
At present, most procedures to expand stem cells are carried out at an ambient oxygen concentration where cells are exposed to 21% O2, which is 4- to 10-fold greater than the concentration characteristic of their “in vivo” niches. 2,3 Several studies have demonstrated the deleterious impact of a 21% oxygen tension on the physiology of stem cells, for example, neuronal stem cells, 4 bone marrow stem cells, 5,6 umbilical cord stem cells, 7 or adipose tissue stem cells. 8,9 In fact, in vitro culture of MSCs exposed to a 21% oxygen concentration might increase the production of reactive oxygen species (ROS) and, as a consequence, to be subjected to environmental oxidative stress. In general, ROS, including hydrogen peroxide (H2O2) and superoxide anion (O2 −), plays crucial roles in stem and progenitor physiology. Nevertheless, the “redox window” hypothesis must be considered: suitable ROS level is vital for physiological cellular functions, 10 whereas excess ROS contribute to adverse effects on MSCs, including premature senescence, increase of the population doubling time, damages to DNA, 11,12 and reduced engraftment after transplantation. 13 Human MSCs are particularly susceptible to early senescence and typically halt proliferation at approximately passage 8, which limits their expansion potential and therapeutic value. 14 Another issue to be considered is the need of long-term cryopreservation of stem cells, a routine practice that requires extremely controlled and safe techniques of specimen storage. However, more and more studies are showing contradictory results on the influence of stem cell cryopreservation and thawing, such as wide stresses at physical and biological level, mitochondrial damages, modifications to basal respiration and ATP synthesis, apoptosis and necrosis, shortening of telomeres and cellular senescence, damage to DNA, and oxidative stress. 15 These results have raised serious alarms concerning the therapeutic effectiveness and safety of stem cells. Ability to enhance MSC culture expansion and delay senescence could substantially improve efficacy for autologous stem cell therapy. On these bases, a promising strategy to counteract oxidative stress induced by hyperoxia or by cryopreservation and thawing could be the use of compounds with antioxidant activity. In particular, many natural antioxidant compounds have been demonstrated to counteract oxidative stress by directly scavenging free radicals or activating antioxidant defense pathways. 16 –18 Moreover, the safety of many natural antioxidant compounds has been largely demonstrated 19 –21 and this is an important issue considering that stem cell expansion has principally performed with the intent of implantation.
Another important issue to take into consideration in using natural antioxidant compounds to counteract oxidative stress in stem cells is that these compounds can induce changes in cells that should be checked before using them for treatment. A good antioxidant compound should not impair stem cell stemness, potency, and proliferation. In this review we investigate and critically examine the available information on the ability of natural compounds to counteract hyperoxia-induced damage in different type of stem cells ex vivo. In particular, we focused on proliferation and stemness maintenance in an attempt to draw up useful indications to define new culture media with a promoting activity on cell expansion in vitro.
From the Niche to the Bank
In living organisms, stem cells reside in a specific microenvironment called niche. Quiescent stem cells are located in the niche until specific signals finish the dormant state and activate the differentiation process. In particular, this dynamic microenvironment accomplishes primarily three functions, thanks to secreted or cell surface molecules: stem cell proliferation control, stem cell daughters fate determination, and protection of stem cells from exhaustion or death. 22 Not only structural and soluble factors, but also hypoxia plays a crucial role for the stemness maintenance of mesenchymal, embryonic, neural stem, and hematopoietic cells, and could also impact on proliferation and differentiation. 23
Oxygen tension in the niche
In MSC niches, the local oxygen concentrations fluctuate depending on stem cell type: between 10% and 15% O2 in the adipose tissue, 24 about 1.3% both in the amniotic fluid and amniotic membrane, 22 and between 1% and 7% O2 in bone marrow. 25
The bone marrow niche is the most studied in literature. Although bone marrow has a highly complex vascular network, the entire bone interior is relatively hypoxic, with pO2 in the range of 1%–4%. 26 In general, HSCs stay quiescent in the bone marrow niche, whereas various stimuli that cause loss of quiescence induce strong entry into the cell cycle, and trigger proliferation frequently accompanied by stress, apoptosis, and damage to DNA. 27,28 Zhou et al. 29 observed that a low-oxygen niche in bone marrow reduces ROS generation, therefore providing long-term protection against oxidative stress. Ex vivo expansion techniques should promote symmetric stem cell divisions, 30 and consequently stem cell self-renewal without inducing differentiation and apoptosis.
