
Abstract
Healthspan science aims to add healthy, functional years to human life. Many different methods of improving healthspan have been investigated, chiefly focusing on just one aspect of an organism's health such as survival. Studies in Drosophila melanogaster have demonstrated that a reversal to a long-abandoned ancestral diet results in improved functional health, particularly at later ages. Meanwhile, pharmaceutical studies have demonstrated that botanical extracts have potent antiaging properties, capable of extending the mean lifespan of D. melanogaster by up to 25%, without a decrease in early fecundity. In this study, we combine these two different approaches to healthspan extension to examine whether a combination of such treatments results in a synergistic or antagonistic effect on Drosophila healthspan. One botanical extract, derived from Rhodiola rosea, mimicked the effects of the ancestral apple diet with better performance at later ages compared with the control. Another extract, derived from Rosa damascena, decreased age-specific survivorship when combined with the apple diet providing support for the “Poisoned Chalice” hypothesis that combinations of various supplements or diets can elicit adverse physiological responses. More experiments in model organisms should be completed researching the effects of combining healthspan-extending substances in various diet backgrounds.
Introduction
Antiaging intervention studies aim to reverse or delay the effects of aging by targeting and manipulating the multiple biological pathways that cause this complex phenomenon. 1 Historically, studies of such interventions have focused solely on lifespan extension, without considering the trade-offs that may affect other aspects of health. 2 For example, although green tea supplementation has been shown in the past to increase mean lifespan of male fruit flies by 16%, recent studies reveal that it also impairs their reproductive fitness. 3 These supplements have minimal use for human application if the proposed lifespan extension results in impaired health. Thus, there has been a call to shift the focus of antiaging studies from lifespan extension to healthspan improvement. 4,5 Healthspan offers a more holistic measure of an organism's health, encompassing not only lifespan but also other physiological functions that contribute to an organism's state of health. These may include, but are not limited to, reproductive fitness, locomotor activity, metabolic activity, and cognitive function. By evaluating healthspan, antiaging scientists can be sure that life extension therapies help organisms live both longer and healthier lives.
A current argument in healthspan improvement is that humans should revert to the consumption of a “Paleolithic diet.” 6 More specifically, it is theorized that because human ancestors consumed a hunter/gatherer diet for over a million years, our genome has been selected for optimal survival and reproductive fitness when consuming animal tissues and natural plants, leaving modern-day humans inadequately adapted to the current agricultural diet incorporating wheat, dairy, and processed foods. 7 This diet, they suggest, has resulted in a decline in health and an increased incidence of chronic diseases such as cardiovascular disease. 8 Some suggest that a reversal to the Paleolithic diet would improve the current state of health by nourishing the body properly with food that the human genome carefully adapted to through years of evolutionary selection. In fact, some findings have already proven the health benefits of reverting to this hunter/gatherer-type diet, reporting improved blood pressure, decreased low-density lipoprotein cholesterol, and other health benefits in patients consuming the Paleolithic diet. 9 –11
Recently, a study using Drosophila melanogaster populations has revealed that a reversal to an ancestral diet results in healthspan improvement, but in an age-specific manner. 12 In this study, laboratory fly populations were fed two different diets: banana food and apple food. These laboratory flies had lived on banana–molasses food for over 30 years since they were collected from the wild (∼1000 generations). Before laboratory domestication, these flies consumed rotting apples in an apple orchard in the northeastern United States for centuries. 13 Thus, the laboratory banana food constitutes their evolutionarily recent diet, and laboratory apple food crudely represents their ancestral, long-standing diet. Healthspan was measured using age-specific assays of female survivorship probability (px ) and fecundity (mx ), which when multiplied together (pxmx ) provides a healthspan curve over all adult ages (see Materials and Methods section). 14 The results showed that pxmx remained approximately equal in the banana- and apple-fed flies at early ages (days ∼10–25 from egg) with some evidence of a decrease in age-specific survivorship in the first 3 days of adult life among apple-fed flies. At later ages (days ∼25–40 from egg), the apple group began to show superior pxmx . These data indicate that the benefits of reverting to a long-abandoned ancestral diet may become manifest only at later ages.
This finding was not unexpected. Our intuitive understanding of adaptation by natural selection is dominated by the power of selection at early ages in large outbred populations. 15 But, as the forces of natural selection fall with adult age, we expect adaptation to decline with age. This suggests that populations should adapt to a novel environment quickly at early ages, but slowly and incompletely at later ages. 16 This attenuation in the forces of natural selection should result in populations being better adapted to an abandoned ancestral diet at later ages. However, populations should be able to achieve reasonable health on a more recently imposed, although sufficiently longstanding evolutionary, diet at younger ages. Whether populations are better off consuming a recently imposed diet, or a long-abandoned ancestral diet at early ages remains unclear.
Meanwhile, research with botanical extracts has demonstrated lifespan improvement without adversely affecting other healthspan measurements. Two botanical extracts derived from Rhodiola rosea and Rosa damascena have been shown to extend mean lifespan by 25% and decrease mortality by 22%, respectively. 17,18 Both botanical extracts extended life without causing significant impairment to physiological functions, including fecundity. Although fecundity was not measured throughout the entire fly lifespan, 10-day dose/response assays were conducted for both botanicals to investigate gross impairment of reproduction. Over a course of 10 days, various doses of R. rosea and R. damascena did not significantly affect fecundity in female flies. 17,18 However, these data did not demonstrate whether later-age fecundity in Drosophila is affected by the botanicals.
