
Abstract
The ultrastructural effects of prolonged normothermic and cold ischemia on the cerebral cortex of the adult rat were investigated. Complete cerebral ischemia was produced by cardiac arrest and the animals' temperature was maintained at 37°C for periods ranging from 0 to 81 hours before electron microscopy preparation. Electron micrographs of cold cerebral ischemia were generated after stabilizing the rat's temperature at 0°C after cardiac arrest for periods ranging from 0 hours to 6 months. A qualitative examination of the electron micrographs shows structural signatures of energy depletion such as vessel leaking and chromatin clumping after 1 hour at 37°C and after 24 hours at 0°C, followed by synapse degradation after 6 hours at 37°C and 1 week at 0°C. Evidence of advanced necrosis was observed after 36 hours at 37°C and 2 months at 0°C. A deep learning algorithm was introduced that demonstrates the temperature dependence of ischemia-induced ultrastructural changes and that can also successfully distinguish between early ischemic changes and advanced necrosis.
Introduction
The brain is one of the most metabolically active organs in the human body and is highly dependent on the continuous delivery of energy through the process of oxidative phosphorylation. For example, Hall et al., in a 2012 study, reported that “oxidative phosphorylation, not glycolysis, powers presynaptic and postsynaptic mechanisms underlying brain information processing.” 1 When oxygen delivery to the brain is interrupted, a cascade of biochemical events is set in motion that eventually leads to loss of cellular viability and autolysis. Raichle speaks of a “complex picture of interacting and cascading events that tend to reinforce the initial ischemic insult and initiate biochemical reactions that ultimately destroy vital cellular elements.” 2 The narrow limits of cerebral recovery from cardiac arrest have contributed to the popular perception that these limits reflect rapid disintegration of brain structure. Previous studies of prolonged global ischemia, however, have shown that the ultrastructure of the brain changes rather gradually after the interruption of oxygen, glucose, and other energy substrates. The processes of autolytic degradation of brain macrostructure and microstructure have been shown to occur not over a matter of minutes, but of hours and even days, and are critically dependent on temperature and the conditions under which ischemia occurs (i.e., asphyxia, shock, sepsis, and sudden cardiac arrest). 3 Better understanding the time course of the degradation of brain morphology during prolonged global ischemia under normothermic and hypothermic conditions is important for utilization of postmortem brain tissue for neuroscience research, forensic investigation, and future theoretical limits of cerebral resuscitation.
Our research has three goals: The first goal was to obtain a deeper understanding of how very long periods of cerebral ischemia without reperfusion affect the ultrastructure of the brain and how these changes are modulated by temperature. The second was to investigate whether temperature-adjusted equivalent periods of ischemia produce identical ultrastructural properties (as a simple thermodynamic model of damage would predict). A third aim was to develop and test a mathematical algorithm that can be used to estimate the duration of ischemia entirely by analyzing the ultrastructural properties of the brain (given a certain temperature). The findings are first presented and then discussed within the broader context of contemporary definitions of death.
Scientific background
The ultimate limit for resuscitation without cognitive deficits is an active field of investigation and discussion. 4 Unlike aestivating animals, 5 most mammals cannot use physiological mechanisms to induce hypometabolism in times of stress. In swine, resuscitation with good neurological recovery was possible after 13 minutes, although outcomes declined after 15–17 minutes. 6 Twenty minutes of normothermic circulatory arrest in monkeys is compatible with complete recovery of neurological function. 7 Survival of dogs without neurological deficits after 20–30 minutes of cerebral anoxia and up to 30 minutes after asanguineous normothermic total body perfusion has been reported. 8 Hossmann and Sato demonstrated recovery of the pyramidal response and evoked EEG activity from normothermic cerebral ischemia up to 60 minutes. 9 A 2019 study demonstrated the restoration and maintenance of microcirculation and cellular function of the pig brain under ex vivo normothermic conditions up to 4 hours postmortem. 10
Hypothermia increases the tolerance of the brain to ischemia. In emergency medicine and surgery, the properties of hypothermia to mitigate the damage associated with cerebral ischemia are routinely demonstrated through the use of deep hypothermic circulatory arrest in which blood circulation, respiration, and brain activity are paused for up to 40 minutes to allow advanced medical procedures. 11 In controlled conditions, a human can be cooled to as low as +9°C for a period of 1 hour of circulatory arrest with later recovery. 12 In preclinical research, dogs have been resuscitated after 3 hours of ultra-profound hypothermic circulatory arrest without blood replacement. 13 In 1966, White et al. investigated the surgically cannulated, isolated, canine brain at approximately 2°C–3°C. Brains that were stored up to 4 hours recovered electrical activity upon reperfusion and rewarming. Much more surprisingly, brains stored for 1, 2, 8, and 15 days demonstrated sustained recovery of perfusion and A-V O2 and V-A CO2 differences that were comparable with those observed in the brains stored for vastly shorter periods of time and which demonstrated recovery of EEG activity. 14 These studies demonstrate that the brain can remain viable for extended periods of time, provided temperatures are lowered to reduce metabolism. If sufficiently nontoxic solutes to prevent ice formation could be found, in principle, it should be possible to lower the temperature of the brain to a point where any diffusion-driven biochemical activity ceases, to permit indefinite storage in a viable state. In our hypothermic experiments, the temperature of the rats was maintained at 0°C because it is the lowest practical temperature at which to study cerebral ischemia without risking the ultrastructural changes associated with ice formation.
At the level of the neuron, Dutch researchers reported that, up to 8 hours after death, human brain neurons have the potential to recover function, metabolism, and axonal transport, and remain alive in culture for several weeks. 15 The survival of mitochondria isolated from human brains 8.5 hours postmortem has also been demonstrated, as indicated by the ability of these organelles to generate ATP. 16 In a 2007 histological study, a woman's brain was examined after 2 months' storage at 3°C without chemical fixation. Some macroscopically visible surface deterioration had occurred; histologically, however, normal brain structures were preserved, including all important cell types (neurons, astrocytes, oligodendrocytes, and microglia), neuropil, axons, and myelin sheaths. 17
These studies support the view that, at both the macroscopic and microscopic level, complete ischemia does not result in instantaneous and complete disappearance of brain structure. The process of brain death is a gradual one that begins with the loss of brain ion homeostasis, 18 followed by the cessation of brain metabolism, postmortem up and downregulation of genes, 19 proceeding to the loss of the viability of individual brain organelles. This ultimately leads to time-and-temperature-dependent disintegration of brain structure at the molecular level, at which point death, the irretrievable loss of life, may reasonably be said to have occurred.
