
Abstract
Telomere length (TL), sirtuin (SIRT) 1, growth differentiation factor (GDF) 11, as well as inflammaging have been related to age-related diseases. In healthy subjects, we aimed to investigate whether leukocyte TL (LTL) associated with family history of coronary heart disease (CHD), age, sex, and lifestyle, and further potential covariations between LTL, GDF11, SIRT1 and selected proinflammatory markers. In 118 healthy subjects (18–81 years, 58% females), whole blood was collected for DNA and RNA isolation and polymerase chain reaction relative quantification of LTLs and gene-expression of SIRT1, GDF11, interleukin (IL)-18, and interferon (IFN)ƴ, respectively, and serum SIRT1 and IL-18 analyses. Shorter LTLs were associated with a seven-fold higher frequency of hereditary CHD in subjects with LTLs in quartile (Q)1 compared with Q2–4 (odds ratio = 7.5, 95% confidence interval: 2.5–21.6, p < 0.001, adjusted). We also observed that LTLs in Q4 compared with Q1–3 associated with higher leukocyte expression of SIRT1 and GDF11 (p = 0.052 and p = 0.058), lower IFNƴ expression (p = 0.009), and lower circulating IL-18 levels (p = 0.027). SIRT1 and GDF11 expression were strongly intercorrelated (Spearman's rho = 0.85, p < 0.001). Overall, smoking, snus, and alcohol consumption were not associated with LTLs. The observed shorter LTLs in association with elevated expression of SIRT1 and GDF11 and dampened inflammation in hereditary CHD subjects, suggest impending risk of disease. More research are warranted to shed light on early lifestyle interventions targeting these mechanisms, to promote healthier aging in individuals with hereditary burden.
Graphical Abstract
Color images are available online.

Introduction
Age is the main risk factor for human diseases, including coronary heart disease (CHD). Thus, to strive for healthy aging is important and requires knowledge in its biology and on the variations in how people age. Telomere attrition, thought to be involved in the aging process, is the consequence of replication-mediated mechanical loss of telomere sequences per each cell cycle division. 1 Consequently, telomeres shorten with age and may act as a biomarker for biological aging. Short telomeres have been suggested to be a major risk factor for cardiovascular disease (CVD), the causality is partially accepted, but underlying mechanisms are still elusive. 2 Telomere length (TL) is influenced by multiple factors, including heritage, sex, and unhealthy lifestyle, 3 –5 thus, critical telomere attrition, even in healthy individuals, may increase disease risk. 2
Lately, several potential regulators of lifespan and TLs have been investigated, including the intracellular health-sensing mediators named sirtuins and certain growth hormones. 6 Sirtuins are nicotinamide adenine dinucleotide (NAD+)-dependent deacetylases involved in genetic regulation. Sirtuin 1 (SIRT1) in particular, are thought to sense lifestyle changes and thus impacting health status, although its role in longevity is mainly evidenced from animal models. 7 Previously, the growth differentiation factor (GDF)11 was described in mice parabiosis experiments to retard the aging process through yet unknown mechanisms 8,9 ; however, GDF11 as a youthful factor is debated. 10,11 Both molecules are thought to decline with age in humans. 12
Reduced TLs have further been shown to associate with inflammaging, that is, low-grade chronic inflammation typical for aging. 13
We have recently reported on the association between leukocyte TL (LTL), SIRT1, and GDF11 expression in blood cells and the adversative impact of proinflammatory cytokines in patients with coronary artery disease (CAD). 12 Previous studies have reported on the associations of LTL in healthy offspring of subjects with and without CVD, however, with diverging results. 14 –16
In this explorative study, we aimed to investigate potential associations between LTLs and hereditary CHD, sex, lifestyle factors (i.e., body mass index [BMI], tobacco use and alcohol consumption) in healthy subjects free of CHD. Potential associations in healthy LTLs and SIRT1, GDF11, and the selected proinflammatory cytokines interleukin (IL)-18 and interferon (IFN)ƴ were additionally explored.