The studies performed in both the Rose laboratory and the Jafari laboratory yielded complementary results; healthspan was improved through increased later-age fecundity with an ancestral diet, and healthspan was also improved through increased lifespan with botanicals. 12,17,18 This study will evaluate the effects of combining both these antiaging techniques. In addition, we will study the effects of combining multiple botanicals on healthspan. This second idea was inspired by a meta-analysis of randomized controlled trials using multivitamin/multimineral supplementation. 19 Numerous studies using humans and mice have not found combined supplementation to be useful for preventing diseases and decreasing mortality. 20 –22 In fact, Spindler et al. found that some complex nutraceutical formulations decreased lifespan in mice. 22 More recently, Jenkins et al. performed a meta-analysis in humans and found that antioxidant mixtures and niacin resulted in an increase in all-cause mortality. 23 Rutledge and Rose developed the hypothesis of a “Poisoned Chalice Effect” to explain the lack of success in these studies, suggesting that excessive multisupplementation could interfere with an organism's complex network of biochemical reactions built through evolutionary refinement of positive and negative feedback controls. 14 Thus, individual substances that alone improve healthspan may have a negative impact on healthspan when combined by undermining the complex physiological balance which underlies healthspan.
In this study, we will investigate in D. melanogaster the effectiveness of R. damascena in two diet backgrounds researched extensively by Rutledge 12 in a large-scale, well-replicated, full-lifespan study. If a long-abandoned diet improves later-age fecundity, and if the botanical R. damascena increases lifespan without negatively impacting healthspan, then a combination of these two treatments might result in improved healthspan more than is observed from either treatment individually. But if that combination does not, there is evidence for a Poisoned Chalice Effect. Additionally, if R. damascena and R. rosea both increase lifespan separately, then combining these botanicals might result in improved lifespan more than observed from either treatment alone, or again we would have evidence for a Poisoned Chalice Effect.
Materials and Methods
Study system
This study was completed in flies and is not subject to IRB or IACUC approval.
This study used large, outbred populations of D. melanogaster selected for accelerated development. 24 Experiments using outbred populations may be more applicable to natural or human populations as opposed to those using inbred populations. In addition, inbreeding has been shown to destroy genetic variation and results in deterioration of fitness-related traits, such as viability, fecundity, and male mating success. 25 Five “ACO” replicates (ACO1–5) have been reared on banana–molasses food for ∼1000 generations and have a 10-day life cycle. The wild Drosophila population from which these laboratory populations were derived is that of Northeastern United States; the local agricultural setting is one that has featured apples as the chief cultivated fruit for centuries. 13 A more detailed and up-to-date description of the history and culture methods for these lines can be found in Burke et al. 26
Overall experimental design
Two experiments were performed that monitored time-to-death (mortality) and 24-hour fecundity. In one experiment, ACO1–5 were exposed to four treatments: banana food with R. damascena supplementation, banana food without R. damascena supplementation, apple food with R. damascena supplementation, and apple food without R. damascena supplementation. In another experiment, ACO1–5 were exposed to four different treatments: banana food (control), banana food with R. damascena supplementation, banana food with R. rosea supplementation, and banana food with the combination of both R. damascena and R. rosea. This second experiment was performed with the banana–molasses diet only and will be referred to as the “combination experiment.” The design for these two experiments is summarized in Supplementary Table S1.
Food preparation
ACO populations were reared on a banana–molasses diet for stock maintenance and for select experimental assays. The banana–molasses media are composed of the following ingredients per 1 L distilled H20: 13.5 g Apex® Drosophila agar type II; 121 g peeled, ripe banana; 10.8 mL light Karo® corn syrup; 10.8 mL dark Karo corn syrup; 16.1 mL Eden® organic barley malt syrup; 32.3 g Red Star® active dry yeast; 2.1 g Sigma-Aldrich® Methyl 4-hydroxybenzoate (antifungal); and 42.5 mL 190 proof EtOH. The apple medium is prepared in the same manner as the banana medium, except the diet lacks the barley malt, corn syrups, and we substitute 1 to 1 Trader Joes® organic unsweetened apple sauce for the peeled banana.
Supplement administration
Populations were administered extracts as described by Jafari et al. 17,18 The R. rosea (SHR-5) root extract of 1.42% salidroside and 3% rosavins, as characterized by high-performance liquid chromatography (HPLC), was administered in 3% yeast solution at a 25 mg/mL dosage. The R. damascena petal extract, prepared and obtained from Dr. Asghar Zarban, was composed of 0.330% myricetin, 0.040% gallic acid, 0.051% quercetin, and 0.003% kaempferol as characterized by HPLC. 27 This extract was administered in 3% yeast solution at a 2 mg/mL dosage. Both concentrations have been experimentally reported by Jafari et al. as the ideal biological dosage. 17,18 The combination treatment of R. rosea and R. damascena was a combined dose of 25 and 2 mg/mL, respectively. Each food plate received 1 mL of yeast solution, and each cage received fresh food and supplement every 24 hours. Supplement was not administered during the developmental stage (days 1–9 from egg).
Mortality and fecundity assays
Populations were initially reared in 8-dram polystyrene vials with ∼6 mL of either banana food or apple food at ∼70 eggs per vial and given 9 days to develop. On the 10th day, flies were transferred to acrylic cages using light carbon dioxide (CO2) anesthetic and given fresh food with the respective treatment. Individual mortality was assessed every 24 hours, the flies were sexed at death, and the observed cohort size was calculated from the complete recorded deaths. During the assay, flies were transferred to clean cages once a week using light CO2 anesthesia. Transferring was performed to prevent the buildup of feces, which made assessing mortality difficult and may have subjected flies to higher levels of ammonia. All ACO1 treatments were transferred on Mondays, ACO2 treatments on Tuesdays, so on and so forth. Cohorts were assayed in 6-L cages at ∼1000 flies per cage. Flies were transferred to 3-L cages at 50% starting cohort size to control for density effects. Age-specific fecundity was also assessed every 24 hours. This parameter was estimated from the number of eggs laid by females on the culture medium plates placed in each mortality assay cage, divided by the number of females still alive. Media plates were washed on filter paper with the laboratory's fecundity funnel system and then scanned for counting at a later time.
