
Abstract
Astaxanthin (AST), a xanthophyll belonging to the family of carotenoids, is a potent antioxidant. The effect of AST on longevity and its physiological and molecular mechanism are still unclear. In this study, we proved that AST could prolong the life span of Caenorhabditis elegans. To uncover whether AST could delay aging by upregulating autophagy, we measured the expression of autophagy gene and the life span of autophagy gene bec-1 mutant nematodes, and the results showed that the expression of autophagy gene was upregulated after AST intervention and the disruption of bec-1 weakened the extension of the life span. To explore the molecular mechanism of AST-induced autophagy upregulation, we knocked out the daf-16 or hlh-30 (key genes of insulin/insulin growth factor-1 [IGF-1] signal pathway or target of rapamycin [TOR] signal pathway) by RNA interference, and the expression of autophagy gene lgg-1 decreased. Collectively, our results strongly suggest that autophagy, which is both the insulin/IGF-1 signal pathway dependent and TOR signal pathway dependent, plays a role in the prolongation of the life span of Caenorhabditis elegans by AST.
Introduction
Astaxanthin (AST) is an antioxidant carotenoid produced by marine animals, 1 which has gained attention in combating aging. 2 –5 Current studies have shown that AST could extend the life span of Caenorhabditis elegans through its antioxidant properties. 6,7 A recent review reported that AST could modulate signaling pathways that regulate autophagy. 8 As is known to all, autophagy is a process of cytoplasmic catabolism that protects cells from stress conditions. 9,10 The autophagy degradation ability of cells also decreases with age, which may be one of the causes of aging. Studies have proved that mutation inactivation of autophagy gene accelerates the senescence of nematode tissues, 11 and upregulation of autophagy in a suitable range could prolong the life of animals. In C. elegans, we focused on the bec-1 and lgg-1 genes, which are homologous to the yeast and mammalian autophagy genes APG6/VPS30/beclin1 and APG8/AUT7p/lc3, respectively. 12,13
Studies demonstrated that the autophagy gene cascade functions downstream of insulin/IGF-1 signaling (IIS) and target of rapamycin (TOR) signaling pathways. 11 The IIS pathway is a nutrition sensing signal system, which is evolutionarily conservative in higher organisms. 14 The main genetic factors of the IIS pathway in C. elegans are the daf-2 gene, encoding an insulin and insulin growth factor-1 (IGF-1) receptor, and the daf-16 gene, a forkhead (FOXO) transcription factor. Researchers observed that the autophagy genes are required for the long life span observed in daf-2 (IGF-1 receptor) mutants, 12,15 suggesting that IIS pathway may be required for autophagy induction in C. elegans. TOR signaling pathway is evolutionarily conserved as an important participant in cell metabolism, autophagy, and aging. 16 –20 HLH-30, which is a member of a group of 42 HLH transcription factors in C. elegans and has the highest homology to TFEB, 21 has a role in autophagy regulation in C. elegans through TOR. 22 Upregulation of the transcription factor HLH-30/TFEB induced autophagic activity and thus increased longevity. 22 Therefore, whether AST induces autophagy through these two pathways remains to be further explored.
We used C. elegans as a model to study aging due to its short life span, rapid generation, and well-defined genetic pathways. 23 –26 The purpose of our study was to examine whether AST could extend life span through inducing autophagy and to explore the mechanism.
Materials and Methods
Reagents, strains, and growth condition
AST (96%, solarbio, Tianjin, China) was dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO), and added to nematodes growth medium (NGM) and the OP50 bacterial feeding suspension. A final DMSO concentration of 0.2% (v/v) was maintained in AST containing plates and control, and this DMSO concentration does not have any effects on C. elegans locomotion. 27 About 12.5 mg/L fluorodeoxyuridine was used as reproductive suppressant. Heat-killed OP50 (30 minutes at 65°C) was utilized to prevent AST metabolized by Escherichia coli. 28 Strains used in this study were Bristol N2 wild-type (WT); VC517, bec-1(ok691); DA2123, GFP::LGG (adIs2122). All strains as well as the E. coli OP50 were obtained from Caenorhabditis Genetics Center (University of Minnesota). RNAi bacterial strains containing targeting genes are a gift from professor Zou Chenggang of Yunnan University. This type of study does not require any IRB approval or waiver. Age synchronous cultures were obtained by treating the adult hermaphrodites with a lysis solution (5 M NaOH and 5% NaOCl); the eggs collected were incubated in M9 buffer to L1 larvae, and then grown to L4 larvae in new NGM plates.