Indeed, oxygen concentration has a major role in the proliferation and differentiation of stem cells, also in the placenta. It has been demonstrated that cytotrophoblasts proliferate in low-oxygen tension (2% O2), whereas they differentiate in high-oxygen tension (20% O2). 31
HIF role in the niche
The reduction of oxygen tension below a specific threshold leads to the initiation of a gene expression program by the transcription factor, hypoxia-inducible factor-1 (HIF-1). HIF-1 signaling pathway plays a fundamental role in regulating key processes such as energy metabolism, angiogenesis, cell proliferation, and viability depending on the availability of oxygen in the cell (reviewed in Ref. 32 ). The low-oxygen tension stabilizes HIF-1, the oxygen-sensitive monomer of the HIF-1, since its ubiquitination and proteasomal degradation are prevented. 33,34 HIF-1 is linked to transcription of at least 70 genes holding functional hypoxia-responsive element. Most of these genes are involved in cell survival, through the expression of cell surface receptors, angiogenic growth factors, glucose transporters, and glycolytic enzymes. 34,35 Moreover, under low-oxygen tension stem cell pluripotency is maintained by HIF-2 interaction with the promoter region of OCT4 causing increased transcriptional activity. 36 OCT4 is the principal regulator of pluripotency in embryonic stem cells (ESCs) and essential to obtain induced pluripotent stem cells (iPSCs) but disappears upon differentiation. 37
For example, bone marrow HSCs (BHSCs) are quiescent in vivo through conservation of a hypoxic phenotype with high levels of HIF-1α protein and the hypoxic marker pimonidazole, even when contiguous to arterioles. 38 Thus, in preserving quiescence into the niche, HSCs must depend on anaerobic glycolysis for energy production. 39 Indeed, glycolysis is induced by HIF-1α in response to hypoxia, expressing pyruvate dehydrogenase kinase 1 (PDK1), which inhibits pyruvate dehydrogenase and avoids mitochondrial acetyl-Co-A production and oxidation. The clear result is a metabolic shift that induces glycolysis and decreases ROS generation in hypoxia. Correspondingly, expansion of MSCs in atmospheric oxygen levels upregulates OXPHOS genes and mitochondrial respiration, and accelerates cellular senescence, while hypoxic culture conditions delay senescence. 40
ROS unbalance and senescence
It is known that the use of O2 in several fundamental metabolic processes by living organisms has got an evolutionary price, as O2 metabolism is the main source of ROS in the cells. The condition known as oxidative stress occurs when the endogenous production of ROS exceeds the cellular capacity of the antioxidant systems. Unfortunately, not always the cellular adaptive processes are able to counteract the accumulation of oxidative damage to key molecules, such as lipids, DNA, and proteins. 29 While all the cells of an organism might be subjected to the accumulation of damages triggered by oxidative damage, the effects of ROS on stem cells in most self-renewing tissues are of specific interest to the processes of aging and cancer development owing to their undifferentiated condition and longevity of replicative potential. 41,42
Excessive ROS production leads to the overexpression of the tumor suppressor genes, p21, p53, and p16. 43,44 Accordingly, oxidative stress is one of the principal causes of physiological decline during cellular senescence. Forkhead box (Fox) O proteins control ROS generation in stem cells by regulating catalase and SOD expression. 45,46 It has been observed that the increase of ROS level in FoxO-deficient mice is associated with the disruption of HSC quiescence, increase in HSC apoptosis, and impairment in hematopoietic repopulating abilities.
In animal models with damaged cellular antioxidant defense system, such as SOD2, an antioxidant enzyme that is present in mitochondria and modulated by FoxO3, a potential correlation between ROS and HSC impairment has been observed. SOD2 evoked a protective activity in mouse neural stem cells (NSCs), and its upregulation improved NSC's survival both in vitro and in vivo. In addition, the alterations related to aging in SOD2 modulation by FoxO3 and DNA damage, recognized by serine/threonine protein kinase ATM (ataxia telangiectasia mutated), contribute to HSC functionality reduction with aging. 46 HSC self-renewal capacity relies on ATM-mediated reduction of ROS production as demonstrated by studies carried out in Atm −/−. There is a crosstalk with the ATM-p53 signaling converging on p16 to trigger stem cell senescence. 47
Other key modulators of aging, oxidative stress, and stem cell functionality are sirtuins (SIRT), a family of NAD-dependent protein deacetylases. Recently it has been suggested, a role for SIRT1 in maintaining MSC growth and differentiation that have been observed to deteriorate with age. Interestingly, its activation in aged cells protects the cells from p53-dependent apoptosis or senescence. 48 SIRT3, is crucial in the aged HSCs in the maintenance of mitochondria metabolism and in controlling oxidative stress. Moreover, ectopic upregulation of SIRT3 enhances the function of HSCs during aging by increasing SOD2 antioxidant activity. 49
Modifications of culture conditions
On these bases, the aging of stem cells may be controlled by regulating their metabolic and redox state, which in turn might impact on the intracellular accumulation of ROS. The condition defined as “cell culture stress” might induce genetic instability and therefore constitutes a biological risk for cell therapy protocols. These pathways should be reversed or avoided to obtain an efficient stem cell expansion in vitro.
Recently, the increasing interest in the use of MSCs in the clinical practice has led to a high number of studies on the effects of culture conditions and expansion on pathways, behavior, and function of stem cells. As in vivo HSC self-renewal and differentiation are regulated by niche cells secreting cytokines and growth factors, including pleiotrophin, angiopoietin-1, -3, thrombopoietin, granulocyte colony-stimulating factor, stem cell factor, and Notch ligands, 50 cytokines were the first drugs studied for HSC ex vivo expansion. Overall, cytokines and their combinations can maintain HSCs and progenitors counteracting apoptosis in in vivo proliferation but leading to a contained human HSCs (hHSCs) expansion. For this reason, further elements are obviously essential for a positive expansion of hHSCs ex vivo.
The impact of oxygen concentration in adult stem cell phenotype, morphology, proliferative capacity, and functionality has been largely investigated with controversial outcomes.