25
Egg counting was performed using ImageJ (
pxmx statistical analysis
The age-specific survival probability (px) is the probability of a female surviving to age x, given that she survived to the start of the age interval. It is calculated using the following equation:
where dx is the number of females that die at age x, and nx is the number of females that were alive at the start of age x. Age-specific fecundity (mx ) is the average number of eggs laid per surviving female at age x. The product of these two variables gives an estimate of how cohorts are functioning at each age. In our experiments, the unit interval for x is a single day. We will refer to this parameter as age-dependent fitness or health.
The age-dependent measure of fitness, pxmx
, may decline more or less slowly over age with different diets and supplements. We tested this using a linear model with age as the independent variable and pxmx
as the dependent variable. Thus, we let zi
(t) be the value of pxmx
with diet-i [i = 1 (banana control), i = 2 (banana with R. damascena), i = 3 (apple control), and i = 4 (apple with R. damascena)] at time-t (t = 11–49 days from egg). The linear model is,
where
The effects of R. damascena on the age versus pxmx
values were also examined using Bayes factors. A Bayes factor is a ratio of two probabilities calculated under two hypotheses. These hypotheses are the probabilities of the experimental observations under the hypothesis of (i), in the denominator, a pxmx
schedule, which changes as a linear function of age, diet, and an interaction of age and diet, and (ii), in the numerator, a similar hypothesis without the interaction term. Diet here is either one of the fruit substrates (apple or banana) and fruit plus R. damascena. If this ratio is >1 that indicates support for the model with no interaction. The tests were implemented with the lmBF function in the R (
pxmx age interval statistical analysis
We tested for differences in pxmx
in 13, 3-day age classes (day from egg 10–12, 13–15, …). The observations consisted of pxmx
at an age (x) within an age interval-k (k = 1, 2, …, 13). Within each age interval, pxmx
was modeled by a straight line allowing diet-j [j = 1 (banana control), j = 2 (banana with R. damascena), j = 3 (apple control), and j = 4 (apple with R. damascena)] to affect the intercept, but not the slope of the line. Slope could vary between age intervals. Populations-i (i = 1, 2, …, 20) contributed random variation to these measures. With the notation above, the pxmx
at age (x), age interval (k), diet (j), and population (i) is yijkx
and can be described by,
where
px age-specific survivorship analysis
For each combination of treatment × sex 3-day survivorship intervals were computed. For each age interval, a new categorical variable was then created, defining the status of each one of the flies (0 = dead or 1 = alive). The counts of each age interval were used in a chi-squared test to compare all treatment combinations in both the R. damascena experiment and the combination experiment. A Bonferroni correction was applied to correct for the multiple age classes per comparison. Analysis was completed with the survival package in R (
Fecundity statistical analysis
Average eggs per surviving female (mx ) was analyzed for both experiments using a paired t test with replicates 1–5 treated as pairs across treatments analyzed. Early fecundity comprised the average of the first half of the assay (∼20 days) and later fecundity comprised the average of the second half of the assay (∼20 days). Average fecundity comprised the average of the entire assay (∼40 days). A p-value <0.05 was considered statistically significant.
Mean longevity analysis
Mean longevity was analyzed using a LME model in the R (
where
Results
R. damascena supplementation experiment
pxmx analysis
pxmx for the banana treatment declines significantly faster with age compared with the apple treatment (p < 0.0001; Supplementary Fig. S1). Notably, the rate of change in pxmx for the apple treatment is not significantly different than 0 (p = 0.1382; Supplementary Fig. S1). This is also the case for the apple with R. damascena treatment (p = 0.253; Supplementary Fig. S1). The rate of change in pxmx for the apple with R. damascena treatment and the banana with R. damascena treatment compared with their respective controls is not significantly different (p = 0.8098 and p = 0.1045, respectively; Supplementary Fig. S1). R. damascena does not significantly change the rate of change in pxmx with age (x) in either diet background (p > 0.05; Supplementary Fig. S1).
The Bayes factor, which tested the value of R. damascena on the slope of the age versus pxmx relationship, was 6.31 indicating little support for an effect of R. damascena on the slope. A similar test in the apple food yielded a Bayes factor of 10.8, again providing little support for an interaction between R. damascena and the slope.
pxmx age interval analysis
Post hoc tests were completed on age intervals to determine what ages were affected by the diet treatments. Flies exposed to either the apple treatment or the apple with R. damascena treatment generally show lower pxmx at early ages and higher pxmx at later ages when compared with the banana treatment or the banana with R. damascena treatment (Fig. 1a, d; Supplementary Tables S2 and S3). We see no differences between the apple treatment and apple with R. damascena treatment at any of the 13 age intervals (Fig. 1b; Supplementary Table S4). This is also generally the case for the banana treatment compared with the banana with R. damascena treatment, except the banana treatment is statistically higher than banana with R. damascena treatment for one age interval early in the assay (Fig. 1c; Supplementary Table S5). We see the greatest later life pxmx enhancement in the apple with R. damascena treatment when compared with the banana with R. damascena treatment (Fig. 1d).
