
Abstract
The telomere length (TL) has increasingly been used as a biomarker of human aging because it has been shown to predict the chances of survival and longevity. Oxidative stress is presumed to be a major cause of telomere shortening, but the importance of oxidative stress as a determinant of telomere shortening remains less clear and has recently been questioned. We analyzed 105 healthy subjects of both sexes between the ages of 20–77 years. The TL and biomarkers of oxidative stress were estimated as per standard protocols. A significant (p < 0.001) age-dependent decline in TL was observed. TL was positively correlated with the ferric reducing ability of plasma value (r = 0.8811) and reduced glutathione (r = 0.8209), whereas negatively correlated with malondialdehyde (r = −0.7191). Our findings supported the idea of a possible correlation between the TL and biomarkers of oxidative stress in aging. The study has remarkable scope in medical science as the findings on correlation of TL with biomarkers of oxidative stress in aging are novel and they will help in further research against oxidative stress.
Introduction
Telomeres are chromosome end components in eukaryotes that stabilize the ends of chromosomes. 1 The telomeres usually are consecutive repeats of simple short sequences in which one strand is rich in guanosine residues. The normal DNA polymerase cannot replicate the telomeres. This results in the shortening of telomeres with each cellular division. 2 The overhanging single-strands left after replication, look like damaged DNA and hence require protection from DNA repair systems.
In most organisms including humans, telomeres form protective loops by binding to complementary sequences in the overhang itself. The shortening of telomeres has been shown related to the aging of the organism. It has been shown that telomere shortens with age and in several age-associated diseases. 3,4 Consequently, telomere length (TL) has been suggested as a biomarker of “biological age.” 5
Oxidative stress generated by environmental and genetic reasons can alter the TL in the blood. 6 Telomeres are more prone to oxidative stress than the whole chromosome. 7 Reactive oxygen species (ROS) and free radicals are generated during normal metabolic reactions and chronic inflammation. The ROS and free radicals oxidize the cellular components such as lipids, proteins, nucleic acids, and generate alterations in their functions. 8 The chronic accumulation of these ROS and damaged by-products cause oxidative stress. According to the free radical theory of aging, the accumulation of oxidative stress over the lifespan of an organism leads to aging of the organism. 5
Now the question arises—“Is there any correlation between oxidative stress and the telomere shortening during aging?” There have been several attempts to answer this question. The correlation between telomere shortening and oxidative stress has been studied in mouse models, human populations, and in cell cultures. 9 However, there is no unequivocal evidence on the fact that the telomere shortening directly relates to the oxidative stress. Some studies support the correlation between oxidative stress biomarkers and TL during aging and age-related diseases, 10 –12 whereas others consider TL as a biomarker of somatic redundancy. 13,14
We measured TL variation in 105 healthy subjects as a function of age. To investigate the extent to which oxidative stress and TL were correlated, the panel of oxidative stress biomarkers, including total antioxidant potential in terms of ferric reducing ability of plasma (FRAP) value, glutathione (GSH), and lipid peroxidation in terms of malondialdehyde (MDA), was assessed.
Materials and Methods
Sample
The sample consisted of 105 normal healthy subjects of both sexes between the ages of 20 and 77 years. Exclusion criteria include (1) treatment with drugs, (2) smokers, and (3) acute or a chronic general medical condition such as diabetes mellitus, asthma, tuberculosis, or any other major illness. In addition, women of child-bearing age were excluded if they were pregnant or in the postpartum period. The study protocol was approved by the Institutional Human Ethics Committee of Central University of Haryana (Reference no. CUH/2020/IHEC/03).
Blood collection
A sample of 10 mL of peripheral blood was obtained from each subject by arm venipuncture. The blood collection was made in a standardized time of day (between 08:00 A.M. and 10:00 A.M.) and after a 12 hours period of fasting. The blood was centrifuged at 1800 g for 10 minutes at 4°C. After removal of plasma, buffy coat, and upper 15% of the packed red blood cells, the RBCs were washed twice with cold PBS (0.9% NaCl, 10 mM Na2HPO4, pH 7.4).