Material and Methods
Study populations
Two different cohorts were used in the present study. Cohort 1 consisted of 53 healthy young individuals, age18–40 years, collected at Oslo University Hospital Ullevål, Norway, from January to March 2018. Individuals were included based on health status; free of active or chronic disease, no use of regular medication, except from contraceptives, both sexes and not pregnant. Cohort 2 consisted of 69 healthy older individuals, 51–81 years of age, both sexes, included between October 2014 and December 2015 at the Norwegian Diabetics' Center (NDC), Oslo, Norway. This cohort was identified as healthy by being controls in the DIALONG (diabetes type 1-long-term survivors with a new syndrome of late complications) study. 17 Clinical data were recorded from both cohorts and blood samples from the respective biobanks were used. Hereditary CHD was defined as having first- or second-degree relatives (including siblings, parents, and grandparents) with known CHD (myocardial infarction [MI], ischemic stroke, known atherosclerosis), when <55 years of age in men and <65 years of age in women. Smoking and snus use were categorized as never, current, ex-, or occasional user, the latter only present in Cohort 1, as was also snus use. Norwegian snus is smokeless tobacco and a dry/moist form of snuff placed in the upper lip. Self-reported alcohol intake was categorized into four groups: abstainers, 1–14, 14–21, and >21 units/week. The study was approved by our Institutional Review Board (Oslo University Hospital, Ullevål, Norway), and the Regional Committee of Medical Research Ethics in South-Eastern Norway. All subjects gave written informed consent to participate and no sex-based or race/ethnicity-based differences were present.
Laboratory methods
In fasting conditions, venous blood samples were collected at inclusion between (8.00–10:30 a.m.). Routine analyses were performed by conventional laboratory methods. Serum was prepared by centrifugation within 1 hour at 2.500g in 10 minutes and stored at −80°C until the analysis of circulating SIRT1 and IL-18. EDTA blood and PAXGene Blood RNA tubes (Pre-Analytix GmbH, Hombrechtikon, Switzerland) were collected for DNA extraction and RNA isolation, respectively. Circulating leukocytes in peripheral blood were hence the source for DNA (LTL analysis) and RNA (GDF11, SIRT1 IL-18, and IFNƴ gene expression analysis). All materials were kept frozen at −80°C until further preparation and analysis. DNA was isolated by the QIAamp DNA Blood Mini Kit (Qiagen GmbH, Hilden, Germany). Total RNA was extracted using the PAXGene Blood RNA Kit (produced by Qiagen GmbH for PreAnalytix), with an extra cleaning step (RNeasy MinElute Cleanup Kit; Qiagen), according to the manufacturer's instruction. DNA and RNA purity and quantity was tested on NanoDrop, ND-1000 (Saveen Werner, Sweden).
LTL determination
Equal amount of DNA per experiment (5 ng/μL) was used to measure relative LTL by quantitative real-time polymerase chain reaction (PCR).
18
PCR amplification was performed on the VIIa7 instrument (Applied Biosystems by Life Technologies, Foster City, CA), using telomere-specific primers (Invitrogen by Thermo Fisher Scientific, Waltham, MA)
Gene expression analysis
Equal amount of total RNA per experiment (100 ng) was reversely transcribed into complementary DNA (cDNA) by use of qScript cDNA SuperMix (Quanta Biosciences, Inc., Gaithersburg). Leukocyte expression of GDF11 (Hs00195156_m1), SIRT1 (Hs01009006-m1), IL -18 (Hs00155517_m1), and IFNƴ (Hs00989291_m1) were normalized to β-2-microglobulin (Hs99999907_m1), previously tested as a valid housekeeping gene. 19 The messenger RNA (mRNA) gene expression analyses were measured on the VIIa7 instrument, using TaqMan Universal PCR Master Mix, No AmpErase UNG, and the TaqMan assays as noted above (Applied Biosystems), as RQ (2−ΔΔCt method). 20 Gene expression analysis were successfully analyzed in all samples (n = 122), except for GDF11 (n = 121).
Determination of circulating proteins
The Enzyme-Linked Immunosorbent Assay (ELISA) Kits were used to measure serum levels of SIRT1 (Human SIRT1; LSBio LifeSpan BioSciences, lnc., Seattle), and IL-18 (Medical Biological Laboratories, Naka-ku, Nagoya, Japan). The interassay coefficients of variation were 11.6% and 8.1%, respectively, and the markers were successfully analyzed in all samples (n = 122).