Life span assay
All life span assays were performed at 20°C. The age-synchronized nematodes were transferred to NGM plates with or without various concentrations of AST, and it was defined as day 0. To ensure that AST remained effective throughout the experiment, worms were transferred to fresh plates every other day until all worms had died. Life span assays were repeated at least three times independently.
Assessment of motility
The age-synchronized WT worms were bred on NGM plates with or without various concentrations of AST (0, 60, 120, and 240 μM), and it was defined as day 0. On the 2nd, 4th, 6th, 8th, and 10th day of adulthood, 10 worms were randomly selected from each group to measure body bends for 1 minute as previously described. 29 A body bend was defined as a change in the reciprocating motion of bending at the midbody. 30 The assays were carried out three times independently.
Pumping rate assay
Pumping rate of 10 worms from experimental groups and control group was counted for 1 minute under a microscope on the days 3, 6, and 9 of the intervention. Worms were treated with AST as described in the life span assays. The assays were carried out three times independently.
Autophagy quantification
Autophagic events were observed under a fluorescence microscope (IX81; Olympus) on the day 6 of the intervention. Worms DA2123, GFP::LGG-1 (adIs2122) were mounted with a drop of sodium azide (NaN3) placed on a cover slip covered with 2% agarose. Approximately 10 worms were observed in each group, and the assays were repeated for three independent trails.
RNA interference
E. coli strain HT115(DE3) expressing dsRNA was grown 12 hours in LB broth containing 100 μg/mL ampicillin and 12.5 μg/mL tetracycline at 37°C with shaking, then spread onto NGM plates containing 100 μg/mL ampicillin and 1 mM isopropyl 1-thio-β-D-galactopyranoside (IPTG). Synchronized L1 larvae were transferred to the plates at 20°C before spawning period. Young adult animals were used for further experiments.
RNA isolation and quantitative real-time PCR
Nematodes were cultured in NGM with or without AST, and the worms were collected with M9 buffer on the day 6 of the intervention, total RNA was isolated by using Trizol reagent. RNA purity was checked using UV absorbance (260/280 ratio). RNA was reverted to cDNA by using SPARK script II RT Kit (Spark Jade, China). Real-time polymerase chain reactions (PCRs) were performed with 20 μL of qPCR Master Mix (Promega), in triplicate for each sample. PCRs were carried out in a Real-Time PCR Machine Roche Light Cycler 480 under the conditions of 95°C for 5 minutes for PCR initial heat activation, followed by 40 cycles of 95°C for 10 seconds for denaturation, and 60°C for 30 seconds for combined annealing/extension. All experiments were repeated for three independent trails. The gene expression data were analyzed using the comparative 2-ΔΔCt method with act-1 as the normalizer.
31
The quantitative real-time PCR (qRT-PCR) primers were as follows: bec-1, 5′-ACGAGCTTCATTCGCTGGAA-3′ (F) 5′-TTCGTGATGTTGTACGCCGA -3′ (R); lgg-1, 5′-GCCGAAGGAGACAAGATCCG-3′ (F) 5′-GGTCCTGGTAGAGTTGTCCC -3′ (R); daf-2, 5′-CGGTGCGAAGAGAGGATATT-3′ (F) 5′-TACAGAGGTCGCCGTTACTG-3′ (R); daf-16, 5′-CCAGACGGAAGGCTTAAACT-3′ (F) 5′-ATTCGCATGAAACGAGAATG-3′ (R); let-363, 5′-CACAGGCGAACTAATACCGT-3′ (F) 5′-TTTCCGAGTTGCTTGATGAT-3′ (R); hlh-30, 5′-TCAGACAGGTCGTCTCAAGTTCG-3′ (F) 5′-ATCTTCGTCGGCGTTCAATC (R); act-1, 5′-CCAGGAATTGCTGATCGTATGCAGAA-3′ (F) 5′-TGGAGAGGGAAGCGAGGATAGA-3′ (R).