For example, MSCs cultured in low oxygen tension (1%–10% oxygen) has revealed enhanced regenerative/reparative properties in animal models of heart, brain, and lung damage. 51 –53
In particular, several studies have shown that human stem cells cultured in a range of 1%–5% of oxygen tensions showed improved cell proliferation and longer lifespan. 5,12,54,55 Culturing in hypoxia have also been demonstrated to avoid double-strand breaks (DSB) and chromosomal abnormalities in several types of stem cells. 56,57 In agreement with this notion, Estrada et al. 11 showed that, compared with cells grown at 3% O2, short-term growth of hMSC at 20% O2 tension enhances oxidative stress and DNA damage markers, such as DSB, chromosomal aberrations, aneuploidy, and telomere shortening rates. In this study, they showed that expansion at 3% O2 significantly increases the growth rate of adipose tissue-derived hMSC with a better conservation of telomere length. However, telomerase activity in hMSC was low and insensitive to O2 concentration suggesting that the protection from oxidative stress, due to the growth at 3% O2, avoids telomere erosion.
Moreover, hypoxia upregulated the expression of OCT4 and CXCR7 in MSCs, and deferred modifications in phenotypic characteristics such as morphology, cell size, and senescence. 58 Furthermore, besides these effects, hypoxia also increased the levels of MSC-secreted factors, such as matrix metalloprotease-(1 and 3) and secreted lysyl oxidase (LOX), which are required for migration and cell matrix adhesion. 59
However, other authors described different results. For example, Pezzi et al. 60 found that cells exposed to protracted hypoxia behaved similar to their controls in normoxia concerning viability, immunophenotyping, differentiation, and ROS levels. Cells subjected to low oxygen tensions (4%–1%) showed lower proliferation, smaller cell size, greater cellular complexity, lower mitochondrial activity, and autophagy. These data validate a study by Holzwarth et al., 61 that, by culturing MSCs in conditions of 21%, 5%, 3%, and 1% oxygen, concluded that hypoxia limits MSC's proliferation in incubation period of 7 days. This inconsistency could be due to heterogeneity of culture conditions, such as supplementation and period of exposure to hypoxia, as well as intradonor variability that may explain the contradictory effects of hypoxia on the proliferation and differentiation capacities of MSCs. 61 –64 Furthermore, donor's age directly worsens cell proliferative capacity, concurring to the variability of results. 65
Supplementation with molecule affecting ROS levels has been tested in many studies. For example, El Alami et al. 66 demonstrated that for human dental pulp stem cells (hDPSCs) cultured under 21% O2, oxidative stress promotes the sequential activation of p38 MAPK, p21, and the nuclear factor erythroid 2-related factor 2 (Nrf-2) antioxidant defense pathway. The antioxidant properties of p21 depend on Nrf-2 activity that upregulates HO-1 and NQO1. Therefore, incubation with Trolox, a water-soluble analog of vitamin E, or with a p38 inhibitor, reestablishes high proliferation rate of hDPSCs even under 21% O2.
Antioxidant Defense System in Stem Cells
ROS include nonradical species, such as hydrogen peroxide and singlet oxygen, as well as radical species (superoxide, hydroxyl, hydroperoxy, peroxy, and alkoxy radicals). Biologically, superoxide and hydrogen peroxide play a pivotal role in modulating redox signaling. 67 The effects exerted by these molecules is related to their nature, intracellular levels, and localization: ROS can be involved not only in macromolecule's oxidative damage and cell death induction, but also in signal transduction linked to cell proliferation and gene transcription activation. 68 In particular, numerous experimental evidences suggest a clear correlation between intracellular H2O2 levels and features of stem and progenitor cells. 10,41,69 –71 As is well known, excess ROS contributes to pathological conditions, nevertheless, a proper and controlled ROS production is mandatory for physiological cellular functions, the so-called and previously cited “redox window” hypothesis. 72,73
ROS sources in stem cells
Much remains to be learned about the identification of the main ROS source in stem cells. In mammalian cells, a number of ROS sources are known, including: mitochondrial OXPHOS, 67,74 plasma membrane NOXs, 75 cytochrome P-450, and b5 enzymes at endoplasmic reticulum level, 76 peroxisome oxidases, 77 lipoxygenases, 78 and xanthine oxidase. 79
The fewer mitochondria content of stem cell compared with their progeny is in accordance with their utilization of glycolysis rather than OXPHOS for ATP production. 39,80 Since it is often assumed that mitochondria are the main ROS source, 67 members of NOX family could represent major cellular sources of ROS in HSCs, as also suggested by Drehmer et al. 81 in relation to the metabolic switches for adipogenic stem cell differentiation. hHSCs indeed express NOX1, 2, and 4 and various regulatory subunits of NOXs 80,82 and Piccoli et al. estimated that NOX-mediated extramitochondrial O2 consumption accounts for about half of the endogenous cell respiration in hHSCs. 80 Interestingly, mouse BHSC-enriched LSK-cells express NOX1, 2, and 4, whereas NOX4 is not present in HPCs, Lin− cells, and mononuclear cells from mouse BM, suggesting that NOX4 expression is downregulated during HSC differentiation and that NOX4 may play an important role in the regulation of HSC function. 83 However, the increase in ROS production in HSCs is associated with upregulation of NOX4 expression, 83 but is not related to mitochondrial dysfunction nor elevated mitochondrial superoxide production. 84
Nevertheless, it has been shown that cells, including HSCs, from Bmi1−/− mice exhibit abnormal mitochondrial function with consequently an increased ROS production. 85 In addition, an elevation of mitochondrial biogenesis and oxidative activities seems to be related to an increased production of ROS in HSCs from Tsc1−/− mice. 86
ROS derived from various cellular sources and compartments could be used by different stem cells to perform unique functions. For example, Udx1 (a dual NOX), allows the conversion of O2 •− to H2O2 to prevent polyspermy and regulate cleavage in the zygote of the sea urchin. 87 Cardiovascular differentiation of ESCs depends on increased ROS generation derived from various NOXs. 88 However, an increase in mitochondrial mass and production of ATP and ROS is linked to human ESC (hESC) differentiation. 89
Endogenous antioxidants in stem cells
To regulate redox signaling and to counteract excessive amounts of ROS derived from aerobic metabolism, a complex antioxidant system plays an important role. In particular, MnSOD plays a primary role in the cell protection from mitochondrial ROS. Nevertheless, although MnSOD has been extensively studied in relation to aging and tumor biology, the crucial role of MnSOD in stem cell physiology still remains unclear. 90 The cytosolic Cu/Zn SOD isoform not only is present in stem cell but also represent a factor secreted by MSCs. Importantly, MSCs have been reported to restore the radiation-induced reduction of Cu/Zn SOD levels after whole thorax irradiation, suggesting a paracrine action exerted by MSC-derived SOD1 in the protective function exerted by MSC. 91
Outside mitochondria, the glutathione-based systems (GSH/GSSG ratio regulated by glutathione S-transferase, glutathione peroxidase/reductase) usually represent the major redox-regulatory mechanisms leading to H2O2-scavenging mechanism involving its reduction to water and a similar reaction is catalyzed by CAT inside the peroxisome.