Ischemia without reperfusion
Many ultrastructural studies of ischemic brains and brain tissue have been conducted with the aim of understanding the histological signatures of focal ischemia, cerebral ischemia-induced impairment of cerebral blood flow, and the effects of specific neuroprotective strategies. Relatively few studies have aimed at understanding the long-term effects of global ischemia without reperfusion of the brain, because there are currently no perceived therapeutic applications. The few studies conducted in this area have been motivated by either a purely scientific interest in the biochemistry of decomposition or by forensic concerns (i.e., deducing the time or cause of death).
Reperfusion of the ischemic brain produces regional and microcirculatory perfusion impairment (the so-called “no-reflow” phenomenon) 20 leaving some areas of the brain poorly perfused. This also affects fixative perfusion (unpublished laboratory results). Solenski et al. showed that, at 24 hours, neuronal mitochondria subjected to permanent ischemia are better preserved than mitochondria that have sustained an equivalent period of ischemia and reperfusion. 21 In a no-reperfusion study of complete cerebral ischemia in the rat, Jenkins et al. proposed that the neuronal selective vulnerability that is often reported in the literature is not observed if perfusion is not restored, and thus may be a consequence of reperfusion. 22 A study comparing the Golgi Cox and rapid Golgi staining methods in postmortem human brains reveals striking differences in structural preservation, 23 which illustrates how the processing of ischemic tissue can itself alter the tissue under observation. To address these issues in our experimental design, we only used perfusion fixation for controls and used immersion fixation for the brain-derived ischemic slices.
This study builds upon a prior 2008 article that documented the ultrastructural changes in the rat cerebral cortex after various periods of room-temperature ischemia. 24 Body temperature falls much faster in rats than in humans after circulatory arrest; to better approximate the body temperature changes that occur in humans postmortem, we maintained rats at 37°C in our normothermic circulatory arrest model. Our study further expands on this article by adding more ischemic exposure times (up to 81 hours), adding a series of cold ischemia experiments, and using deep-learning algorithms to characterize the results.
Deep learning
Another novel approach is the use of deep learning to characterize and infer the degree of ischemia from a series of electron micrographs alone. Preliminary efforts to computationally estimate the extent of autolysis in cortical, postmortem rat brain tissue as a function of normothermic ischemic exposure time were informally published in 2011. 25 A learning algorithm was developed, based on spatial frequency spectral analysis of the (grayscale) electron micrograph (EM) images, expressed as numerical arrays. The algorithm could be trained on a series of images to estimate the corresponding ischemic times. The algorithm would then, in theory, be able to estimate the ischemic time for other, not previously processed images without any previous knowledge of the samples that were imaged. In this manner, it would be able to assess the extent of damage, expressed in units of ischemic time at constant body temperature, and the correctness of its assessment would be reflected in how accurately it was able to estimate the ischemic exposure time. Expectations included gaining insight into the process of tissue deterioration under ischemia, and obtaining a means to assess the quality of any preservative method. The method achieved only very limited numerical success, mainly a very rough tracking of actual ischemic times, with better results obtained with simplified versions of the problem. Useful insight was gained in constructing a learning algorithm, which appears to be the first applied to the problem at hand. Our new approach is to use greatly improved learning algorithms based on deep neural networks (DNNs).
Materials and Methods
Animals
The animals (22 Sprague-Dawley rats, male, 90–120 days, 350–400 g) were purchased from Envigo, Inc. (Indianapolis, IN). All animals received humane care in compliance with the “Guide for the Care and Use of Laboratory Animals” (
Methods
In normothermic ischemic experiments, the animals were euthanized with isoflurane, and after cessation of heartbeat were kept at normothermic body temperature (37°C) for the duration of the ischemic period in a laboratory incubator. Rectal temperatures were monitored and recorded throughout the ischemic period using a digital logging thermometer. For electron microscopy experiments, the animals' brains were dissected 0, 1.0, 2.25, 3.86, 6.0, 9.0, 13.5, 21.0, 36.0, and 81.0 hours after cardiac arrest. (These time periods were used in the preliminary study in attempting to simplify the analysis. They are defined as the ratio of two linear functions of an “exposure index” u, with the time period t in hours given by t = 9u/(10 − u), u taking integer values from 0 through 9.)
In cold ischemic experiments, the animals were euthanized with isoflurane, and after cessation of heartbeat cooled, and then kept at ice water temperature (0°C) for the duration of the ischemic period. Animals were cooled by packing in a container of wet ice and then placed in a refrigerator. Water was drained from the bottom of the container and ice was replenished daily to maintain the animal at a constant temperature. Rectal temperatures were monitored and recorded throughout the ischemic period using a digital logging thermometer. For electron microscopy, the animals' brains were dissected at 0, 24, 48, and 72 hours, and 1 week, 2 weeks, 1 month (31 days), 2 months, and 6 months after cardiac arrest.
To approximately compare warm and cold exposure times (37°C vs. 0°C), we assumed the “Q10 rule” whereby ischemic change/damage per unit of time was reduced exponentially with a drop in temperature and declined by a factor of 2 for each 10°C reduction. 26 A drop of 37°C, from warm to cold, would then yield a reduction factor of ∼13. On this basis, then, an hour of warm time would be equivalent to 13 hours of cold time. Estimates of equivalent warm or cold times are included in Table 1, bearing in mind, that the Q10 rule is only a crude, approximate indicator of what is actually occurring. For example, it does not explain the potent neuroprotective effects of reducing brain temperature by only a few degrees after cerebral ischemia. It is possible that different elements of ischemic injury, like excitatory amino acid release, are reduced by a rate exceeding that expected by applying the Q10 rule. 27
Normothermic and Cold Ischemia Samples
Electron microscopy
After the ischemic period, cortex specimens (2 × 2 × 2 mm) were cut from the frontal lobe of the brain and fixed in 4% paraformaldehyde / 2% glutaraldehyde in 0.1 M phosphate-buffered saline (PBS) for at least 24 hours and then washed in 0.1 M PBS. Samples were postfixed with 1% osmium tetroxide in PBS for 2 hours. After postfixation, samples were treated with aqueous 0.5% uranyl acetate for 2 hours, dehydrated in a graded series of acetone solutions, and slowly infiltrated with Spurr's epoxy resin 28 over a period of 2 days. Samples were embedded in BEEM capsules and polymerized at 60°C for 36 hours. Ultrathin 70 nm sections were cut on a Leica Ultracut-R microtome using a diamond knife and sections were collected on formvar-coated copper slot grids. Sections were poststained with 2% uranyl acetate in 50% ethanol followed by Sato's lead citrate. 29 Images were generated on a Philips CM12 transmission electron microscope operated at 80 kV and micrographs were acquired with a Gatan model 791 CCD camera.
Algorithmic estimation of cortical autolysis: DNN model
Deep convolutional neural networks have emerged as the standard method for image classification. 30 They are composed of many stacked layers, each consisting of a filter set followed by a nonlinearity and acting on the output of the previous layer. Given an image as input, filters in the first layer typically extract basic structures such as edges, whereas those in deeper layers build more abstract features. Trained on a large dataset of natural images, their learned representations are transferable to a variety of image interpretation tasks. 31 An existing network consisting of 16 convolutional layers trained on natural images was used 32 as a module for generating a descriptor for any generic input image. This network outputs a descriptor in the form of a 4096-dimensional vector produced by its deepest layer.