Statistical analyses
Data are presented as mean (±SD), median (25, 75 percentile), or proportions, as appropriate. The Student t-test and Mann–Whitney test were used to compare continuous data with normal and skewed distribution, respectively. The Kruskal–Wallis test was used to compare skewed data across categorized groups. Proportional data were compared using chi-square test, including linear-by-linear association. Correlation analyses were performed using Spearman's rho and related multiple comparisons were adjusted for by Bonferroni correction, as appropriate. Multivariate logistic regression was used to assess the impact of hereditary CHD on LTLs, adjusted for age, sex, smoking, and additionally cohort, BMI, and LDL cholesterol, as shown from Table 2 and Supplementary Table S2, respectively. p-values ≤0.05 were considered statistically significant. Analyses were performed using the IBM SPSS statistic software, version 24.
Levels of Leukocyte Telomere Lengths (Relative Quantification) as Related to Sex, Hereditary Coronary Heart Disease, and Lifestyle Factors in the Total Population and According to the Two Cohorts
Values are median (25, 75 percentile).
Numbers with successfully analyzed LTLs were 119, therefore, the dissimilarity in numbers in each subcategory versus the other tables.
p-values ≤0.05 are bolded.
Values refer to differences in LTLs between subgroup in the total population.
Values refer to differences in LTLs between subgroups within the young and elderly cohort separately.
Values refer to differences in LTLs between the young and elderly cohort.
Due to low number in subcategories, current, quitted, and occasional smokers and snus users have been merged, as has also the number of subjects consuming 14–21 and >21 alcohol units/week.
BMI, body mass index: divided at 25 according to being overweight or not; LTL, leukocyte telomere length; RQ, relative quantification.
Results
Demographic data in the total population and separated in the two cohorts are presented in Table 1. Mean age in Cohort 1 and 2 were 29 and 63 years and the presence of female gender 66% and 55%, respectively, 58% in total. Hereditary CHD was more frequently present in the young, whereas current smokers were only present in the elderly, as was also hypertension. Elderly subjects presented with significantly higher BMI, serum LDL cholesterol, and fasting glucose levels as compared with the young.
Demographic Variables in the Total Population and Separately in Cohort 1 and Cohort 2
Values are n (%) or mean (standard deviation) if not otherwise stated.
p-values refer to difference in variable between cohorts, p-values ≤0.05 are bolded.
BMI, body mass index; CHD, coronary heart disease; Cohort 1, healthy young; Cohort 2, healthy older; HDL, high-density lipoprotein; HRT, hormone replacement therapy; HT, hypertension; LDL, low-density lipoprotein.
LTLs as related to hereditary CHD in the total population
LTLs were statistically significantly shorter in subjects with a positive family CHD history and borderline significantly shorter in the cohorts analyzed separately (Table 2). In those reporting hereditary CHD (n = 36), 16 presented with LTLs RQ values in quartile 1 (<1.14), 7 in quartile 2 (1.14 to ≤1.48), 6 in quartile 3 (1.48 to ≤1.84), and 7 in quartile 4 (>1.84), giving significant difference in LTL distribution across quartiles (p = 0.018, Kruskal–Wallis test) (Fig. 1). Univariate logistic regression analysis revealed that subjects with LTLs in quartile 1 presented with 3.9 times higher likeliness to have first-line relatives of CHD compared with those in upper quartiles (2–4) (unadjusted odds ratio [OR] = 3.94, 95% confidence interval [CI]: 1.65–9.44, p = 0.002). In multivariate logistic regression analysis adjusting for potential covariates (age, sex, smoking, cohort, LDL cholesterol, and BMI), LTLs in quartile 1 were still associated with higher risk of hereditary CHD (OR = 7.49, 95% CI: 2.54–21.60, p < 0.001).