Thioredoxin is mainly localized in the endoplasmic reticulum and reduces disulfide bonds of oxidatively modified proteins. Sundaramoorthy et al. 92 demonstrated that the treatment with TXN, 24 hours following irradiation, mitigates radiation-induced cell injury of hematopoietic stem in mice.
Natural Compounds to Optimize Expansion of Stem Cells In Vitro
Many studies have explored the effect of natural antioxidant compounds against hyperoxia-induced oxidative stress in ex vivo cultures of different stem cells to increase cell proliferation, maintain potency, counteract senescence and DNA damages (Table 1). In particular, purified phytochemicals and their potential in vivo metabolites, plant extracts, combinations of purified compounds, and/or extracts have been investigated.
Phytochemicals with a Positive Effect on Stem Cell Proliferation and Stemness Maintenance In Vitro
3D, three-dimensional; ASCs, adipose-derived stem cells; ERK, extracellular signal-regulated kinase; hAFSCs, human amniotic fluid-derived stem cells; hBMSCs, human bone mesenchymal stem cells; hESCs, human embryonic stem cells; iPSCs, induced pluripotent stem cells; mBMSC, mouse bone marrow stem cells; mESCs, murine embryonic stem cells; mMSCs, mouse mesenchymal stem cells; NSCs, neural stem cells; ROCK, rho-associated protein kinase; STAT3, signal transduction activator transcription factor 3; rASCs, rat adipose stem cells; rBMSCs, rat bone marrow stem cells; rNSCs, rat neuronal stem cells; rNSPCs, rat neural stem/pre-cursor cells; mNSCs, mouse neuronal stem cells; hNSCs, human neuronal stem cells; hASCs, human adipose stem cells; TR, thioredoxin reductase.
Purified phytochemicals
Many purified phytochemicals have been studied for their ability to enhance proliferation and maintain stem cell potency, and the majority of them are polyphenolic compounds, in particular flavonoids (Fig. 1).
Chemical structure of phytochemicals used to optimize expansion of stem cell in vitro.
Epigallocatechin gallate
Epigallocatechin gallate [(−)-cis-3,3′,4′,5,5′,7-hexahydroxyflavane-3-gallate] is the most abundant and biologically active catechin in green tea. This compound is a polyphenol belonging to the flavonoid family whose members are known for their in vitro and in vivo antioxidant activity. 93 Wang et al. 94 showed that epigallocatechin gallate treatment (10–40 μM) significantly increased cell proliferation of adult hippocampal neural progenitor cell cultures. Another study demonstrated that low concentrations (0.1 μM) of epigallocatechin gallate increased the number of dividing cells and stimulated neurosphere formation of cochlea NSCs, confirming the positive effect of this flavonoid on stem cell proliferation. 95
Quercetin
Quercetin (3,3′,4′,5,7-pentahydroxyflavone) is a natural flavonoid present in many edible plants, including grapes, apples, onions, and berries. 96 The effect of this flavonoid, along with its major in vivo metabolite, quercetin-3-O-glucuronide, has been considered in cultures of human embryonic NSCs. 97 The results showed that quercetin and quercetin-3-O-glucuronide have totally opposed effects on cell viability. Quercetin (20–60 μM) decreased NSC viability, decreased Akt phosphorylation, and increased apoptosis. In contrast, quercetin-3-O-glucuronide (20–60 μM) increased proliferation through the Akt/cyclin D1 and BDNF signaling pathway. Quercetin's negative effect on cell viability was also observed in hESCs. In particular, quercetin (25–200 μM) induced mitochondrial ROS production that were responsible for quercetin-mediated hESC cell death. 98 From our point of view, as quercetin is a very active compound, the range of concentrations tested in these studies are too high to evoke a proliferative effect. Further studies should be carried out to assess lower quercetin concentrations on NSCs viability. Our hypothesis is reinforced by the study of Nichols et al., 99 which showed that 0.3–3 μM quercetin, but not 10 μM quercetin, increased cell viability of primary cortical neuron subjected to oxygen–glucose deprivation.