Within this computational framework, we explored the question of the discriminability of electron microscopy images of samples subjected to different ischemic time and temperature conditions. Specifically, given a pair of experimental conditions (IA:TA, IB:TB) where IA, IB each denote either cold or warm (normothermic) ischemia and TA, TB each denote an elapsed time, we tasked the computational model with deciding whether an image belongs to experimental condition IA:TA or IB:TB. Tested on an equal number of images from each of the two conditions, the baseline model (random chance) would have an accuracy of 50%. The extent to which a trained model is capable of outperforming chance is indicative of the visual distinctiveness of neural tissue subject to the two different experimental conditions. All possible pairs of ischemic conditions were considered, and a discriminative model was trained for each pair.
To facilitate this analysis, a series of preprocessing steps was applied to all electron microscopy images. All images were cropped to exclude overlaid axes and scale indicators. They were then rescaled to a resolution of 75 pixels per micrometer, through bilinear downsampling. All original images were at no less than this resolution, so such resampling placed all data at a standard scale without introducing artifacts or blurring. To be compatible with input dimensions expected by our pretrained neural network, and to provide more examples for training pairwise classifiers, sampled 112 × 112 pixel patches centered at random locations were used in place of full size images. A new image dataset was then formed from these patches. Sampling ∼500 patches per original image, this yielded 127 K cold ischemic examples (64 K from 129 training images, 63 K from 126 test images), and 148 K warm ischemic examples (73 K from 146 training images, 75 K from 151 test images), each in the form of a 112 × 112 image patch. The sampling process ensured that patches from any original image were either all in the training set or all in the test set. As a final preprocessing measure, to remove the possibility of absolute intensity being a confounding factor in the analysis, the dynamic range of each patch was normalized by pinning its minimum and maximum intensity values to 0 and 1, respectively, and linearly rescaling. The training and testing sets were further subdivided according to ischemic duration. All preprocessing steps were implemented in MATLAB.
Pairwise classifiers were trained from an equal number of examples of each condition drawn from the training set. These example 112 × 112 patches were processed by the pretrained neural network, yielding a set of 4096-dimensional descriptor vectors. A random projection matrix reduced descriptor dimensionality to 256, and a logistic classifier was trained on the resulting 256-dimensional vectors. The same procedure was followed at test time, except with the last step replaced by application of the classifier learned during the training phase. All processing was carried out in MATLAB with use of the MatConvNet 33 and Liblinear 34 frameworks.
Classifiers using DNN features (the deep model) were also compared with a baseline model in which pairwise classifiers were learned in the same manner but restricted to use simple histogram descriptors extracted from the same 112 × 112 patches. Specifically, a histogram descriptor was formed by quantizing patch brightness into 64 intensity bins and recording the fraction of pixels belonging to each bin. This baseline was motivated by the observation that the amount of white or blank space in an EM image may be correlated with ischemic duration. The histogram descriptor was strictly more informative than counting whitespace area; the high intensity bins captured this property, whereas the other bins may capture additional characteristics.
Results
Electron microscopy
In their review article of structural damage in experimental ischemia, Van Reempts and Borgers distinguished between the neuronal, glial, and vascular parts of the brain to reflect the phenomenon that damage does not occur and evolve in a uniform manner in the brain. 35 In a more general sense, it should be recognized that ischemia-induced changes in ultrastructure have stochastic properties. When looking for ultrastructural signatures of cerebral ischemia at different time intervals, it is possible to identify qualitative trends, but not every individual organelle may exhibit those properties. For example, it is not only possible to find examples of extensive damage in individual mitochondria after 60 minutes of normothermic ischemia but also well-preserved mitochondria after 81 hours of normothermic ischemia.
We present qualitative descriptions of the progressive damage that unfolds as a function of ischemic time under normothermic and hypothermic conditions. Annotated electron micrographs for each time period are given in Supplementary Figures S1, S2, S3, S4, S5, S6, S7, S8, S9, S10, S11, S12, S13, S14, S15, S16, S17, S18.
We follow these qualitative characterizations by grouping images with different durations of ischemia to visually illustrate the progressive damage to specific structures (synapses, vessels, etc.). All individual panels contain images at the same scale (Figs. 1 –8).
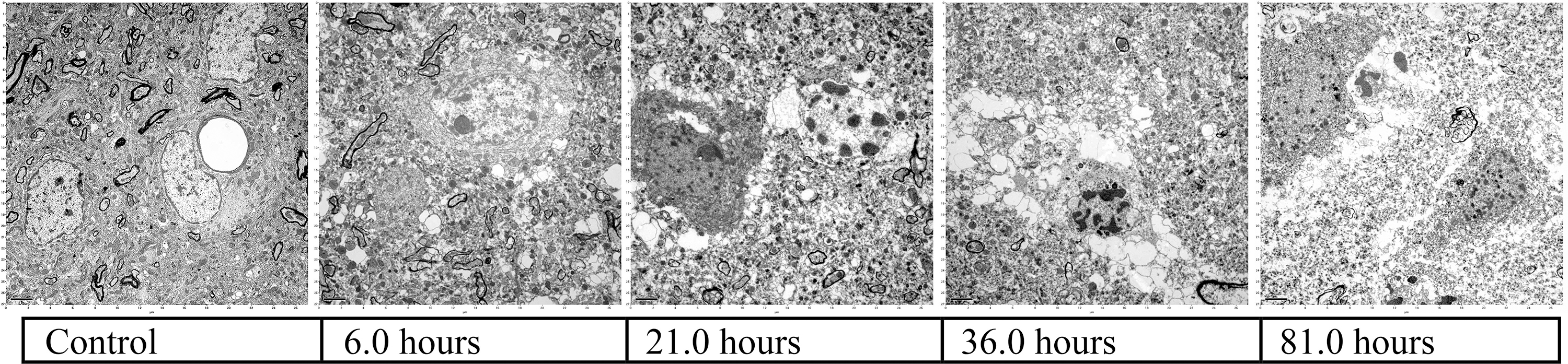
Progressive loss of fine brain structure as a function of time (normothermic ischemia).

Blood vessel integrity compromise and leaky blood vessels (normothermic ischemia).

Gradual loss of myelinated axon definition and increase in vacuolated incisures (normothermic ischemia).

Clear distinction between pre and postsynaptic structures replaced by dark, blurry, synaptic clefts (normothermic ischemia).