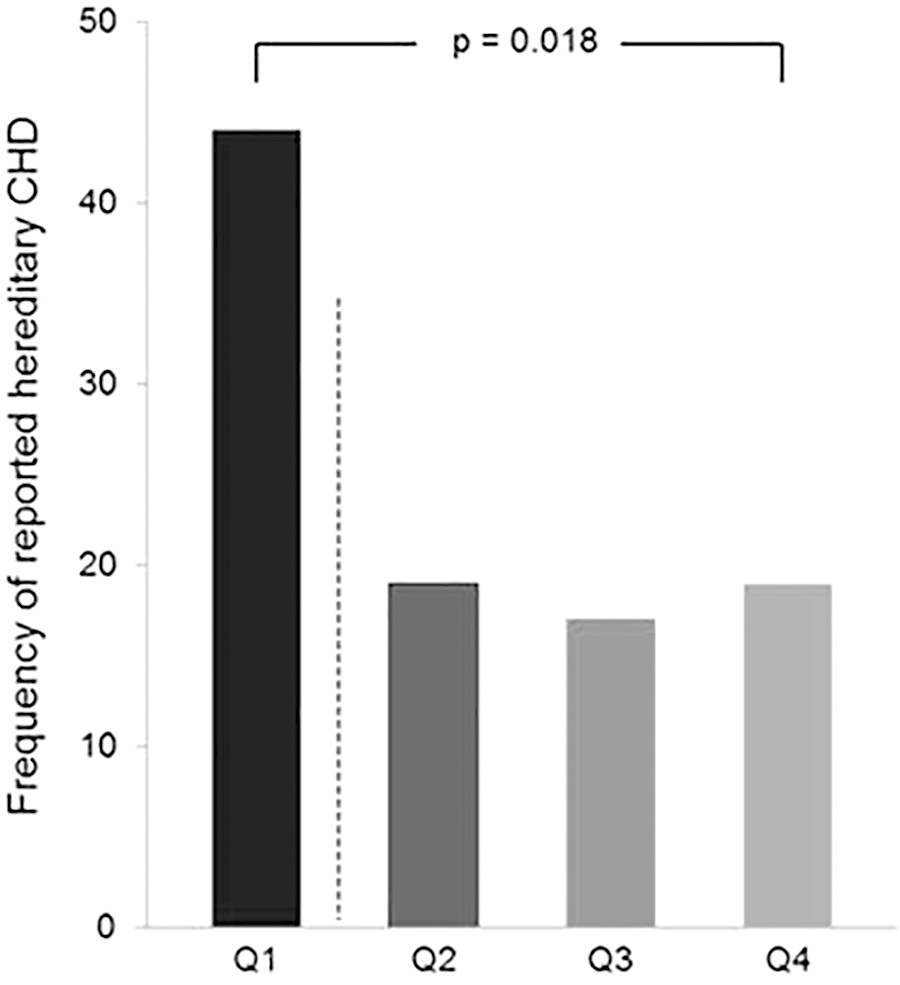
Frequency of reported hereditary CHD in quartiles (Q) of LTLs in the total population. Q1: ≤1.14, Q2: 1.14–1.48, Q3: 1.48–1.84, Q4: ≥1.84 (RQ values). Dotted line indicates the cutoff level used between Q1 and Q2–4. The p-value refers to linear-by-linear association chi-squared test. CHD, coronary heart disease; LTL, leukocyte telomere length; RQ, relative quantification.
LTLs in the two cohorts as related to sex, hereditary CHD, and lifestyle factors
The young cohort presented with statistically significantly longer LTLs than the older (p = 0.003) (Table 2). No significant difference in LTLs was observed according to sex in the total population. However, in the young cohort, male presented with numerically longer LTLs than females, whereas in the older cohort, females presented with significantly longer LTLs than males (p = 0.041). The difference between young and elderly males was also statistically significant, with the longest LTLs observed in the young male subjects (p = 0.003). LTLs were numerically longer in subjects with overweight (BMI >25 kg/m2) versus the lean, however, only significantly in the young (p = 0.024). No significant differences in LTLs were observed with respect to smoking, snus use, and alcohol consumption. In the older cohort, LTLs were also not significantly different between those with hypertension (n = 14) compared with those without: Median (25, 75 percentile): 1.67 (1.21, 1.89) versus 1.28 (1.07, 1.63), p = 0.107, or between users of hormone replacement therapy (n = 3) compared with nonusers: 1.62 (1.20, —) versus 1.29 (1.08, 1.72), p = 0.357. In the young cohort, no difference between users (n = 26) versus nonusers of contraceptives was observed [1.58 (1.30, 2.44) versus 1.34 (1.15, 2.23), p = 0.382].