Kaempferol
Kaempferol (3,4′,5,7-tetrahydroxyflavone) is a flavonoid structurally similar to quercetin apart from the lack of the hydroxyl group in 3′ position. The treatment with high concentrations (200 μM) of kaempferol negatively affected murine ESCs (mESCs) viability and growth and increased mitochondrial ROS, although the resulting cell colonies were smaller but remain pluripotent. 100 Nevertheless, lower concentrations (20 μM) of this flavonoid increased the pluripotency marker expression in mESCs but, at the same time, inhibited proper differentiation of mESCs.
Naringenin
Naringenin (4′,5,7-trihydroxyflavanone) is a flavonoid compound belonging to the flavanone class mostly present in citrus fruits, including lemon, orange, tangerine, and grapefruit. 101 The supplementation of naringenin (1–100 μg/mL) to human bone MSCs (hBMSCs) resulted in enhanced cell proliferation. 102 The activity of naringenin was also studied in human amniotic fluid-derived stem cells (hAFSCs) by Liu et al. 103 In agreement with the previous study, in the presence of naringin, hAFSC proliferation increased dose dependently between 1 and 100 μg/mL. Unfortunately, no mechanisms have been suggested to explain the observed effect on cell proliferation.
Icariin and its derivatives
Icariin (4′-O-methyl-8-γ,γ-dimethylallylkaempferol-3-rhamnoside-7-glucoside) and its derivatives are flavonoids obtained from traditional Chinese medicinal herbs of the Epimedium genus. 104 The first study that observed a promoting activity of icariin on proliferation of stem cells was carried out by Fan et al. 105 Icariin in the range 10 nM–1 μM induced the in vitro cell proliferation of hBMSCs. The same effect was observed on NSCs derived from forebrains of mice embryos. 106 Icariin promoted neurosphere formation and proliferation of NSCs in a dose-dependent manner, achieving the maximum effects at 100 nM. The authors suggested that these effects were partially mediated by the extracellular signal-regulated kinase (ERK)/MAPK. The regulatory roles of icariin in increasing cell proliferation of NSCs in vitro was also investigated in human NSCs derived from fetuses. 107 The treatment of human NSCs with 10 μM icariin led to a higher proliferation rate compared with control cells. The microarray analysis carried out in this study showed that icariin modulates the expression of different genes in the Wnt and bFGF pathways, in particular, icariin significantly enhanced the expression of FGFR1 and three important Wnt family members: CTNNB1, FZD7, DVL3, and, at the same time, reduced the expression of GSK-3β. These results were further defined and broadened by Fu et al. 108 who observed that icariin promoted the growth and proliferation of rat hippocampus NSCs in a dose-dependent manner through the regulation of the cell cycle genes, cyclin D1 and p21. Higher concentrations of icariin (20–320 μM) increased proliferation of rat BMSCs. 109 The increased rat bone marrow stem cells (rBMSC) proliferation appeared to be mediated by the phosphorylation of ERK and p38 and by the overexpression of MAPK targets, Elk-1 and c-Myc. Another study evidenced that 100 nM icariin promotes in vitro proliferation of rat adipose-derived stem cells (ASCs) through the RhoA and Rho-associated protein kinase (ROCK) signaling pathways. 110 Icariin has also been investigated in relation to the cellular damages caused by oxidative stress experienced during cryopreservation and thawing. 111 The addition of icariin to the freezing solution containing DMSO significantly increased the postthawed cell viability, decreased the apoptosis rate, improved cell adherence, and maintained the mitochondrial functions, as compared with the freezing solution containing DMSO alone. Moreover, icariin inhibited oxidative stress and upregulated heat shock proteins 70 and 90.
Icaritin [3,5,7-trihydroxy-2-(4-methoxyphenyl)-8-(3-methylbut-2-enyl)chromen-4-one], an icariin derivative, has been shown to increase proliferation and enhance self-renewal of mouse ESCs. 105,112 Icaritin cooperates and stimulates ERα, functioning as a effective inhibitor of CDX2 and p130, which successively activates cyclin E/CDK2 signaling to trigger G1/S phase progression and upregulates pluripotency markers like Klf4, Oct4, Nanog, and Sox2. The proliferative effect of different concentrations of icaritin for different times has also been investigated in MSCs. 113 Icaritin treatment increased MSC proliferation after 2 and 3 weeks of exposure with the highest MSC proliferation measured with 100 nM icaritin treatment. The study suggests that icaritin promotes the proliferation of human adult MSCs by the activation of the signal transduction activator transcription factor 3 (STAT-3), with a resulting increase of cysteine (C)-X-C motif chemokine receptor 4 (CXCR4) expression and activity.
Chrysin
Another highly investigated flavonoid in relation to the optimization of stem cell cultures in vitro is chrysin (5,7-dihydroxyflavone), a flavone predominantly present in honey, propolis, and passion fruit. 114 In a recent article, ASCs were cultured on three-dimensional nanofibrous mats loaded with ASCs. 114 Interestingly, ASCs, cultured in the presence of chrysin, exhibited improved adhesion and increased viability compared with cultures on nanofibers without chrysin or tissue culture polystyrene. Moreover, ASCs on chrysin-based nanofibers showed enhanced stemness maintenance capability evidenced by the overexpression of four stemness markers: Nanog, Sox-2, Oct-4, and Rex-1. In another study, chrysin was incorporated into biocomposite scaffolds encompassing chitosan, carboxymethyl cellulose, and nanohydroxyapatite, and mouse MSCs (mMSCs) were seeded on these scaffolds. 115 Chrysin released from scaffolds enhanced cell proliferation downregulating cyclin C1 mRNA, thus inducing the transition of cells from G0 phase to G1 phase, upregulating Cyclin B1 and E1 and promoting the progression of mMSCs from G1 phase to S and G2/M phases.