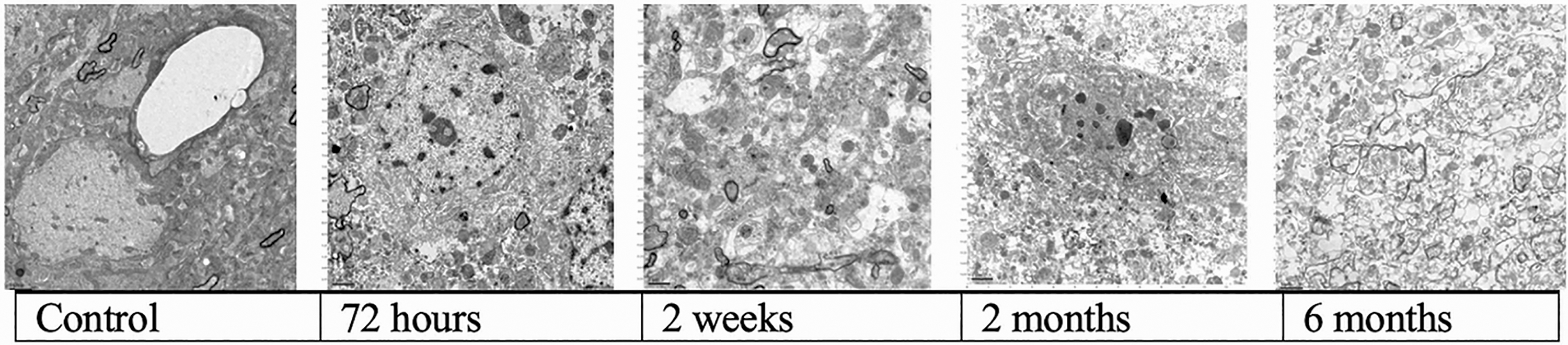
Progressive loss of fine brain structure as a function of time (cold ischemia).

Blood vessel integrity compromise and leaky blood vessels (cold ischemia).
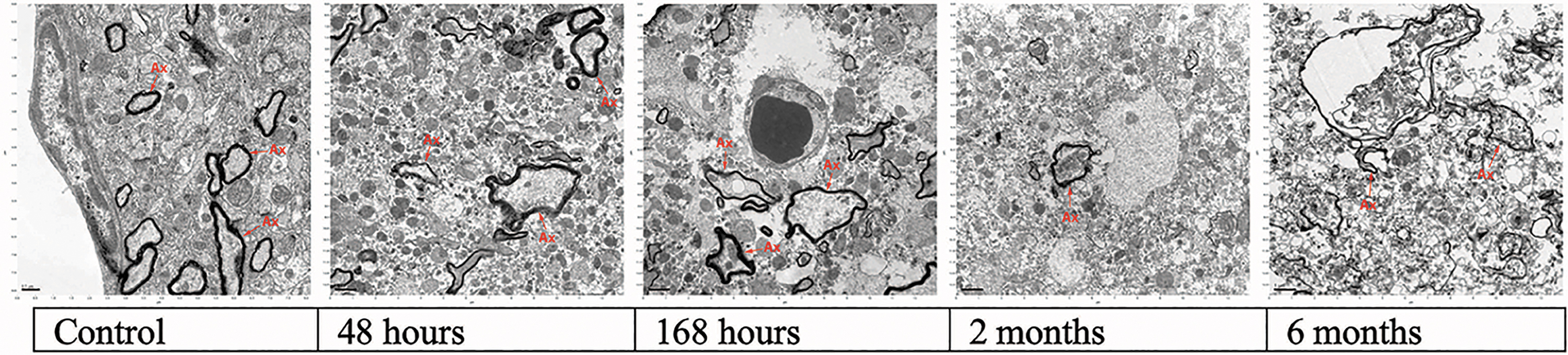
Gradual loss of myelinated axon definition and increase in vacuolated incisures (cold ischemia).

Clear distinction between pre and postsynaptic structures replaced by dark, blurry, synaptic clefts (cold ischemia).
We complete our results by presenting a table that aimed to provide an approximate time estimate for eight distinct ultrastructural changes as a function of ischemic exposure time, including two specific central nervous system (CNS) changes (synapse degradation and myelin fragmentation) (Table 2).
Timeline of Ultrastructural Changes
Normothermic Ischemia
1-Hour normothermic ischemia
Organelles were still well-defined. Internal structure of mitochondria looked intact, although swollen. Osmophilic clumping of chromatin in the nucleus with microvacuolization appeared near the endoplasmic reticulum, whereas larger vacuoles surrounded the capillaries, with areas indicative of fluid accumulation.
2.25-Hour normothermic ischemia
Organelles were still recognizable and intact. Mitochondria appeared swollen, and there was progressive loss of internal structure with some vacuoles forming. More clumping of nuclear chromatin with electron-dense materials concentrated near the nuclear membrane. Nucleoli remained visible. Distinguishing between swollen endoplasmic reticulum and vacuoles became more challenging.
3.86-Hour normothermic ischemia
There was swelling of endoplasmic reticula with large vacuoles. An increasing number of mitochondria showed distorted cristae, had vacuoles, or were missing internal structure. There was general increase in electron-dense materials and lack of identifiable nucleoli in some nuclei. Synapses were still identified. Some thinning of the myelin sheaths was seen.
6-Hour normothermic ischemia
A substantial increase in the number of empty structures, larger vacuoles, and extensive areas indicative of edema. There were disorganized and swollen endoplasmic reticula. Some mitochondria retained clear internal organization and firm membranes, whereas others looked fragmented and washed out. General membrane damage started to appear, notably affecting the nuclear membrane. It was more challenging to distinguish between mitochondria, lysosomes, and small electron-dense areas.
9-Hour normothermic ischemia
Membrane damage became more pronounced. Most mitochondria still had distinct definition, but others were damaged or ruptured. Large vacuoles in the cytoplasm and near the nuclear cell membrane started to appear. There was some unraveling of myelin sheaths.
13.5-Hour normothermic ischemia
Some images looked better than the 9.0-hour images. Many mitochondria still looked intact, but others looked washed out and showed membrane damage. The endoplasmic reticulum was no longer observed in most of the images. Myelin sheaths remained recognizable in general, but some were thinner and unraveled. A growing contrast between pale areas and electron-dense areas of the nucleus was observed.
21-Hour normothermic ischemia
There was progressive damage to membranes, including the nuclear cell membrane, leading to infiltration of organelles into the nucleus, and nuclear chromatin spilling into the cytoplasm. An increasing number of myelin sheaths lacked definition and some had exterior damage. The increase of dispersed electron-dense areas made it more difficult to distinguish between synaptic and nonsynaptic adjacent structures.
36-Hour normothermic ischemia
There was extreme electron-dense chromatin accumulation near the nuclear membranes and large vacuous spaces. In the cytoplasm only mitochondria were easily detected. Myelin sheaths were visible in some images.