LTLs as related to age
Figure 2 shows the correlations between LTLs and age with respect to sex in the total population (Fig. 2A), and in each cohort separately (Fig. 2B, C). The figure illustrates that LTLs decreased with age in both men and women in total, R 2 = 22% and 9%, respectively (Fig. 2A). When stratifying the model according to cohorts, LTLs were significantly correlated to age in the young cohort (p = 0.008), and with respect to sex, only in females (R 2 = 21%) (Fig. 2B). No statistically significant correlations between LTL and age were observed in the older cohort, independent of sex (Fig. 2C).

Correlations between LTLs and age as related to sex in the total population
Levels of SIRT1 and GDF11 according to the cohorts and related to sex, hereditary CHD, and lifestyle factors
Circulating levels of SIRT1 and gene expression in circulating leukocytes (RQ values) of SIRT1 and GDF11 with respect to categorized groups are shown in Table 3. SIRT1 and GDF11 were statistically significantly higher expressed in young versus the older cohort (p ≤ 0.001, both). Circulating SIRT1 levels were significantly higher in overweight versus lean subjects (BMI >25) (p < 0.001) and gene expression of SIRT1 and GDF11 was significantly lower (<0.02, both). Current, occasional, or former smokers presented with significantly lower expression of SIRT1 and GDF11, whereas circulating SIRT1 was significantly higher, accordingly (p < 0.04, all).
Levels of SIRT1 and GDF11 as Related to Cohorts, Sex, Hereditary Coronary Heart Disease, and Lifestyle Factors
Values are median (25, 75 percentile).
p-values refers to difference in levels of the variables between subgroups, p-values ≤0.05 are bolded.
As GDF11 expression in leukocytes was successfully analyzed in 121, result from 1 sample is lacking in the following categories: Cohort 2, females, yes for reported CHD, in BMI <25 kg/m2, quit smoker, never used, alcohol consumption between 1 and 14 units/week.
Due to low number in subcategories, current, quitted, and occasional smokers and snus users have been merged, as is also the number of subjects consuming 14–21 and >21 alcohol units/week.
BMI, body mass index: divided at 25 according to being overweight or not; GDF11, growth differentiation factor 11; SIRT1, sirtuin 1.
Circulating SIRT1 levels were significantly and differently distributed between groups of alcohol consumption. Those consuming moderate amount (1–14 units/week) presented with lowest circulating SIRT1 levels (p = 0.027) and the highest numerical GDF11 expression levels. Sex, hereditary CHD, and snus use were not associated with significant differences in levels of markers (Table 3), nor were hypertension, use of contraceptives, or hormone replacement therapy (data not shown). Sex or different cohort did not change these results noticeably, when analyzed separately (data not shown).
Correlations between LTLs and the investigated variables
Figure 3 shows the correlation between LTLs and genetically expressed SIRT1, GDF11, IL-18, and IFNƴ, as well as circulating levels of SIRT1 and IL-18. Weak inverse correlations were observed between LTLs and IFNƴ expression (r = −0.21, p = 0.022) and circulating IL-18 (r = −0.22, p = 0.017), whereas LTLs correlated borderline to SIRT1 expression (r = 0.17, p = 0.058). SIRT1 and GDF11 expression was strongly intercorrelated (r = 0.85, p < 0.001) and correlated moderately to IL-18 expression (r = 0.56 and r = 0.41, p < 0.001 in both), inversely to circulating SIRT1 (r = −0.20 and r = −0.31, p = 0.029 and p = 0.001, respectively) and to circulating IL-18 (r = −0.25 in both, p = 0.006 and p = 0.007, respectively). IFNƴ expression was inversely correlated to circulating SIRT1 (r = −0.26, 0.004) and positively correlated to circulating IL-18 (r = 0.20, p = 0.027). After Bonferroni corrections, the intercorrelation between the SIRT1 and GDF11 genes, their associations to expression of IL-18 and the inverse association between circulating SIRT1 and GDF11 expression remained statistically significant (p = 0.002 by 21 performed correlations). When stratifying according to sex, the intercorrelation between SIRT1 and GDF11 genes was stronger in women than men (r = 0.88 and r = 0.77, respectively, p < 0.001, both), as was also the genes' respective correlation to IL-18 expression in women (r = 0.70 and r = 0.54, p < 0.001, both), as compared with the weak correlation between SIRT1 and IL-18 expression (r = 0.29, p = 0.043) and lack of any association between GDF11 and IL-18 expression in men (r = 0.018, p > 0.2).