3,2′-Dihydroxyflavone
A study explored the effect of different flavonoids characterized by different hydroxylation pattern in the B ring of skeleton of diphenylpropane (C6 C3 C6) on cell growth and pluripotency marker expression in mouse embryonic stem cells and iPSCs. 116 Among the tested compounds only 3,2′-dihydroxyflavone increased cell growth and the expression of pluripotency marker Sox2, Oct4, Nanog, Cripto, and Fgf4. The mechanisms behind these effects were a strong activation (AKT and STAT3) or inhibition (ERK and GSK3β) of self-renewal-related kinase of both ESCs and iPSCs. These data suggest that 3,2′-dihydroxyflavone could be useful for the maintenance of high-quality pluripotent ESCs and iPSCs.
Morin hydrate
Morin hydrate (3,5,7,2′,4′ pentahydroxyflavone) is a polyphenol compound extracted from Prunus dulcis, Castanea sativa, Morus alba, and other fruits. 117 It has been observed that morin hydrate (1–2 mg/L) promotes growth and proliferation of isolated mouse inner ear NSCs as measure by MTT assay and BrdU incorporation. 118
Resveratrol
Resveratrol (3, 5, 4′-trihydroxystilbene), a nonflavonoid polyphenol with stilbene structure, is found in different plants such as cranberries, blueberries, grapes, peanuts, and eucalyptus. 119
The first studies that evidenced a stimulatory activity of resveratrol on stem cell proliferation were conducted on human bone marrow-derived mesenchymal stem cells (hBMSCs). 120,121 In particular, 10 nM to 10 μM resveratrol induced mouse bone marrow stem cell (mBMSCs) cell proliferation measured by [ 3 H]-thymidine incorporation through the NO/cGMP signal 120 and stimulates hBMSCs proliferation through an estrogen receptor-dependent mechanism and coupling to ERK1/2 phosphorylation. 121 To gain better understanding of the effect of this polyphenols on MSCs, Peltz et al. 122 investigated the effect of short-term (14 days) versus long-term (28–70 days) exposure to resveratrol at different concentrations (0.1–10 μM) in hMSCs. Resveratrol showed a dose-dependent activity on hMSCs self-renewal as a result of its combinatorial effect on cell doubling time, and on cell senescence and cell proliferation rate. At 0.1 μM, resveratrol showed the highest benefit, enhancing stem cell self-renewal over both short- and long-term exposure, meanwhile 5 or 10 μM treatment inhibited cell self-renewal. In accordance with these observations, the expression of genes involved in cell survival (Sirtuin 1, Sirtuin 2, Birc4, and Birc5) was increased by a lower resveratrol concentration but inhibited by a higher resveratrol concentration. A study conducted on mouse ESCs showed that nM concentrations of resveratrol (50 and 500 nM) restored the pluripotency of ESCs differentiated with retinoic acid by modulating the JAK-STAT3 signaling pathway which, in turn, stimulates the transcription of the stemness key factors, OCT4 and SOX2. 123 In addition, the same resveratrol concentrations enhanced ESC proliferation through the induction of genes related to proliferation, such as Cyclin A, Cyclin D, and Pcna. Of note, the authors observed that higher concentrations of resveratrol played a nonsignificant role in the regulation of ESC self-renewal suggesting that the timing and concentration of resveratrol are fundamental in eliciting its final activity on self-renewal of ESCs. These aspects were further investigated by Safaeinejad et al. 124 who indicated that 50 μM resveratrol might induce cell proliferation of hESCs by the modulation of the cell cycle. Interestingly, this resveratrol concentration did not negatively affect pluripotent markers and upregulated antiapoptotic markers. The authors suggested that resveratrol maintained self-renewal of hESCs at least in part through “SIRT1-MEK/ERK” axis.
Daucosterol
Daucosterol (β-sitosterol β-
Salvianolic acid B
Salvianolic acid B is a hydrophilic compound present in Salvia miltiorrhiza Bunge, commonly known as Danshen and widely used for the treatment of various diseases, such as cardiovascular disease, Alzheimer's, hyperlipidemia, and acute cerebrovascular disease. 126 Salvianolic acid B significantly increased the proliferation of rat neural stem/precursor cells (NSPCs) by activating the PI3K/Akt signal pathway and maintained NSPC's self-renewal by increasing the expression of two self-renewal markers, Nestin and Notch-1. 127
Oleuropein
Oleuropein is a phenolic compound present in olive oil with high antioxidant capacity. 128 Human adipose-derived MSCs showed a higher proliferative rate in the presence of 10–100 μM oleuropein as measured by WST assay. 129
Curcumin
Curcumin is a polyphenol isolated from the rhizomes of the plant Curcuma longa (turmeric) and is a commonly used spice throughout the world. 130 This natural antioxidant has been investigated in relation to the proliferation of multipotent neural progenitor cells (C17.2 cells). 131 Low doses (0.1 and 0.5 μM) of curcumin induced C17.2 cell proliferation, whereas high doses (≥10 μM) triggered a cytotoxic effect. The authors demonstrated that the mechanisms by which curcumin elicits this effect is by activating ERK and p38 MAP kinases.