Measurement of leukocyte TL
In the morning, 10 mL of the blood of all healthy volunteers was collected in EDTA tubes for DNA extraction. The DNA was extracted using the salting-out method, according to Miller et al. protocol. 15 After isolation, the DNA samples were quantified and diluted to 50 ng/μL. The TL measurement was performed by multiplex real-time polymerase chain reaction (PCR) as described by Cawthon with some modifications. 16 The relative ratio (T/S ratio) of ng of telomeres (T) to ng of albumin (single-copy gene, S) was determined in experimental samples using a standard curve. The T/S ratio is proportional to the average TL.
A total of 105 blood samples were analyzed. The final volume in the reaction wells was 25 μL and contained 12.5 μL of 2 × SYBR® Select Master Mix (4472920; Life-Thermo Fisher Scientific), 1 μL sample DNA, 0.9 μL of 25 μM telg (final concentration 900 nM), 0.6 μL of 25 μM telc (600 nM), 0.9 μL of 25 μM albu (900 nM), 0.9 μL of 25 μM albd (900 nM), and 8.2 μL of UltraPure™ DNase/RNase-Free Distilled Water. The standard curve was built based on a 5-point 1:3 serial dilution DNA from a single individual (range from 150 to 1.85 ng). The samples and standard curve were all run in triplicates in a Real-Time PCR System with a fast 96-Well Block (Life-Thermo Fisher Scientific).
The conditions of PCR amplifications were 15 minutes at 95°C, 2 cycles of 15 seconds at 94°C, 15 seconds at 49°C, 35 cycles of 15 seconds at 94°C, 10 seconds at 68°C, and 15 seconds at 74°C with the acquisition of fluorescence (detection of telomere amplification), 10 seconds at 85°C, 15 seconds at 88°C with the acquisition of fluorescence (detection of albumin amplification) and the Melting Curve.
In all runs, a seriated dilution from a standard DNA was added to construct a standard curve used to relatively quantify TL with the albumin gene (T/S ratio). 17
FRAP assay
The FRAP values were determined by following the method of Benzie and Strain. 18 Working FRAP reagent was prepared by mixing acetate buffer (300 mM, pH 3.6), 2,4,6-tri[2-pyridyl]-s-triazine (10 mM in 40 mM HCl) solution and FeCl3.6H2O (20 mmol/L) solution in 10:1:1 ratio, respectively. Three milliliters of FRAP reagent was mixed with 100 μL of plasma; the content was mixed vigorously for making a uniform solution. The absorbance was read at 593 nm at the interval of 30 seconds for 4 minutes. An aqueous solution of known Fe2+ concentration in the range of 100–1000 μmol/L was used for calibration. Using the regression equation, the FRAP values [μmol Fe (II) per liter] of the plasma were calculated.
Determination of MDA content
Erythrocyte MDA was measured according to the method of Esterbauer and Cheeseman. 19 Packed erythrocytes (0.2 mL) were suspended in 3 mL Krebs'-Ringer phosphate buffer (KRP), pH 7.4. The lysate (1 mL) was added to 1 mL of 10% trichloroacetic acid (TCA) and the mixture was centrifuged for 5 minutes at 1000 g. The supernatant (1 mL) was added to 1 mL of 0.67% thiobarbituric acid (TBA) in 0.05 M NaOH and boiled for 20 minutes at a temperature >90°C, cooled and the absorbance was read at 532 nm (OD1) and 600 nm (OD2). The net optical density (OD) was calculated after subtracting absorbance at OD2 from that at OD1. The concentration of MDA in erythrocytes was determined from a standard plot. The concentration of MDA is expressed as nmol/mL of packed erythrocytes.
Determination of erythrocyte GSH
Erythrocyte GSH was measured following the standard method. 20 The method was based on the ability of the sulfhydryl group of GSH peptide to reduce 5,5′-dithiobis-2-nitrobenzoic acid (DTNB) and form a yellow anionic product whose OD was measured at 412 nm. The concentration of GSH was expressed in milligrams per milliliter of packed red blood cells and was determined from a standard plot.