81-Hour normothermic ischemia
Morphology of nuclei was detected in some images but in others there were vast areas of unstained space without recognizable structure. Fragments of cell membranes remained recognizable. There was extensive lysis of mitochondria and leaking of internal structure. Myelin sheaths were in advanced states of unraveling. Many areas appeared flooded and lacked recognizable structure.
Cold Ischemia
24-Hour cold ischemia
Organelles, nuclei, nucleoli, axons, and synapses were still recognized without difficulty. Compressed capillary lumen surrounded by edematous pale spaces, swollen mitochondria, and distended endoplasmic reticulum were observed.
48-Hour cold ischemia
Increased swelling and fragmentation of internal structure of mitochondria were observed. There was increase in electron-dense areas in the nucleus. Nucleoli were no longer observed. Synapses and axons remained visible.
72-Hour cold ischemia
Presynaptic and postsynaptic terminals remained recognizable. Increased cytoplasm fragmentation with vacuoles, pale nuclei, and organelles. There was damage to nuclear membranes. Mitochondria remained recognizable but had membrane damage.
168-Hour (1 week) cold ischemia
In many of the images, the myelin sheaths were dark and well defined. There was extensive edema around capillaries and pale nuclei with concentrated areas of chromatin. There was formation of large vacuoles out of the distended elements of the endoplasmic reticulum. Synapses were difficult to detect.
2-Week cold ischemia
Synapses were visible in some images. Washed out myelin sheaths and extensive edema around capillaries were observed. Compression of capillary lumen and cytoplasm intrusion into capillaries and pale and decomposed mitochondria were seen. Signatures of endoplasmic reticuli were difficult to find.
1-Month cold ischemia
There was a sharp decrease in recognizable myelin sheaths. Extensive formation of empty distended structures was observed. Homogeneous cytoplasm and fragmented nucleus with black electron-dense areas were seen. Many mitochondria had disintegrated and were hard to recognize. Formation of microvacuoles in axons was observed.
2-Month cold ischemia
Some images started looking “pointillist” with debris of comparable size. Some intact capillaries remained visible. Structural integrity of mitochondria, myelinated axons, and cell nucleus was variable with some mostly intact and others in advanced states of disintegration. The variation was such that some 2-month images showed more structure than the 1-month images.
6-Month cold ischemia
Thin myelin surrounding structures remained recognizable in some images. Otherwise, there was extensive decomposition of structure. Pale structures of varying sizes with vast empty spaces in between were observed. Some red blood cells looked pale and fragmented. Isolated mitochondria were seen in some images.
Table 2 demonstrates how structural changes follow the same sequence in both normothermic and cold ischemia. As can be expected, the reduced rate of chemical reactions at lower temperatures slowed the pace of these changes significantly. Near-complete disintegration of structure appeared to occur after several days at normothermic temperatures but took several months at 0°C.
The ultrastructural changes in cerebral ischemia follow roughly four stages: (1) the earliest structural changes follow depletion of metabolic support, intracellular ion regulation, and protein stability. Organelles such as mitochondria and the endoplasmic reticulum start to swell and change shape, the lumen of the capillaries is compressed by interstitial edema, electron-dense nuclear chromatin clumps, and the cytoplasm starts to swell. (2) During the second stage, the internal structure of the cell and organelles is affected. The endoplasmic reticulum further fragments, mitochondrial cristae disappear, and the nucleolus ceases to be visible. (3) This phase is followed by increasing fragmentation of the more stable cellular components; membranes rupture and distinctions between the extracellular space, cell membranes, cytoplasm, and nuclear membranes become harder to recognize. Myelin sheaths appear thinner, lose definition, and start to unravel. (4) During the final phase, there is dissolution of most recognizable biological structure with extensive white spaces. Remnants of previous structure can only be identified by looking at differences in electron density, decomposition products, and “ghost” cells.
Algorithmic estimation of cortical autolysis: DNN model
Figures 9 and 10 show examples from the image patch dataset at all combinations of temperature and ischemic duration. Patches at long ischemic duration, shown in the rightmost columns of Figures 9 and 10, exhibit visual structure distinct from those at shorter ischemic duration. From visual inspection, the amount and rate of structural change differs between warm (Fig. 9) and cold (Fig. 10) ischemic conditions. Note the different labels on the time axes; the center column of Figure 10 displays patches at longer ischemic duration than the rightmost column of Figure 9.
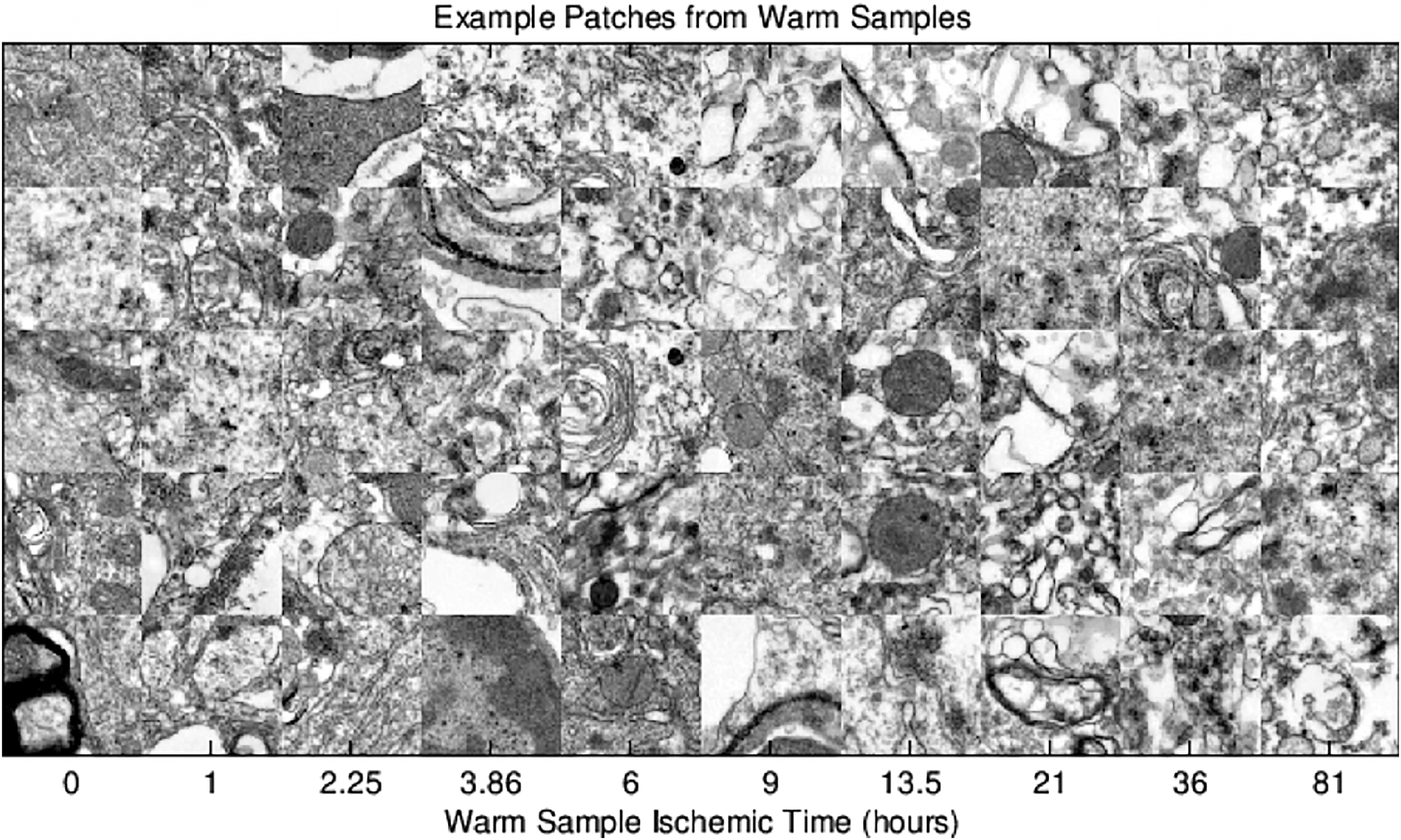
Example patches from warm samples.