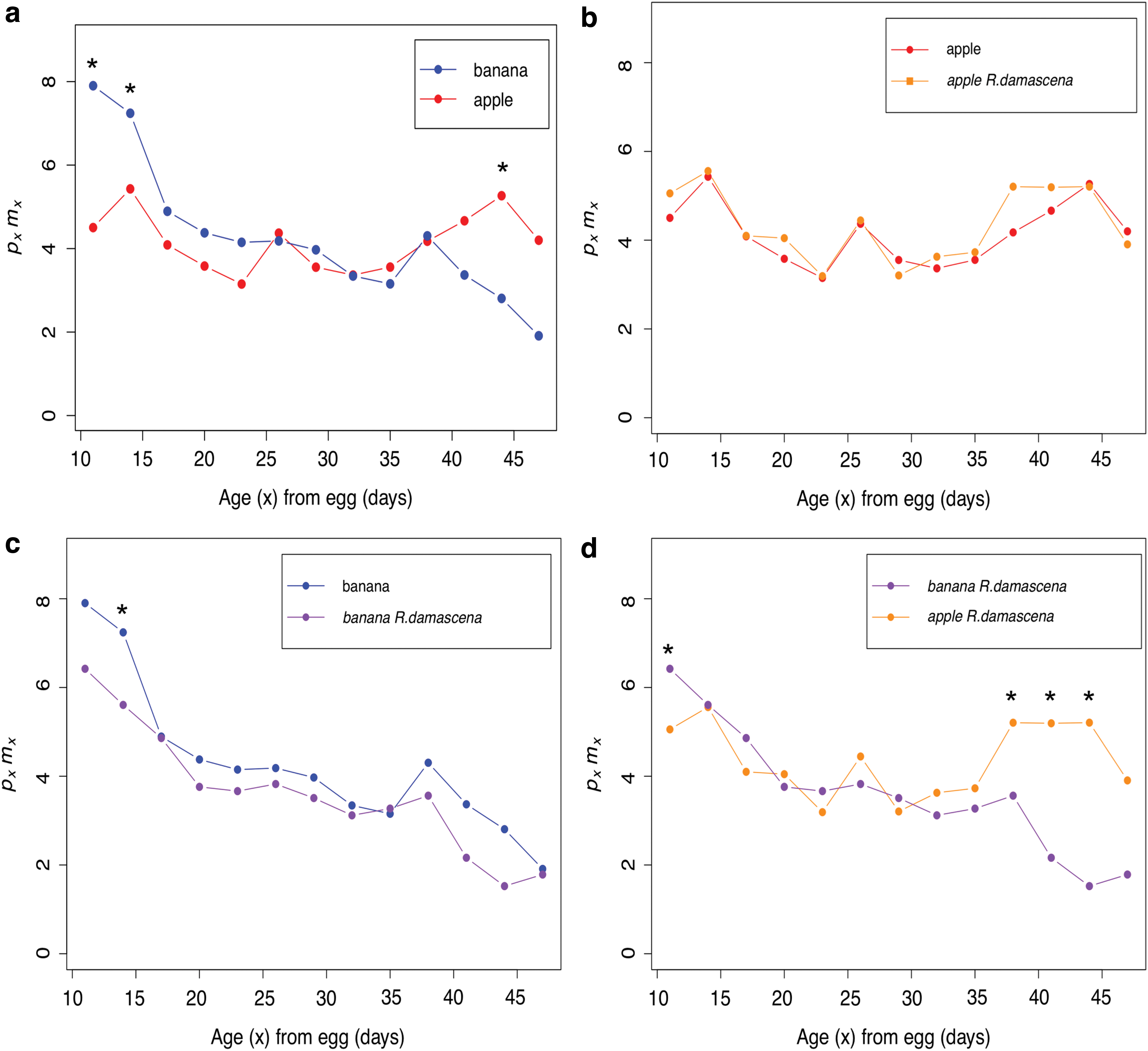
Age-dependent fitness (pxmx
) over adult age (x) for the apple and banana Rosa damascena supplementation experiment.
Age-dependent survival (px ) and mean longevity analysis
When analyzing conditional survival probability (px ), the strongest effects are observed in female flies (Fig. 2). Reponses in males are generally much weaker or nonexistent (Supplementary Fig. S2). Female survival is significantly higher on the apple diet compared with the banana diet for most age intervals (Fig. 2a). However, the first age interval is consistently higher in the treatments with the banana background compared with the apple background (Fig. 2a, d). Female mean lifespan is significantly higher (+13.5%) on the apple diet compared with the banana diet (p < 0.001; Table 1). In the banana-fed flies, female survival is significantly higher with R. damascena supplementation for a number of age intervals (Fig. 2c). R. damascena weakly increases mean longevity (+5.43%) in the banana diet background (p = 0.0469; Table 1). Interestingly, this trend is opposite with apple-fed flies. Female survival is significantly higher in the apple treatment compared with the apple with R. damascena treatment for a number of age intervals (Fig. 2b). In addition, R. damascena reduces mean longevity in the apple background (−3.0%), however, this difference is not significant (p = 0.21; Table 1). This contrast is clear when observing the survivorship curves and to a lesser extent the mortality curves for these treatments (Supplementary Figs. S3 and S4).