Statistical analysis
Relationships between various parameters were assessed using the Pearson correlation coefficient (r). Statistical analyses were performed using GraphPad Prism version 8.00 for Windows, GraphPad Software (San Diego, CA). A p-value <0.05 was considered statistically significant.
Results and Discussion
In our study, we hypothesized that the TL in leucocytes was correlated with oxidative stress biomarkers in erythrocytes and the total antioxidant capacity of blood as a function of age. The oxidative stress biomarkers such as levels of MDA content, levels of GSH in RBCs, and total antioxidant status (TAS) in plasma were assayed. The TL was quantified relatively with the albumin gene, a housekeeping gene (T/S ratio) using the standard curve. We found that the T/S ratio was significantly (p < 0.000) decreasing (r = −0.4047) with age as shown in Figure 1.
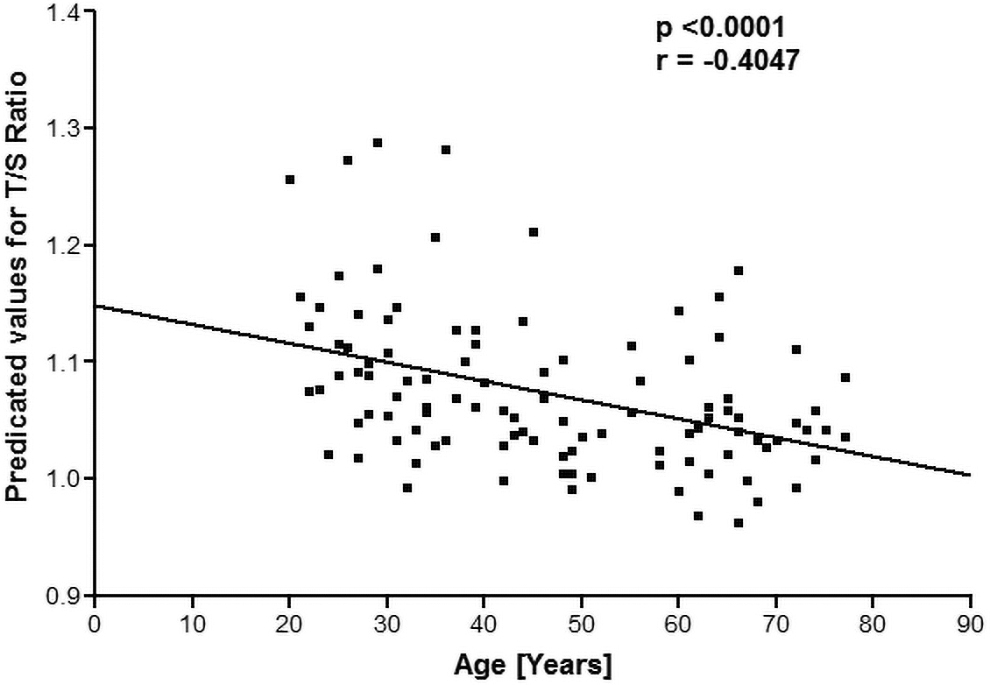
The predicted value of the T/S ratio plotted as a function of human age. T/S, relative ratio of ng of telomeres and ng of albumin genes.
The TAS was assayed in the form of FRAP. The change in absorbance per min (ΔAmin −1) was calculated for each sample and FRAP values [μmol Fe (II) per liter] were calculated from the standard plot plus plotted against the age of subjects. We observed that there was a positive (r = 0.8811) significant (p < 0.001) correlation between FRAP values and TL with increasing age. Figure 2 shows the correlation between TL and total antioxidant potential, measured in terms of FRAP values, by plotting a quotient of T/S ratio/FRAP values as a function of human age.