Example patches from cold samples.
Figure 11 shows the test set accuracy of all the learned pairwise classifiers. The triangular sub-blocks of this result matrix report performance in distinguishing cold samples from other cold samples, or warm samples from other warm samples, while only ischemic duration is varied. Performance on these tasks shows the expected result that samples taken under the same temperature condition and nearby in time are fairly difficult to distinguish (light blue coloring on the accuracy scale). Outliers are the very long duration cold (c4464) or warm (w81) ischemic samples, which can be distinguished from most other conditions with consistently high accuracy.

Test set pairwise discrimination accuracy.
The square sub-block in the upper right of Figure 11 reports accuracy of classifiers for warm versus cold at varying ischemic times. Average accuracy is higher here than for the classifiers within the same temperature group (cold vs. cold or warm vs. warm) at varying ischemic times. However, note that the control conditions of cold at time zero versus warm at time zero are successfully distinguished with 76% accuracy, whereas one would expect chance (50%) performance here as no structural changes should take place with zero elapsed time, regardless of temperature. The explanation for this discrepancy is unclear. One possibility is a statistical difference in the content of the images collected for these controls. As the normothermic and hypothermic control images were obtained from different animals, nonpathological quantitative and qualitative ultrastructural brain differences should be expected. For example, one set of images might capture more cell nuclei or axons than another. As the classifier is free to use any available signal, including differences in content not related to structural degradation, it would do so to achieve above-chance performance. Unfortunately, checking this speculation requires annotating and cataloging the true content of each image in detail; such analysis by human experts is at present unavailable.
Although the above-chance classifier performance for warm versus cold control remains curious, there appears to be a consistent time-dependent trend for warm versus cold discrimination difficulty. Specifically, given any fixed set of cold samples (except the c4464 outlier), warm samples at longer ischemic times are easier to distinguish from them than warm samples at shorter ischemic times. Indeed, the accuracy increases with warm ischemic time, as shown in the top plot of Figure 12. The same does not hold true in the inverse scenario. The bottom plot of Figure 12 illustrates the lack of any increasing or decreasing trend in discrimination accuracy as a function of cold ischemic time. These results are consistent with the hypothesis that the warm ischemic samples undergo more rapid structural changes than the cold samples. Hence, the time dependence in Figure 12 (top) would be expected as a consequence of degradation from warm ischemia, and the lack of time dependence in Figure 12 (bottom) might be consistent with less rapid degradation from cold ischemia.
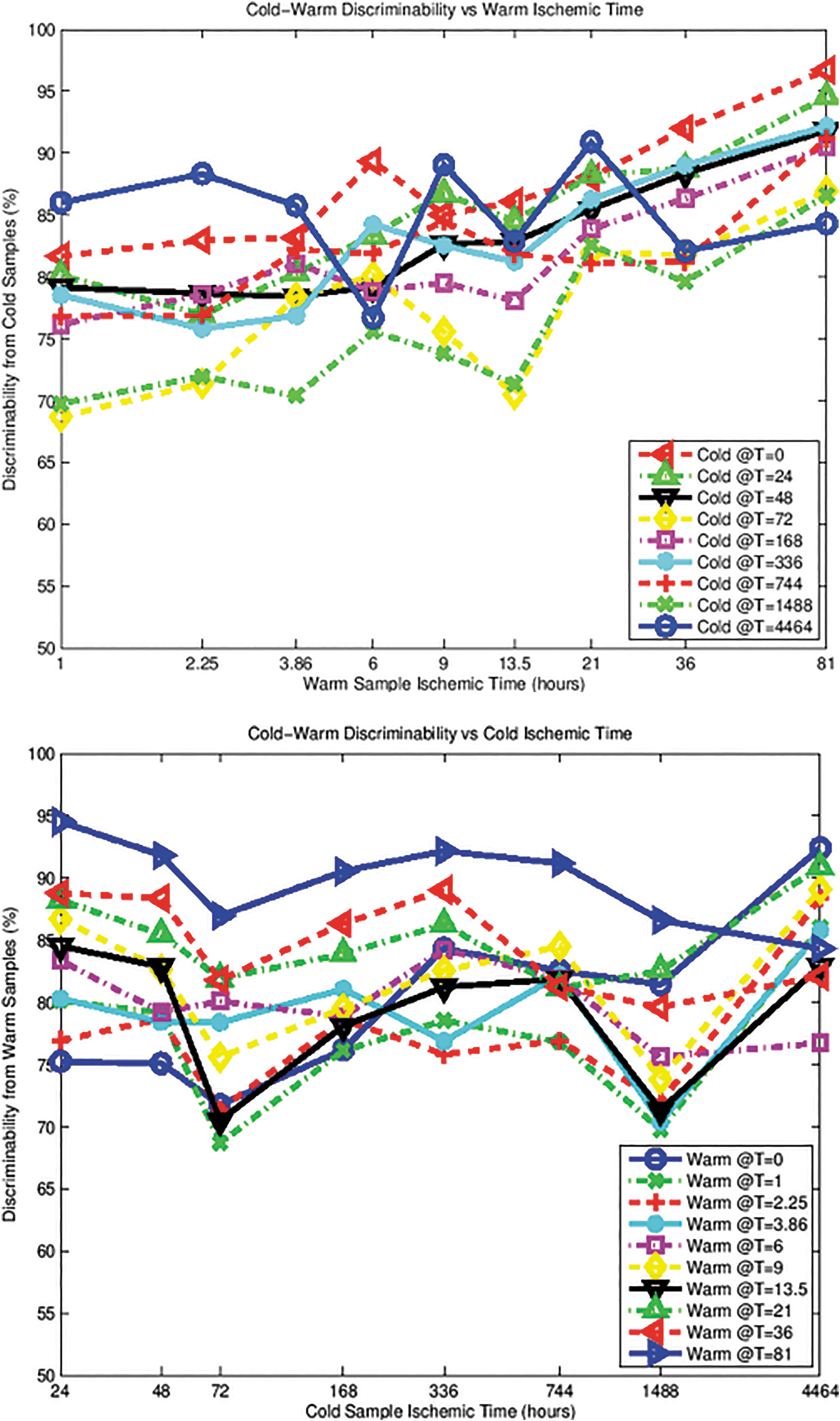
Top: Cold-warm discriminability versus warm ischemic time. Bottom: Cold-warm discriminability versus cold ischemic time.