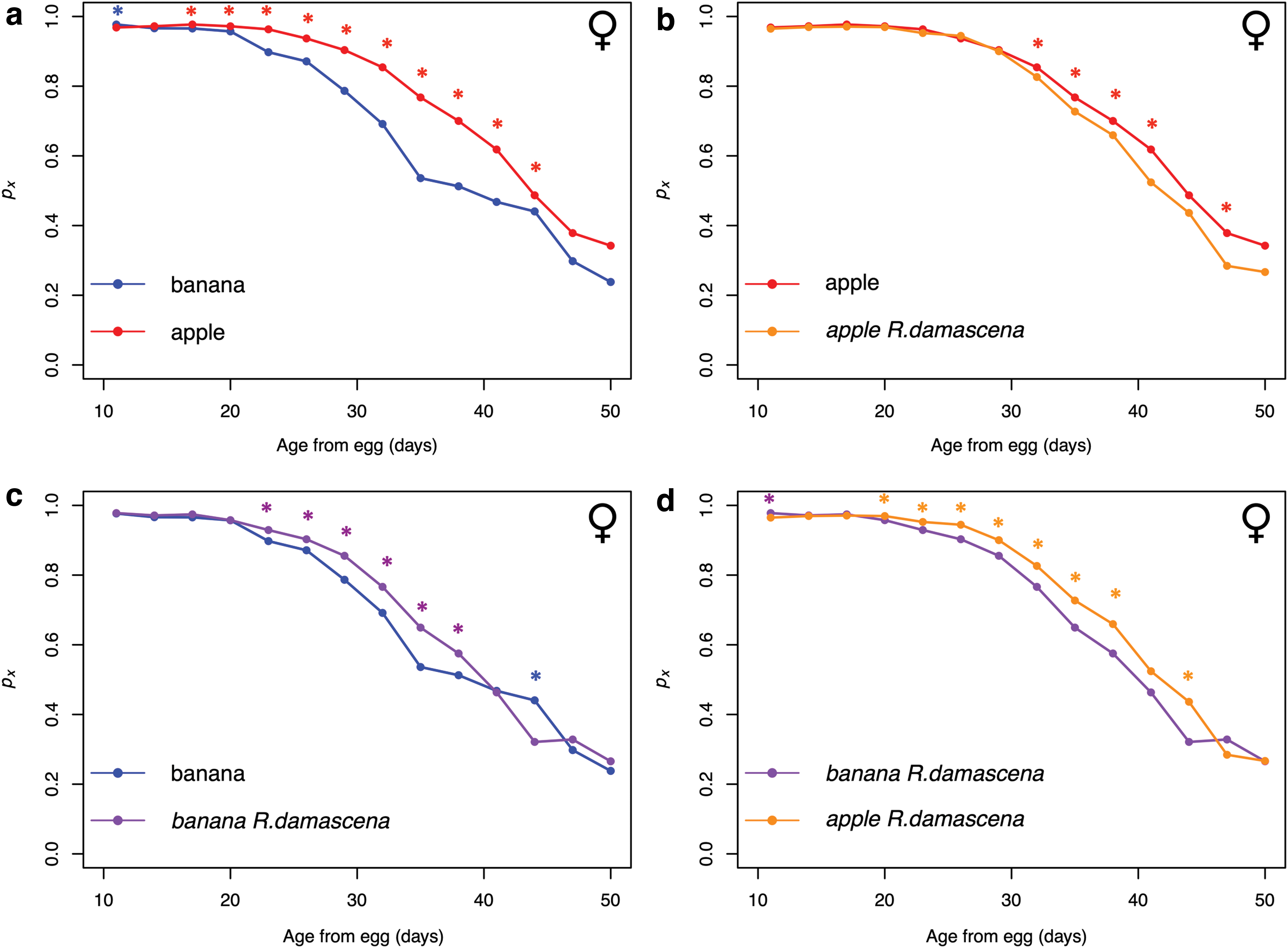
Conditional survival probability (px
) over adult age (x) for female flies in the apple and banana Rosa damascena supplementation experiment.
Results from the mean longevity analysis for the apple and banana Rosa damascena experiment
Approximately 10,000 flies per treatment were assayed in ACO1–5 (see Supplementary Table S1). p-values <0.05 are bolded.
A, apple; B, banana; Rd, Rosa damascena.
Fecundity (mx ) analysis
No difference is observed in average and late fecundity between the banana and apple treatments (p = 0.10 and p = 0.34, respectively; Fig. 3a), however, the apple diet significantly reduces early fecundity compared with the banana treatment (p = 0.01; Fig. 3a). R. damascena supplementation does not significantly affect average and early fecundity in both the banana and apple diet backgrounds (p < 0.05; Fig. 3b, c), however, R. damascena supplementation significantly decreases later fecundity in the banana diet background (p = 0.02).

Average, early, and late eggs per surviving female (mx
) across day 11–49, 11–29, and 30–49 days from egg, respectively, for the apple and banana Rosa damascena experiment.
R. rosea and R. damascena combination experiment
pxmx analysis
pxmx for the banana control declines significantly faster with age compared with the R. rosea treatment (p < 0.0001; Supplementary Fig. S5). Interestingly, the rate of change in pxmx for the R. rosea treatment is not significantly different from 0 (p = 0.158; Supplementary Fig. S5). This is also the case for the R. rosea and R. damascena combination treatment (p = 0.753; Supplementary Fig. S5). The rate of change in pxmx for the R. damascena treatment does not significantly differ from the control (p = 0.0607; Supplementary Fig. S5), and the combination treatment does not differ from the R. rosea supplemented alone (p = 0.222; Supplementary Fig. S5).
pxmx age interval analysis
Post hoc tests were completed on age intervals to determine what ages were affected by the diet treatments. When R. rosea is supplemented, we see a general trend of lower pxmx at early ages and higher pxmx at later ages when compared with the control, or R. damascena treatments (Fig. 4a, d–f; Supplementary Tables S6 and S7–S9). This is similar to the trend observed in the apple versus banana treatments, but pxmx is more reduced at earlier ages in the R. rosea treatments (Figs. 1 and 4). When observing pxmx for the control compared with the R. rosea treatment, we see significantly higher pxmx at early ages in the control and significantly lower pxmx at later ages (Fig. 4a; Supplementary Table S6). When comparing the control with the R. damascena treatment, we see no difference in pxmx at any of the age intervals (Fig. 4b; Supplementary Table S10). In addition, we see no difference in the R. rosea treatment versus the combination treatment (Fig. 4c; Supplementary Table S11).