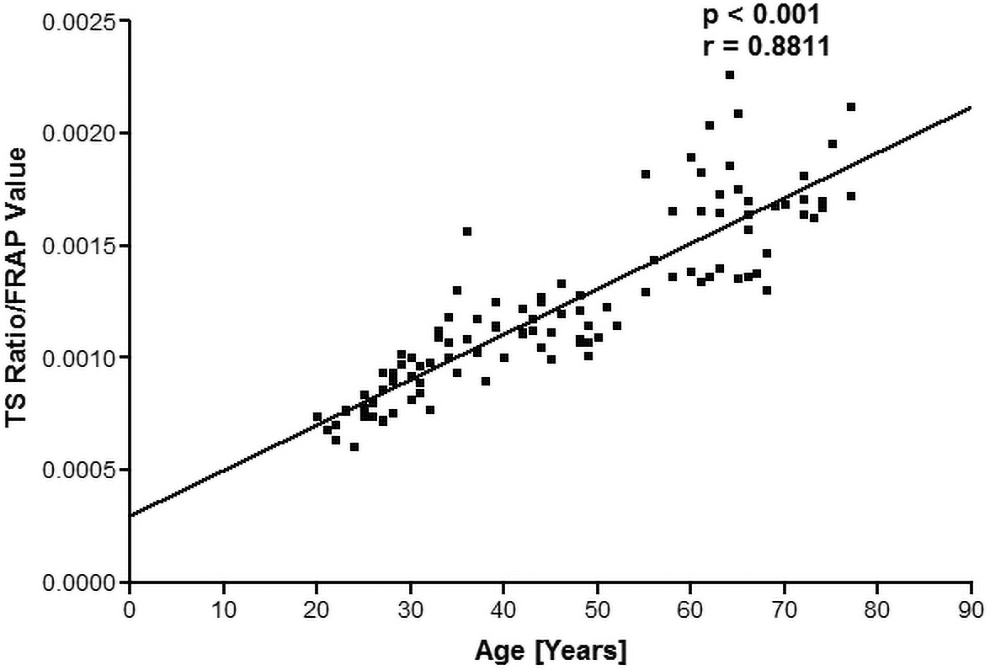
Plot of the quotient (T/S ratio/FRAP values) as a function of human age. FRAP (Ferric Reducing Ability of the Plasma) values expressed as μmol Fe (II) per liter of plasma. FRAP, ferric reducing ability of plasma; T/S, relative ratio of ng of telomeres and ng of albumin genes.
The levels of oxidative damage were calculated through MDA assay as nmol/mL of packed erythrocytes. The correlation analysis showed that there was a significant (p < 0.001) negative correlation (r = −0.7191) between TL and MDA levels as shown in Figure 3. The levels of GSH, an antioxidant were measured as milligrams per milliliter of packed red blood cells against a standard curve. We monitored that the levels of GSH were decreasing with the age of the subject. Figure 4 shows the correlation between TL and GSH, by plotting a quotient of T/S ratio/GSH values as a function of human age. TL positively correlated (r = 0.8209) with GSH as a function of human age.