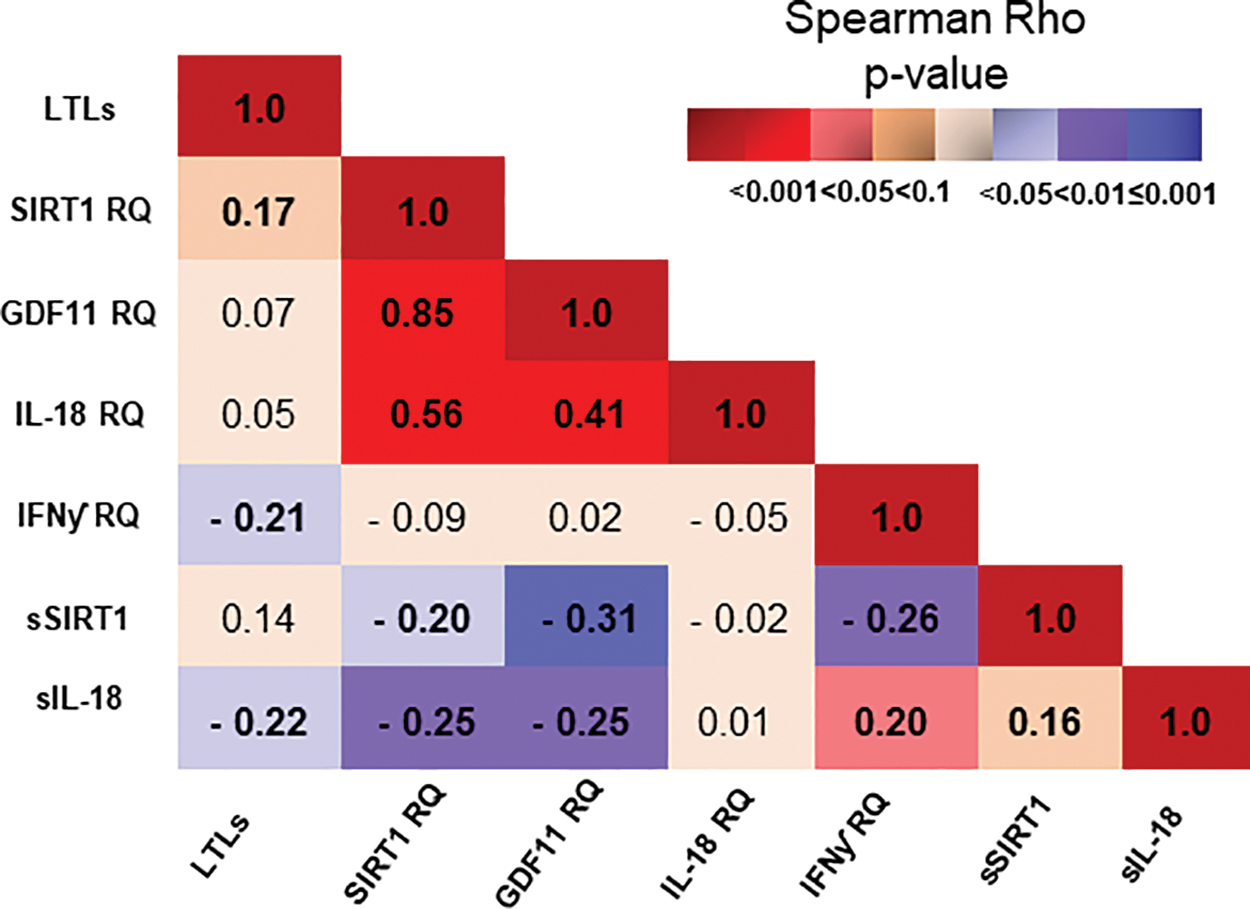
Spearman's rho coefficient of correlation between the investigated variables in the total population. r-values is specified in each box, whereas color intensity indicates strength of the p-value. Shades of red indicate positive correlations and shades of lavender indicate inverse correlations. Color images are available online.