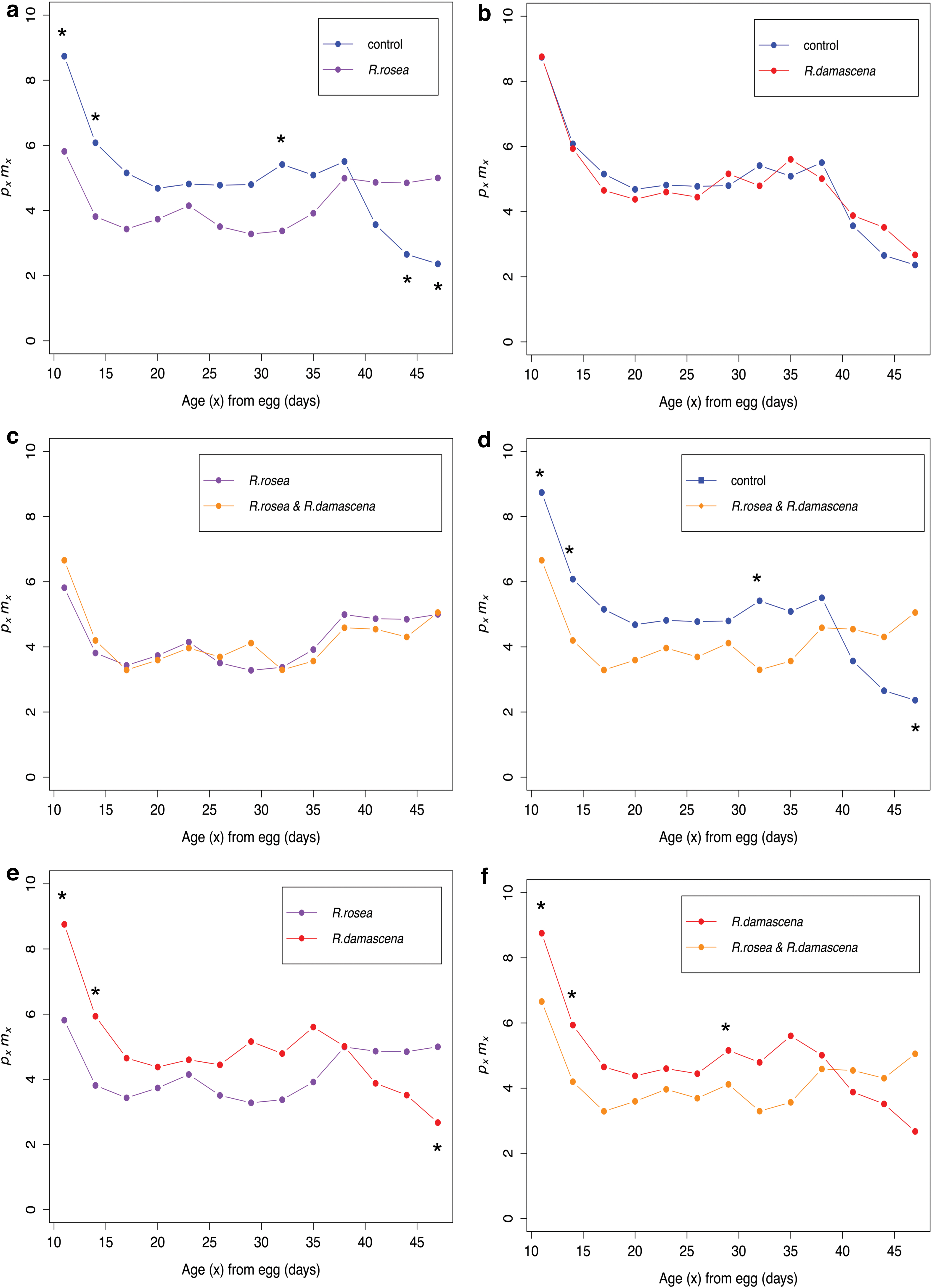
Age-dependent fitness (pxmx
) over adult age (x) for the Rosa damascena and Rhodiola rosea combination experiment.
Age-dependent survival (px ) and mean longevity analysis
When analyzing age-specific survival probability (px ), we see the greatest difference in survival in males and females from the R. rosea and combination treatments compared with the control and R. damascena treatments (Fig. 5; Supplementary Fig. S6). This contrast is clear when observing the survivorship curves and mortality curves for these treatments (Supplementary Figs. S7 and S8). R. damascena increases survival significantly in only some age intervals in the males (Supplementary Fig. S6b) and one age interval in the females (Fig. 5b), and its effectiveness in increasing survival is not as strong as the R. rosea (Fig. 5e; Supplementary S6e). Mean longevity is significantly longer in males (+16.3%) and females (+18.83%) in the R. rosea treatment compared with the control (Table 2). Mean longevity is not significantly increased in the R. damascena treatment compared with the control (Table 2). Interestingly, we see that survival is significantly decreased in a few of the later age intervals of the combination treatment compared with the R. rosea (Fig. 5c; Supplementary S6c), however, this difference is slight. Mean longevity for this comparison is reduced in the combination treatment, although it is not significant (Table 2).