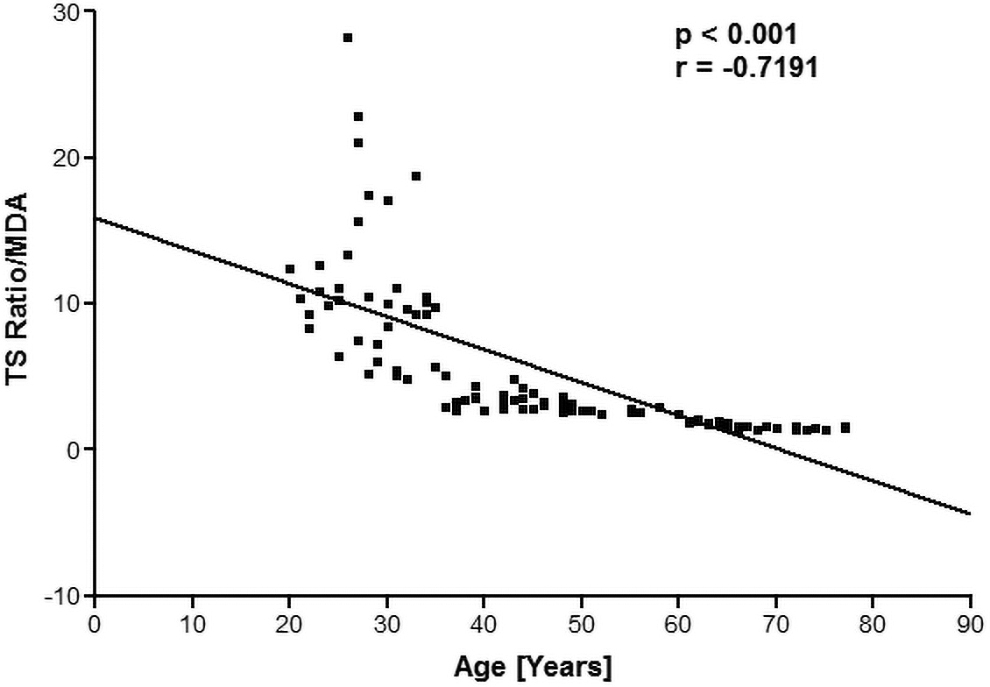
Plot of the quotient (T/S ratio/MDA) as a function of human age. The concentration of MDA is expressed as nmol/mL of packed erythrocytes. MDA, malondialdehyde.
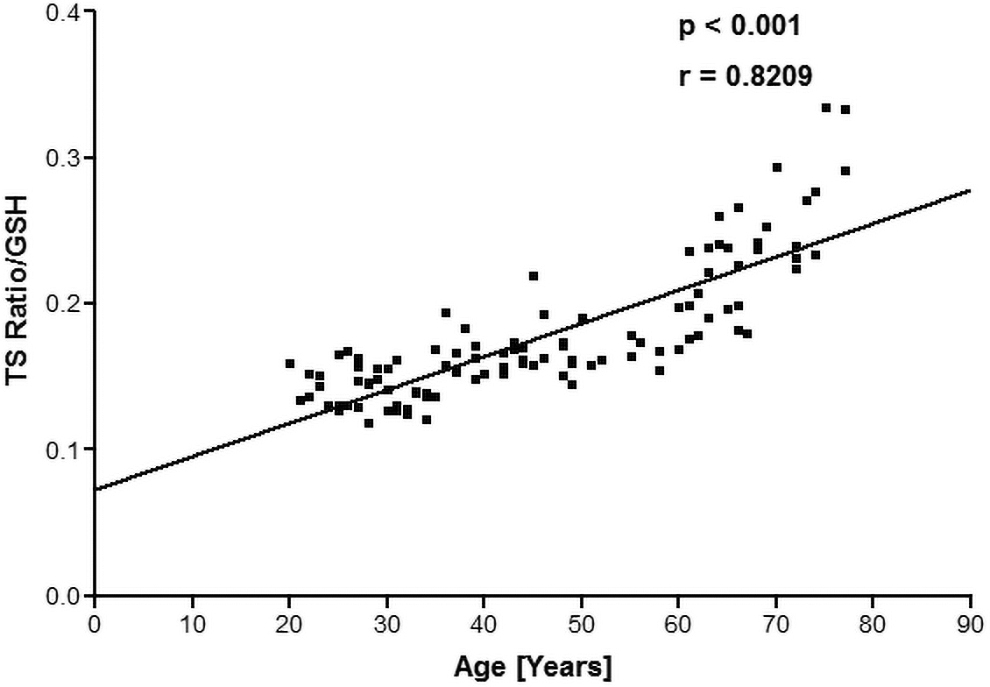
Plot of the quotient (T/S ratio/GSH) as a function of human age. The concentration of GSH is expressed in milligrams per milliliter packed erythrocytes. GSH, glutathione.
In Figure 1, the correlation analysis shows that TL is significantly (p < 0.000) negatively correlated (r = −0.4047) with age. Age-dependent decrease in TL has been reported previously. 21 Cawthon et al. showed that the relative T/S ratio decreased by 0.0048 for every 1-year rise in age, equating to around 14 bp of telomere sequence lost per year. 22 This study also substantiates the increase in mortality rate with aging due to telomere shortening. A significant (p < 0.001) positive correlation (r = 0.8811) was observed between TL and total antioxidant capacity.