Distribution of SIRT1, GDF11, IL-18, and IFNƴ through quartiles of LTLs
Circulating levels (SIRT1 and IL-18) and gene expression levels of SIRT1, GDF11, IL-18, and IFNƴ with respect to LTL quartiles are shown in Figure 4. With increasing quartile number (indicated with darker colors), borderline significantly lower circulating IL-18 levels (p = 0.080) and borderline significantly reduced IFNƴ expression (p = 0.067) were detected. We compared the longest LTLs versus the shorter (quartile 4 vs. quartile 1–3) and observed that LTLs in quartile 4 associated significantly with lowest levels of circulating IL-18 (p = 0.027) and genetically expressed IFNƴ (p = 0.009) and highest levels of genetically expressed SIRT1 and GDF11, the latter borderline statistically significant (p = 0.052 and p = 0.058, respectively).

Distribution of biomarkers through LTL quartiles in the total population. Red colors define circulating levels of SIRT1 and IL-18, whereas blue colors define gene expression of SIRT1, GDF11, IL-18, and IFNƴ (RQ values). Increasing darkness of color indicates increasing quartile number of LTL. Values are median (25, 75 percentiles). p 1 indicates difference between quartiles (Kruskal–Wallis test); p 2 indicates difference between quartile 4 and quartile 1–3. p-values ≤0.05 are bolded. GDF11, growth differentiation factor 11; IFNƴ, interferon ƴ; IL-18, interleukin-18; s, serum; SIRT1, sirtuin 1. Color images are available online.
Discussion
The main findings in this study were that healthy relatives of CHD patients presented with significantly shorter LTLs compared with nonhereditary exposed subjects, with a seven-fold adjusted higher risk of heredity if LTLs were in lowest versus the three highest LTL quartiles. LTLs decreased with age exclusively in young females, and women >51 years of age presented with significant longer LTLs than in age-comparable men. The leukocyte expression of SIRT1 and GDF11 was strongly intercorrelated and mirrored inversely circulating levels of SIRT1, with respect to BMI, cigarette smoking, and partly alcohol consumption. Finally, individuals with the longest versus shorter LTLs presented with higher SIRT1 and GDF11 expression and an attenuated proinflammatory profile, shown by lower circulating IL-18 and IFNƴ mRNA expression levels.
We and others have recently reported on the association between MI and LTLs in patients with CAD, suggested to reflect an advanced stage of disease and LTLs to be a marker of ongoing atherosclerosis leading to CVD. 12,21 In the present study we observed that healthy subjects, already in the 20s, have shorter LTLs if their first- and second-line relatives are CHD patients and partly before exposure of long-term cardiovascular risk factors. Our results are in line with two previous reports, but in contrast to the larger Asklepios study by De Meyer et al. 14 –16 Nevertheless, our results strengthen the hypothesis that heritage and “epigenetic” inheritance of gamete telomeres are strong drivers of LTLs, and that LTL at birth is a major determinant of LTL throughout the human lifespan. LTLs may thus not merely be a marker of ongoing degenerative processes associated with aging, but an independent marker of increased disease risk, including CHD. If healthy individuals with short LTLs and positive CHD heritage are exposed to cardiovascular risk factors, which lead to extended telomere attrition, their risk for disease is most likely strongly elevated. On the contrary, individuals with long LTLs, despite hereditary CHD, may potentially have minor risk of developing CHD.