Astaxanthin
Astaxanthin is a xanthophyll carotenoid ubiquitous in nature, especially in the marine environment, and is found in high amounts in algae, and in aquatic animals such as salmon, trout, krill, and lobster. 132 It has been shown that astaxanthin possesses a particularly strong antioxidant capacity compared with other carotenoids. 133 Treatment with astaxanthin (5–10 ng/mL) increased proliferation and colony formation of mouse neural progenitor cells in a dose/time-dependent manner through the activation of the PI3K and MEK signaling pathways. 134 Moreover, astaxanthin upregulated transcription factors associated to proliferation, such as CDK1, CDK2, and Rex1, and increased the expression of stemness genes (OCT4, Nanog, SOX2, and KLF4) suggesting that astaxanthin can also improve stem cell potency.
In conclusion, different phytochemicals showed a promising activity in increasing stem cell proliferation and promoting stemness maintenance suggesting their use in ex vivo expansion of stem cells.
Plant extracts
Not only purified phytochemicals, but also extracts from officinal and not officinal plants have been demonstrated to influence stem cell expansion and stemness maintenance.
Lippia origanoides
Lippia origanoides is an aromatic plant widely found throughout the Brazilian Amazon. Its essential oil has antioxidant activity, thanks to the presence of the monoterpenoids, thymol, p-cymene, and myrcene, and the sesquiterpene, caryophyllene. 135 The supplementation of the essential oil from L. origanoides (1 μg/mL) in the culture medium of stem cells isolated from human adipose tissue induced a strong increase of cell proliferation especially at passage 3. 136 The authors suggested that this effect could be attributed to thymol, the principal constituent of the oil of L. origanoides used in the study (88.2%).
Cirsium vulgare
Cirsium vulgare is a common plant belonging to Asteraceae family whose main components are flavonoids and phenolic acids. 137 The effects of a C. vulgare hydroethanolic extract (200–1000 μg/mL) and its main component, di-(2-ethylhexyl) phthalate (100–600 μM) have been investigated on the proliferation and pluripotency of neonatal rat hippocampus-derived NSCs. Both the extract and di-(2-ethylhexyl) phthalate at the higher concentrations were able to increase proliferation and the phthalate upregulated the expression of the stemness marker Sox2. 138
Hibiscus sabdariffa
Hibiscus sabdariffa is a plant rich in flavonoids whose biological effects seem to be mediated by its antioxidant activity. 139,140 The supplementation of a water extract of H. sabdariffa to murine bone marrow-derived hematopoietic stem cells showed a cell-genoprotective potential and modulated proliferation and intracellular antioxidant status, without causing a remarkable loss of surface marker expression for HSC. 141
Drynaria fortune
Huang et al. 142 studied the proliferative effect of a commercial flavonoid extract of Drynaria fortune, a common type of traditional Chinese herb, on stem cells isolated from rat dental pulp. The extract (0.01–0.1 g/L) induced dental pulp stem cell proliferation in a dose-dependent manner by increasing the number of stem cells entering into S phase dose dependently.
Other extracts
Epimedium flavonoids, extracted from a traditional Chinese Epimedium herb, promoted the proliferation of rat NSCs. 143 Unfortunately, the authors did not characterize the extract they used.
Silymarin, a combination of flavonolignans obtained from Silybum marianum, has been shown to have a strong antioxidant activity. 144 Silymarin (50 and 100 μM) increased cell viability of MSCs from human bone marrow by elevating intracellular GSH. 145
Combination of purified phytochemicals and/or plant extracts
One of the first study that took into account the possibility to use a combination of natural antioxidant compounds to stimulate the proliferation of stem cells was conducted by Bickford et al. 146 Human hematopoietic stem cells derived from bone marrow were supplemented with different concentrations of catechin, carnosine, blueberry extract, green tea extract, and vitamin D3 alone or in combination. All the tested compounds exhibited a positive effect on cell proliferation, although the combined treatments showed a stronger effect with respect to the single treatments. In particular, the combination of blueberry extract, green tea extract, carnosine, and vitamin D3 led to a higher proliferation rate with respect to the positive controls treated with human granulocyte colony-stimulating factor. Of all the tested compounds, blueberry extract significantly increased cell proliferation when combined with the other compounds.
Very recently, we demonstrated that a combination of epigallocatechin-3-gallate and sulforaphane exhibited a higher efficacy against oxidative stress induced by hyperoxia with respect to the single antioxidant treatments in hAFSCs. 147 In particular, the combined treatment was more effective in reducing endogenous ROS generation, enhancing GSH levels and increasing the endogenous antioxidant enzymes glutathione reductase, NAD(P)H:quinone oxidoreductase-1 (NQO1), and thioredoxin reductase (TR). Moreover, the combined treatment upregulated the pluripotency markers, Oct4 and Nanog, and downregulated the senescence-associated gene markers, p16 and β-gal.
Luo et al.
148
investigated the effect of two different combinations of antioxidants, a proprietary antioxidant supplement from Sigma-Aldrich (AOS, Catalogue Number: Sigma A1345) at 10,000-, 50,000-, and 200,000-fold dilution, and a homemade cocktail of
In conclusion, the use of different phytochemicals or plant extracts demonstrated to be effective in reducing hyperoxia-induced damage in vitro. Of note, any compounds elicited its effect on cell proliferation or stemness maintenance targeting specific molecular targets and at different concentrations in the different cell cultures (Table 1). On these bases, the use of a combination of different phytochemicals and plant extracts seems to be the most promising approach to maximize the effect.