Western blot
The worms were collected with M9 buffer on the day 6 of the intervention. Protein was extracted by homogenization using a sonicator; protein concentrations were determined by a bicinchoninic acid (BCA) Protein Assay kit (Beyotime). Equal amounts of proteins were separated by SDS-PAGE and transferred onto PVDF membranes (PVDF; Millipore, Billerica, MA), and blocked with 5% BSA (Sigma) in 1 × Tris buffered saline Tween for 1 hour at room temperature. Subsequently, the membranes were incubated with rabbit anti-GFP (1:1000; Abcam, Cambridge, MA) and rabbit anti-β-actin (1:1000; CST) primary antibodies overnight at 4°C. They were then incubated with the secondary antibodies (horseradish peroxidase [HRP]-linked antirabbit IgG; 1:1000; CST) for 1 hour at room temperature. Then, the proteins were detected by chemiluminescence reagents (Millipore) and observed using a ChemiDocTM XRS+ Imaging System (Bio-Rad, Hercules, CA). The protein levels were quantified by densitometry using NIH ImageJ 1.61 Software (National Institutes of Health, Bethesda, MD).
Statistical analyses
The survival curves were plotted by the Kaplan–Meier survival analysis, and the significance differences were analyzed by log-rank. Statistical analysis between the two groups was conducted using Student's t-test. Three or more groups were tested by one-way analysis of variance, and then by the Student–Newman–Keuls post hoc test. p < 0.05 was regarded as significant difference. The results were expressed by mean ± SD or mean ± SEM. All the statistical analysis was conducted with SPSS 20.0 software, and the mapping was conducted with prism 7.
Results
AST extends the life span of C. elegans and improves motor performance
To investigate whether AST could extend the life span of C. elegans, we treated WT worms in NGM at 20°C with concentrations of 0, 60, 120, and 240 μM AST. Compared with the control group, life span was extended by 9.71%, 13.36%, and 5.76% in worms treated with 60, 120, and 240 μM AST, respectively (Fig. 1A and Table 1). The motility of nematodes also reflects their senescence, 32 so we measured the effects of AST at different concentrations on the motility of nematodes on the days 2, 4, 6, 8, and 10 of the intervention. The results showed that the motor performance of the experimental groups was better than that of the control group, and the group containing 120 μM AST showed better motor performance than other groups. In summary, we found that AST could extend the life span of C. elegans and improve motor performance of C. elegans. Based on results, we identified 120 μM AST as the optimal dosage.
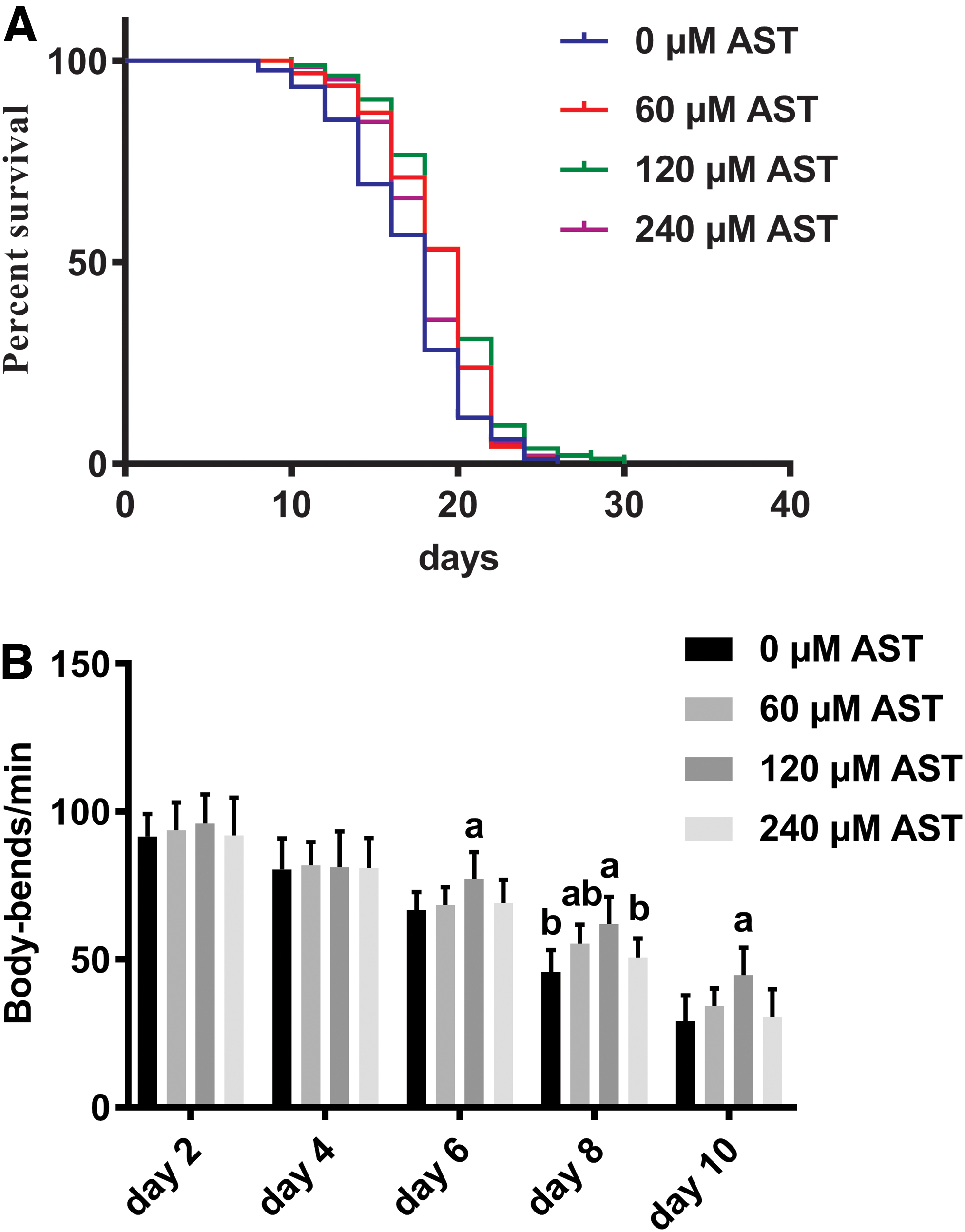
AST extends the life span of Caenorhabditis elegans and improves motor performance. Three days after hatching, age-synchronized, WT worms were freshly plated with 75 μM fluorodeoxyuridine. The NGM contains 0, 60, 120, and 240 μM AST.