Figures 13 and 14 compare performance of classifiers using neural network descriptors (the deep model) with a baseline model in which the pairwise classifiers use histograms as the descriptors for image patches. Figure 13 shows that the baseline model exhibits trends similar to those of the deep model in terms of relative ease of cold versus warm pairwise discrimination. However, the absolute performance of the baseline model is significantly lower than that of the deep model. Figure 14 analyzes this performance difference in detail. On average, a pairwise classifier using deep features is 7.49% more accurate than one using the baseline histogram features.

Top: Histogram Model: Cold-warm discriminability versus warm ischemic time. Bottom: Histogram Model: Cold-warm discriminability versus cold ischemic time.
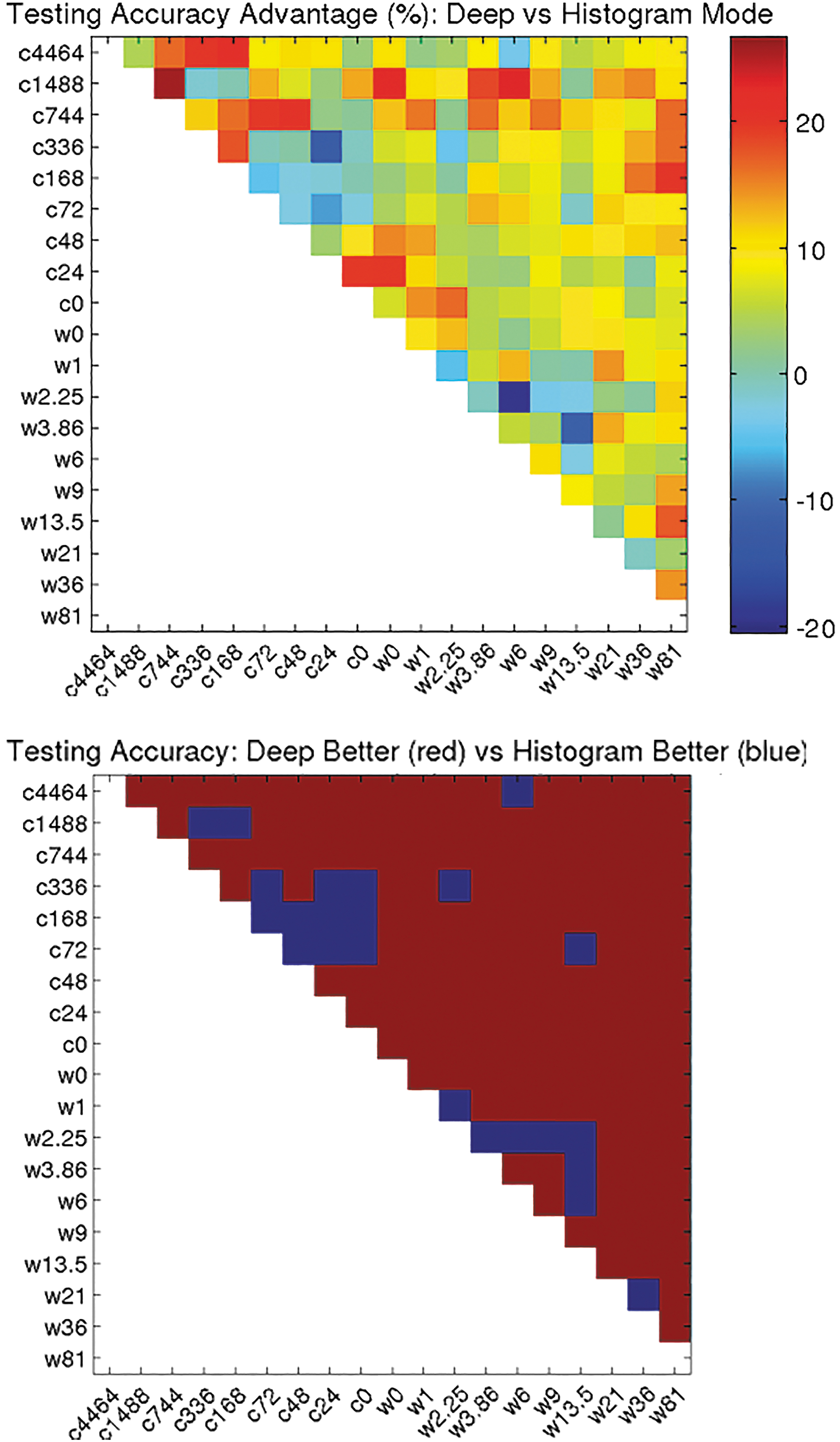
Top: Testing accuracy advantage (%): Deep versus Histogram mode. Bottom: Testing accuracy: Deep better (red) versus Histogram better (blue).
This advantage of the deep model over the baseline persists even if we exclude control conditions (ischemic time zero) from the set of classifiers compared; here the average advantage is 7.28%. Control conditions for both normothermic and cold ischemia were perfusion fixed rather than immersion fixed, which could be the cause for a systematic shift in whitespace characteristics because red blood cells have been washed out of the blood vessels. However, the fact that the deep model outperforms the baseline on average, whether controls are included or excluded, suggests that the deep features are encoding some useful information beyond a simple measurement of whitespace.
Although the results in Figures 11–13 are consistent with the hypothesis of differing rates of structural change in warm and cold ischemic conditions, they are not sufficient to rule out alternative explanations; further work on detailed human and algorithmic analysis of image content is warranted.
Discussion
There is a paucity of ultrastructural studies of permanent cerebral ischemia, and rigorous studies conducted at cold temperatures in particular. This is believed to be the first systematic study that compares the ultrastructural effects of normothermic permanent ischemia with cold permanent ischemia in the mammalian brain. Our results contribute to a greater understanding of the ultrastructural effects of prolonged cerebral ischemia without reperfusion, which further advances the study of ischemia-induced necrosis and (forensic) time of death estimations.