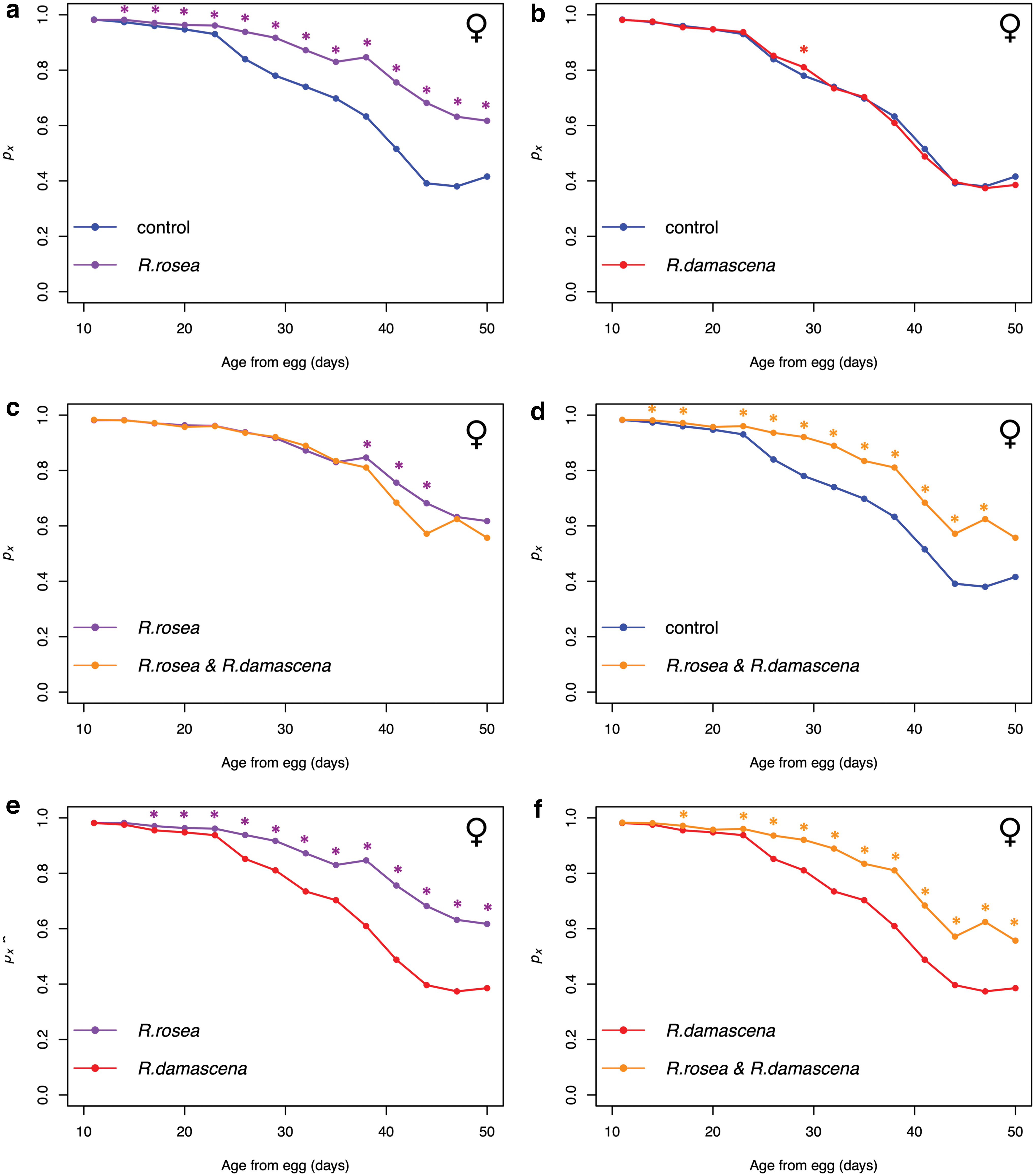
Conditional survival probability (px
) over adult age from egg (x) for female flies in the Rhodiola rosea and Rosa damascena combination experiment.
Results from the Mean Longevity Analysis for the Rhodiola rosea and Rosa damascena Combination Experiment
Approximately 10,000 flies per treatment were assayed in ACO1–5 (see Supplementary Table S1). p-values <0.05 are bolded.
c, banana control; Rd, Rosa damascena; Rr, Rhodiola rosea; RrRd, combination.
Fecundity (mx ) analysis
When analyzing fecundity, it is clear that the treatments with R. rosea supplementation have significantly lower early mx compared with the control and R. damascena treatments (Fig. 6). This dramatic decrease in earlier fecundity is to blame for the significant decrease in average mx for these treatments. However, later fecundity is not significantly different. There does not appear to be a decrease in fecundity in the R. damascena treatment compared with control (Fig. 6b). Lastly, no fecundity difference exists between the R. rosea and combination treatments (Fig. 6c).

Average, early, and late eggs per surviving female (mx
) across day 11–49, 11–29, and 30–49 days from egg, respectively, for the Rosa damascena and Rhodiola rosea combination experiment.
Discussion
In this study, we were able to reproduce the results of Rutledge 12 with flies on the apple diet having significantly higher pxmx at later ages compared with flies on the banana diet. Interestingly, flies treated with the apple diet also had significantly lower pxmx at early ages compared with flies on the banana diet, an effect that was not clear from our previous study. 12 This apparent trade-off could have evolved through antagonistic pleiotropy with genes promoting health and function on the banana diet at early ages selected for despite their negative effects later in life. 30 The apple-fed female flies lived 14% longer on average than the banana-fed females without a significant decrease in average fecundity, demonstrating that the longevity-enhancing effects are not simply artifacts of functional impairment of reproductive characters or dietary restriction (DR). 2,31 The increase in lifespan and the higher pxmx observed later in life may in part be due to the various polyphenols present in apples. Apple polyphenols mainly consist of procyanidins and have been shown to extend lifespan in D. melanogaster 32 and other model organisms. 33 –35 Supplementing Drosophila populations with specific apple polyphenols may be useful to identify the mechanism by which the apple food increases longevity, however, we would contend that the whole-diet system, rather than a single compound, is responsible for the healthspan-promoting effects through synergistic interactions. 36
If these findings are broadly applicable, they are relevant to ongoing discussions of optimal diet choices for present-day human populations. Our findings suggest that the ideal diet for younger individuals may not be the same as the ideal diet for older individuals. Humans with agricultural ancestry may achieve better health while consuming an organic agricultural diet at earlier ages, and a more paleolithic diet at later ages, compared with individuals who consume a strictly paleo diet throughout life. Future diet “switch” experiments treating flies with the apple and banana diet at different ages of life will be important to further study the age-specific benefits of these diets. We would expect flies fed the banana diet at early ages and the apple diet at later ages to have superior performance compared with flies treated with solely banana or apple.