Decreased TL associated with increased age in young females, whereas telomere attrition declined with age, especially in men, confirming previous observations. 12,22 Age explained 21% of TL variation in young females, whereas the association in young males was less obvious. Hence, TL regulation seems to be sex related, with different telomere attrition rate throughout life, as also indicated by others. 23 TL was not significantly different between sexes, however, observed to be numerically shorter in young women versus men and significantly longer in elderly women versus their age-comparable men, also reported by others. 24 Our observations may indicate males' TL to be longer than females' at birth, thereafter declining in both sexes at different rates.
The investigated lifestyle factors (overweight, smoking, and alcohol intake) were not associated with TLs, as also partly reported by others. 25 LTLs and BMI have been reported to be inversely related in children and adults, 23,26 and shown to be strongly associated with adiposity. 26 Our observed association between LTL and BMI >25 in young subjects, may merely indicate higher muscle mass. TLs and cigarette smoking has been reported to be inversely associated, possibly through oxidative stress mechanisms and alcohol consumption seems to have minimal influence on LTL, except for in chronic abuse, addiction, or binge drinking, 27,28 partly in line with our observation.
SIRT1 was observed elevated in serum of overweights, accompanied with lower genetically expressed SIRT1 and GDF11, somewhat in contrast to the report by Mariani et al., 29 but partly in line with other observations. 30 SIRT1 has been reported to be downregulated in cells with high insulin resistance, 31 often present in individuals with high BMI, which is a plausible explanation of our observation. Higher circulating SIRT1 levels were observed in the joint group of smokers and in moderate-to-heavy alcohol consumers versus the nonexposed groups partly accompanied with lower leukocyte SIRT1 and GDF11 expression. Whether elevation of circulating SIRT1 reflects an attempt of counteracting an unhealthy lifestyle and its downregulation in leukocytes indicates leukocytes as sensors of circulating levels is not clear, but SIRT1 and GDF11 expression levels were inversely correlated to circulating SIRT1, as well as to circulating IL-18.
SIRT1 and GDF11 expression levels were not associated with hereditary CHD, but suggested by others to be preventive of CVD. 6 We observed that SIRT1 and GDF11 expression levels were highly intercorrelated, confirming our previous findings, 12 which suggest common upstream regulators and/or common intracellular pathways in regulating cell senescence. As SIRT1 and GDF11 inhibit nuclear factor (NF)-κB, the main transcriptional regulator of genes related to inflammation, 13,32 and SIRT1 suppresses activation of the nucleotide-binding oligomerization domain-like receptor family pyrin domain-containing 3 (NLRP3) inflammasome, 33 their reported declined expression with age may partly be one of the mechanisms leading to chronic low-grade inflammation. Circulating SIRT1 was also inversely correlated to IFNƴ expression, however, whether this influences LTLs is unsure, although higher SIRT1 expression combined with lower IFNƴ expression and attenuated circulating IL-18 in the longest versus shorter LTLs indicates this possibility.
Limitations
Although the study has an exploratory character, results need to be taken with caution. The numbers in some subgroups are low and presence of statistical type II error is conceivable, impending that the negative associations between TLs and smoking, alcohol consumption, and partly BMI may be false negatives. Although LTL seems to be highly correlated to TL in other cells and tissues and is widely used as a measure of TLs, it is continuously debated. 2 A strength of our study is inclusion of healthy subjects at different ages and with limited use of medication.
Conclusion
In this observational study on healthy subjects, the presence of hereditary CHD was associated with shorter LTLs, substantiating the suggested link between herited short TL and disease. We could demonstrate that telomeres shorten in an age- and sex-related fluctuating manner. The inflammatory markers, IFNƴ and IL-18, were more associated with TLs than the strongly intercorrelated longevity factors SIRT1 and GDF11. Whether LTLs have value as a biomarker for age-related diseases, warrants further investigation. However, early lifestyle intervention that dampens inflammation and eventually induces SIRT1 and GDF11 expression may potentially prevent disease, potentially accentuating healthier aging.
Footnotes
Acknowledgments
Sissel Åkra and Jeanette Konstanse Steen are acknowledged for blood sampling and processing for biobanks of both study cohorts.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Norwegian Council for Cardiovascular Diseases and Stein Erik Hagen Foundation for Clinical Heart Research, Oslo Norway. Theses funding sources were not involved in the research and preparation of the article.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.