Examination of the electron micrographs of rat brains subjected to prolonged periods of cerebral ischemia indicate a clear difference between controls and the ischemic images. In particular, ultrastructural changes after short durations of normothermic ischemia appeared to reflect the initial, fast-paced, biochemical, and diffusion-based changes after cerebral ischemia (i.e., loss of ion homeostasis and influx of water into cells and organelles such as mitochondria). The damage to cell membranes increased with ischemic duration. Pronounced loss of structure only appeared to be present in the 36- and 81-hour normothermic ischemia images. At some point, microbial growth is expected to progress to such an extent that bacteria irreversibly erase structure as (sterile) autolysis gives way to putrefaction, but this process could not be inferred from the images.
A similar pattern was observed in the cold ischemia images. A clear difference between controls and ischemic images is followed by a rather slow degradation of the fine structure of the brain as a function of time. As a result of temperature-induced depressed metabolism, these processes proceed at a much slower pace than at normothermic temperatures. Taking this approach to its ultimate limit, in 2015, McIntyre and Fahy demonstrated that a combination of chemical fixation and cryopreservation can, in principle, preserve the ultrastructure of the mammalian brain indefinitely, in a state indistinguishable from that of controls, as verified by transmission electron microscopy. 36
Consistent with our results, one study investigating the stability and autolysis of cortical neurons in postmortem room temperature adult rats reported no evidence of “autolytic” changes during the first 6 hours of ischemia, 24 but provided no ultrastructural characterization of CNS-specific structure, such as myelinated axons or synapses. Another study revealed that when reperfusion does not follow ischemia, rat neural mitochondria exhibit loss of matrix density and pronounced swelling, but retain their shape after 24 hours of ischemia. 21 This result is consistent with our own finding that mitochondria can be recognized as distinct organelles after at least 36 hours of normothermic ischemia. Another study that looked at postmortem changes of the pyramidal neurons in the hippocampus of the rats observed disrupted, but recognizable, myelinated fibers and preserved synaptic contacts (with a decrease of synaptic vesicles) after 10 hours of room temperature ischemia. 37 Yet another study looked at postmortem protein changes in rat brain tissue kept at 23°C for up to 72 hours. The main changes were in structural proteins and enzymes, some evident as early as 6 hours with the majority at ∼48 hours. 38 Another study found that amino acids and related compounds in postmortem rat and human brains were stable for several hours at ambient temperature. 39
These studies support this investigation's conclusion that there appears to be a period of relative stability between the initial rapid, energy depletion-induced changes (most notably expressed as edema) and the global disintegration of the fine structure of the brain. During this period of relative stability, viable recovery is no longer possible, but the appearance of the original structure may still be inferred from the damaged state.
These observations are further reinforced by our deep learning algorithm. This algorithm was successful in distinguishing controls from prolonged periods of ischemia but was less so in distinguishing the intermediate ischemic exposure times. Further research may determine whether this result reflects a lack of algorithmic refinement. The algorithm's “failure,” however, is an indication that major degradation of the ultrastructure of the brain as a result of global ischemia proceeds much more slowly than the initial changes that render cells nonviable.
It is also important to emphasize that the rate of decomposition is not only a function of temperature but is also influenced by other variables, such as pH, oxygen levels, the previous critical condition of the patient (e.g., sudden cardiac arrest vs. septic shock), barbiturate administration (which can depress brain metabolism), and even factors such as the person's weight and microbiome. One study found that previous treatment of rats with the antibiotic tetracycline slowed the postmortem rate of decay. 40
The term “connectome” refers to the comprehensive set of neural connections in the brain, encoding a person's identity and memories. 41,42 The research presented in this article suggests that, in the case of normothermic and cold ischemia, the original connectome might still be inferred from the damaged state after extensive periods of ischemia because the morphological disintegration associated with these events follows a prolonged and predictable time course. Such reconstructions, which we propose to call “reconstructive connectomics” could be conducted in silico and be run in a simulation to check for accuracy.
In his article “The Scientific Limitations of Brain Death,” Robert White posits that advances in tissue engineering and a growing awareness that the brain is not a uniform organ, but rather a collection of cellular subsystems that die in a time-dependent manner, raise doubts about contemporary definitions of brain death. 43 In fact, our finding that the ultrastructure of the brain does not rapidly decompose after cardiac arrest supports the stronger, more rigorous, information-theoretic criterion of death, 44,45 in which the criterion of viability is replaced by the stronger criterion of irreversible loss of identity-critical structure.
In principle it should be possible to articulate a neuroanatomical description of personhood and conduct studies like ours to understand how this structure is altered as a function of circulatory arrest. Trying to decide whether information-theoretic death has occurred is complex because future advanced technologies could provide unanticipated means of inferring structure. Tentatively, however, we propose that, for the rat model used in this study, information-theoretic death occurs no sooner than at ∼36 hours of normothermic ischemia or 2 months of cold ischemia. Information-theoretic death is also relevant to the emerging field of human cryopreservation (i.e., cryonics) in which a deceased person is stabilized at cryogenic temperatures to permit future medical treatment at the molecular level. 46 –49
Limitations and future research
Two-dimensional characterization of the structure of cells, organelles, and two distinct CNS anatomical features of the brain have formed the focus of our studies. Scientific understanding of the exact relationship between the persistence of two-dimensional structures and the overall “wiring” of the brain is still largely unknown. For example, the ability to recognize synapses after a specific period of cerebral ischemia does not imply that these structures can be fully traced in three dimensions. Conversely, poor recognition of synapses in two-dimensional electron micrographs does not mean that these connections cannot be traced when large sets of such images are processed to generate connectomes of these brains. Extensive damage to a fragment of a connection does not mean the connection as a whole cannot be visualized or inferred.
A related and challenging problem concerns quantification. Because the number of organelles and CNS-specific structures like synapses can vary between samples, simply counting them cannot be relied upon as a measure of ischemia. Because it is impossible to subject the exact same tissue slice to different ischemic conditions, quantification of this nature is beyond our current methodologies.
Another limitation of this study concerns temperature management during the period of circulatory arrest. Although it is reasonable to assume that maintaining body temperature is a superior, more conservative approximation of the temperature decline of the human brain after circulatory arrest, a completely realistic model would aim to mimic the slow decrease of brain temperature that would occur in a typical human environment. Furthermore, in a more rigorous research design the choice of cold ischemic time exposure can be fully determined by the Q10 rule so that for each period of normothermic ischemia a roughly equivalent period of cold ischemia is chosen. This will allow for more precise testing of the Q10 rule at the histological level.
Footnotes
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by The Alcor Life Extension Foundation.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Figure S9
Supplementary Figure S10
Supplementary Figure S11
Supplementary Figure S12
Supplementary Figure S13
Supplementary Figure S14
Supplementary Figure S15
Supplementary Figure S16
Supplementary Figure S17
Supplementary Figure S18
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.