Effect of Astaxanthin on Mean and Maximum Life Span of Wild-Type Worms Under Normal Condition
Results on mean life span are indicated as means ± SEM from three independent experiments.
Percentage relative to the control.
Compared with N2 (120 μM AST), p < 0.05, performed by pairwise comparisons.
Compared with N2 (control, 0 μM AST), p < 0.05, performed by pairwise comparisons.
Statistical significance of the difference between the survival curves was demonstrated by log-rank test using the Kaplan–Meier survival analysis.
AST, astaxanthin.
AST does not impact the food intake in C. elegans
Calorie restriction is also a classic way to slow the aging process. 33 We investigated whether AST affected food intake by detecting the pumping rate. The results demonstrated that there was no significant difference between the experimental groups and the control group on the days 3, 6, and 9 of the intervention (p > 0.05) (Fig. 2). Therefore, AST does not impact the food intake in C. elegans.

Pumping rate/min of WT worms on the days 3, 6, and 9 of the intervention. There was no significant difference between the experimental group and the control group. Data are presented as the mean ± SD from three independent experiments, n = 10 per group, performed by Student's t-test.
AST extends the life span of C. elegans by inducing autophagy
To uncover whether AST could delay aging by upregulating autophagy, qRT-PCR analysis was used to examine transcriptional expression of genes involved in autophagy such as bec-1 and lgg-1. Compared with the control, it was noted that bec-1 and lgg-1 mRNA levels increased ∼26.7% and 27.5%, respectively (Fig. 3A). This induction in autophagy was further evaluated by quantifying the level of GFP::LGG-1, which is expressed throughout all tissues of c. elegans. Results show that autophagy was induced, and the GFP::LGG-1 labeled spot-like structure and overall expression level were increased (Fig. 3B, C). Subsequently, to explore the effects of autophagy on longevity, bec-1(ok691) worms were treated with 0 or 120 μM AST (Fig. 3D). The difference of the life span between bec-1 + 0 μM AST and bec-1 + 120 μM AST was statistically significant (p < 0.05), but this result does not suggest that AST's role in extending the life span of nematodes is independent of autophagy, because bec-1 is not the only gene in nematode cells that can regulate autophagy. Merely suppressing the expression of bec-1 gene does not reverse the prolongation of life span by AST. We included part of the data in Table 1 for comparison. By comparing group WT +120 μM AST and group bec-1 + 120 μM AST, we found that after the autophagy gene bec-1 mutation, the life span of nematodes was shortened, and the difference was statistically significant, which showed that AST could extend the life span of nematodes by inducing autophagy (Table 2).