Conclusions
Today, the potential application of stem cells is dramatically increasing, and stem cell therapy will probably be applied in numerous chronic degenerative disease therapies in the future.
So, the maintenance of high-quality stem cells in culture without any side effect in proliferation, potency, or differentiation potential will be critical for all present and future biomedical applications. The best way to isolate, culture, and maintain stem cells from primary cultures should be under their physiological conditions, that is, low oxygen concentration and low glucose. Nevertheless, this often is not possible due to economical and/or practical reasons and therefore, the possibility to use natural antioxidants compounds represents a potentially very effective alternative. The use of natural products, and above all of natural phytochemicals, opens new frontiers. Many of these compounds, generally recognized as safe by the American Food and Drug Administration, can act at different molecular and cellular levels to modulate proliferation and stemness maintenance in several stem cell types (Fig. 2). Further studies are necessary to deepen the knowledge on the wide world of natural compounds that could be used in stem cell cultures, on their mechanisms of action, and on the optimal dose and association of different phytochemicals to give an important contribution to stem cell application in regenerative medicine.
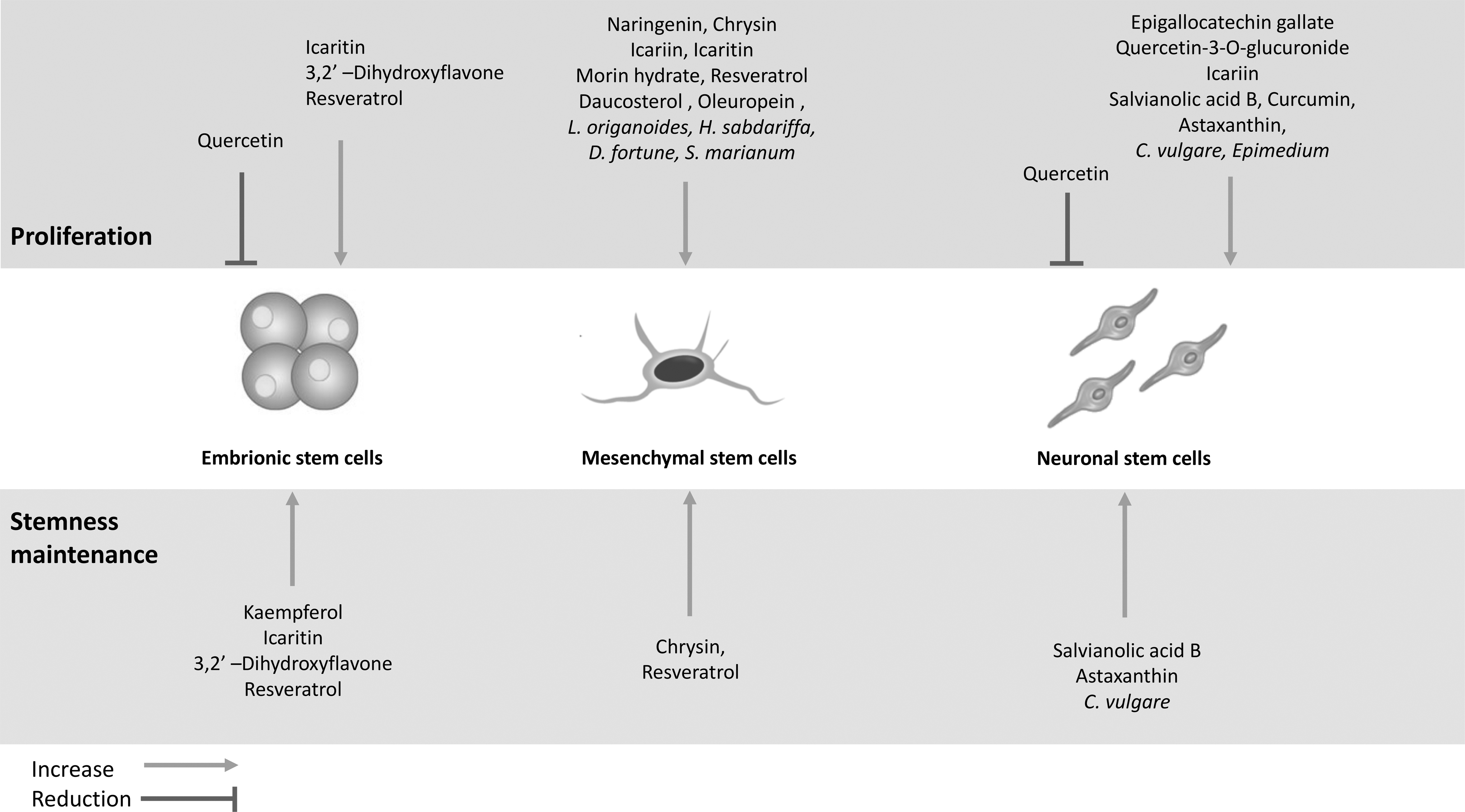
Effect of phytochemicals and plant extracts on proliferation and stemness maintenance of different stem cell types.
Footnotes
Acknowledgment
Data reported in this review were partly obtained, thanks to the support of Fondazione Cassa di Risparmio di Bologna (Italy).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.