We have already reported a significant decrease in FRAP value as a function of human age. 23 Lipid oxidation occurs at a quicker pace as a result of oxidative stress, producing MDA as a by-product. Plotting a quotient of T/S ratio/MDA during human aging reveals a significant (p < 0.001) negative correlation (r = −0.7191) between TL and MDA. Human aging has been linked to a decrease in GSH levels. 20 It was found that intracellular GSH levels in RBCs were negatively (r = −0.5261) correlated with age of human subjects.
This study provides novel evidence that oxidative stress biomarkers in RBCs and plasma are significantly correlated with the shortening of TL in leucocytes during human aging. The results establish that the FRAP value and GSH in the Indian population drop with age. This demonstrates that the elderly population suffer from more oxidative stress than the young; this result could be linked to lower levels of dietary antioxidants. 24 As a result, our data affirm the substantial role of oxidative stress in TL attrition during aging of human subjects.
Increased oxidative stress and decreased antioxidant levels cause telomere shortening, which leads to accelerated aging and other age-related disorders such as cardiovascular problems and neuropsychiatric disorders. Oxidative stress and an increase in proinflammatory mediators such as tumor necrosis factor-α, interferon-γ, interleukin-6, and 10 in response to antioxidant depletion are two proposed mechanisms that contribute to TL shortening. 25,26
Although our correlative data support a link between oxidative stress and telomere attrition, further in vivo studies are needed to fully comprehend the role of oxidative stress in TL variation during human aging. The accumulation of ROS and RNS can cause oxidative stress; however, a majority of intracellular ROS are generated as a by-product of aerobic respiration. 23 When the antioxidant defense system and repair mechanisms deteriorate with age, oxidative stress rises. Telomeres are thought to be particularly susceptible to oxidative damage due to the high content of guanine. 27 Unchecked oxidative stress leads to an accumulation of DNA damage, which ultimately results in accelerated telomere shortening. 28
Telomere shortening has been reported in many mental disorders. 3,29,30 Various mechanisms have been proposed to explain the process. The oxidative damage also affects the replication of telomeres. The 8-oxoguanisine, a by-product of oxidative DNA damage, replaces guanosine, which results in the arrest of replication. 31 However, it remains unclear that whether oxidative stress has a direct or indirect influence on TL. The TL in leukocytes was shown to be associated with oxidative stress in the blood of yoga practitioners. 32 The TL, TAS, and levels of plasma homocysteine were more in yoga practitioners who were practicing yoga for 2 years. Whereas the levels of MDA were lesser in the yoga practitioners than in the control. The TLs were positively correlated with TAS and negatively correlated with MDA and plasma homocysteine.
This study has some limitations. Our results could be influenced by distinct leukocyte-subtype proportions found in different blood samples. The T/S ratio was calculated using average TL, which cannot be interpreted as a direct measure of the telomeric region. Furthermore, oxidative stress levels could have fluctuated over time during blood collection. We also did not measure telomerase activity, which could have impacted current findings because telomerase affects the rate of TL change in peripheral blood mononuclear cells. 33
Conclusion
Taken together, our findings supported the idea of a possible correlation between the TL length and biomarkers of oxidative stress during aging. Therefore, prospective and experimental studies are essential to establish a causal relationship between TL and oxidative stress markers and also the possible mechanisms involved. Based on our findings, we also propose the utility of antioxidative therapies in the treatment of aging-related diseases. In view of tremendous scope for these novel findings in medical and diagnostics and therapeutics, we believe that these findings will aid oxidative stress studies in human aging.
Footnotes
Authors' Contributions
SY: Conceptualization, Writing-Original draft preparation. PKM: Supervision, Reviewing, and Editing.
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships.
Funding Information
The study was supported by a fellowship from the Council of Scientific and Industrial Research (CSIR), Government of India to S.Y. (09/1152(0013)/2019-EMR-I).