AST extends the life span of C. elegans by upregulating autophagy.
Life Span of Control and daf-16-RNAi or hlh-30-RNAi Worms Treated with 0, 120 μM Astaxanthin
Results on mean life span are indicated as means ± SEM from three independent experiments.
Compared with WT +0 μM AST, p < 0.05, performed by log-rank test using the Kaplan–Meier survival analysis.
Compared with WT +120 μM AST, p < 0.05, performed by log-rank test using the Kaplan–Meier survival analysis.
WT, wild type.
AST induces autophagy through IIS and TOR signaling pathway
We used qRT-PCR to detect the expression of key genes in IIS pathway and TOR signaling pathway after intervention. Results showed that the expressions of daf-16 and hlh-30 increased, while the expression of daf-2 and let-363 (the gene encoding CeTOR 34 ) decreased (Fig. 4A). We hypothesized that AST inhibited the IIS pathway (TOR signaling pathways) by downregulating daf-2 (let-363), thereby activating transcription factors DAF-16 (HLH-30). Thus, we used RNA interference to knock out daf-16 and hlh-30 in the DA2123, GFP::LGG-1 (adIs2122), respectively. The GFP::LGG-1 expression of group daf-16-RNAi +0 μM AST and group hlh-30-RNAi +0 μM AST is lower than that of group Control +0 μM AST, suggesting that RNAi successfully reduced the expression of daf-16 and hlh-30 genes. By comparing GFP::LGG-1 expression between group Control +0 μM AST and Control +120 μM AST, we inferred that after intervention with AST, the level of autophagy in c. elegans was upregulated. The GFP::LGG-1 expression of group daf-16-RNAi +120 μM AST and group hlh-30-RNAi +120 μM AST is lower than that of group Control +120 μM AST, suggesting that AST can induce autophagy by activating daf-16 and hlh-30.
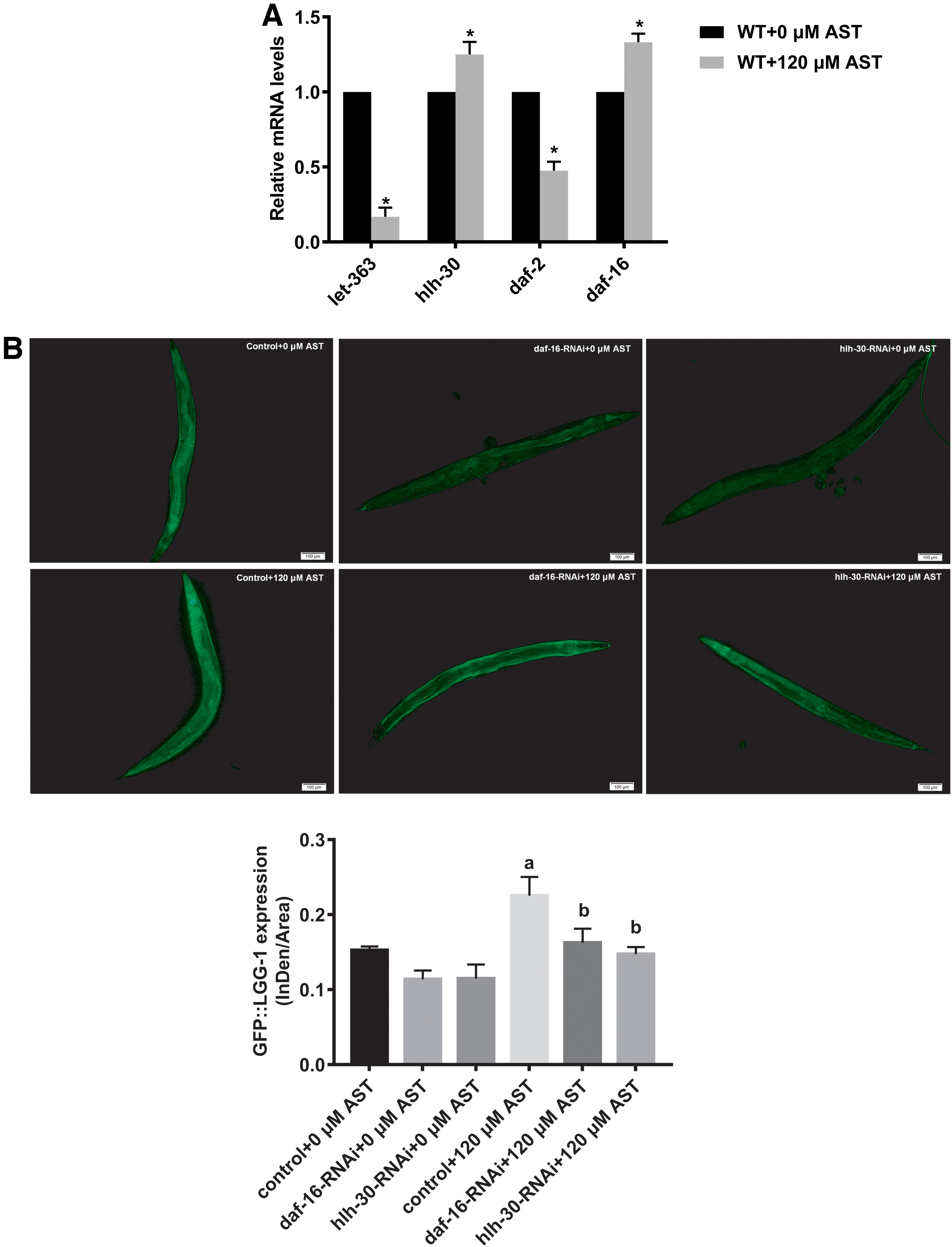
AST upregulates autophagy through activating daf-16 and hlh-30.
Discussion
Autophagy is a cell cycle process that has emerged as a crucial mechanism for life span extension in many different species. 12,35 Many studies have shown that proper upregulation of autophagy could slow down aging and lengthen life span. 12,36 –39 Our experimental results showed that both IIS pathway and TOR signaling pathway could regulate autophagy. According to the previous report, mTOR deficiency does not further increase the extended life span of daf-2 mutants, 40 indicating that TOR interacts with the insulin/IGF-1 hormonal system to control life span. 40,41 Indeed, daf-15, which encodes a Raptor-like (regulatory associated protein of mammalian TOR) protein known to function with TOR in a complex, 42 was shown to be regulated transcriptionally by daf-16. 40,41 Unfortunately, our study did not further discuss whether the IIS and TOR signaling pathways interact in inducing autophagy, and we will explore this further in future studies.