Supplementation with R. rosea produced a similar life-extending effect as was observed in previous studies. 17,37,38 In the present study, R. rosea significantly extended lifespan by 16% in males and 19% in females. Age-specific survivorship was higher for almost every age class in both males and females. Although DR may be one of the most robust methods for extending lifespan, 39 previous studies using R. rosea suggest that the increase in lifespan works independently of DR and major aging pathways such as sirtuins, the target of rapamycin (mTOR), and insulin and insulin-like signaling. 37 However, it may depend on carbohydrate or caloric content of the background diet. 40 Contrary to findings in Jafari et al., 17 R. rosea significantly reduced early fecundity (−29%) and average fecundity (−18%). Late fecundity was not affected. Jafari et al. 17 found that R. rosea decreased fecundity, but only at doses four times higher than the dose used in this study. One possibility is that this significant early fecundity effect arose with this particular “batch” of R. rosea. Another confounding factor is that Jafari et al. 17 conducted experiments in vials, whereas the present study's fecundity assay was performed in population cages.
Despite this significant decrease in female fecundity, the trend in pxmx is quite similar to what is observed in our apple versus banana treatment; pxmx is significantly higher in the control at early ages and lower at later ages, compared with the R. rosea treatment. This switch to better performance in the R. rosea treatment occurs around 38 days from egg, as is observed in the apple diet. In addition, we found significantly higher survivorship and greater mean longevity in the apple diet and R. rosea treatment compared with the banana control treatment. Our results suggest that supplementing R. rosea later in life may be more ideal rather than supplementing at all ages. More experiments should be completed testing the effects of R. rosea supplementation at various ages of life.
We were also able to partially reproduce the R. damascena results from Jafari et al. 18 Using the same background diet as was used in that study (banana–molasses medium), we found that females had significantly higher survivorship for days 22–40 from egg when supplemented with R. damascena. Female mean longevity increased significantly (∼5%), however, this was far from the 22% increase in the previous study. 18 We observed an increase in longevity and survivorship without a significant decrease in average female fecundity, again suggesting that the longevity-enhancing effects of R. damascena are not simply artifacts of the impairment of reproduction. However, pxmx was not significantly different between the banana control flies and the R. damascena-supplemented flies for any of the age classes. The mechanisms by which R. damascena increases lifespan are not fully understood; however, a previous study found that it has protective effects against oxidative stress in females only, and no effect on mitochondrial function or antioxidant defenses. 27
We see some evidence of a Poisoned Chalice Effect with R. damascena supplementation decreasing female age-specific survivorship for five age classes (covering 15 days) when supplemented with the apple treatment. Mean longevity was also reduced, although not significantly. It is well known that food and nutrition play an inextricable role in all aspects of drug absorption and metabolism. 41 This finding suggests that the lifespan extension with R. damascena not only depends on diet; but combining substances with survivorship-enhancing properties does not necessarily produce a positive additive effect. In fact, the interaction may be detrimental to the organism, as is observed with our study for age-specific survivorship.
Lastly, we observe limited evidence of a Poisoned Chalice Effect occurring in the combination of R. rosea and R. damascena. This occurs particularly in age-specific survivorship with female flies receiving R. rosea supplementation only outperforming flies on the combination for three later age intervals (Fig. 5c). This effect was not observed with p x m x or fecundity. This will remain an active area of research in our laboratory, with more supplement combinations to be studied.
Conclusion
In our study, we were able to successfully reproduce the lifespan-extending effects of R. rosea and R. damascena on a large scale with ∼10,000 outbred flies used per treatment rather than hundreds. Our results indicate that supplementing a healthspan-extending diet (apple food) with a healthspan-extending botanical supplement (R. damascena) does not result in an additive beneficial effect. In fact, a negative interaction may be occurring, resulting in a decrease in age-specific survivorship. The effect of R. rosea on healthspan mimics the effect of the ancestral apple fly diet with pxmx significantly lower at early ages and higher at later ages. Lastly, combining botanical supplements may result in negative effects on healthspan. More experiments on the effects of combining different healthspan-extending substances in various diet backgrounds is an obvious next step for this line of work.
Footnotes
Acknowledgments
The authors thank the School of Biological Sciences, the Undergraduate Research Opportunities Program (UROP), and the Summer Undergraduate Research Opportunities Program (SURP) at UC Irvine for funding this work. The authors thank the Department of Education's GAANN program for funding this research. A special thanks to the undergraduate researchers of the Rose, Mueller, and Jafari laboratories for all their hard work on this project. The total number of flies and eggs processed was 105,459 and 6,245,725, respectively.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The authors received support from The University of California Regents, UC Irvine Undergraduate Research Opportunities Program (UROP), UC Irvine Summer Undergraduate Research Opportunities Program (SURP), and U.S. Department of Education Graduate Assistance in Areas of Nation Need (GAANN).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Supplementary Figure S6
Supplementary Figure S7
Supplementary Figure S8
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
Supplementary Table S5
Supplementary Table S6
Supplementary Table S7
Supplementary Table S8
Supplementary Table S9
Supplementary Table S10
Supplementary Table S11
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.