IIS pathway is an evolutionarily conserved central regulatory pathway similar to human aging process. It plays an important role in the regulation of longevity of nematodes and mammals by regulating the conserved transcription factor DAF-16, 14,43 and the activation of daf-16 leads to increased longevity, stress resistance, and the induction of heat-shock genes such as old-1, sod-3, and small heat-shock-related protein chaperones. 44 –47 Koumei Yazaki et al. and Xiaojuan Liu et al. found that AST could keep the optimal intracellular ROS balance by modulating genes involved in IIS pathway and the oxidoreductase system. 6,7 Given our experimental results, we conclude that AST not only regulates levels of intracellular ROS through the IIS pathway to extend life span but also induces autophagy through the IIS pathway to extend life span. Recent studies have shown that ROS could initiate autophagosome formation and autophagic degradation acting as cellular signaling molecules, 48 and autophagy, in contrast, serves to reduce oxidative damage and intracellular ROS levels through removal of protein aggregates and damaged organelles such as mitochondria. 49 Whether intracellular ROS and autophagy interact with each other in the process of AST extending the life span of worms remains unclear, and further studies are needed.
Conclusion
In summary, our study demonstrated that AST activates autophagy through the IIS and TOR signaling pathways to prolong the life span of C. elegans. This study suggested that AST had the potential to delay aging, and AST longevity pathway could be conserved in other organisms, including humans. However, the interaction between IIS and TOR signaling pathways, as well as between ROS and autophagy, needs to be further explored in the life-extending effects of AST.
Footnotes
Acknowledgments
The authors thank Dr. Chenggang Zou from the Yunnan University for distribution of RNAi bacterial strains.
Author Disclosure Statement
The authors declare no conflicts of interest relevant to this study.
Funding Information
This research was financially supported by Nutrition Science Research Foundation of By-Health (TY0181104).