
Abstract
Diabetic encephalopathy (DE) is one of complications of diabetes mellitus. Carnosine is a dipeptide composed of β-alanine and
Introduction
In the past three decades, the number of people with diabetes mellitus has quadrupled, and diabetes mellitus has become the ninth leading cause of death in the world. 1 The epidemic of diabetes mellitus and its complications, such as diabetic encephalopathy (DE), diabetic nephropathy, and diabetic eye disease, pose a threat to global health. 2 The pathogenesis of DE is not clear, but it seems to be related to insulin resistance, hyperinsulinemia, hyperlipidemia and insulin deficiency. 3 The pathology of DE mainly includes neurodegeneration, 4 oxidative stress, 5 inflammation, 6 changes of neurotransmitters, 7 neural apoptosis, 8 and finally cause cognitive impairment in patients.
Previous studies showed that the Ser199/Ser202 sites of microtubule-associated tau protein were hyperphosphorylated and the expression of β-amyloid (biomarkers of Alzheimer's disease, AD) was increased in the hippocampus of diabetic mice, indicating that DE were related with AD pathology. 9,10
Endoplasmic reticulum (ER) is the cell's Ca2+ storage, protein folding, and assembly factory. 11 Various stress (such as hypoxia, starvation, etc.) leads to the interruption of ER protein folding. Unfolded or misfolded proteins will accumulate in the ER, which is referred to as unfolded protein response (UPR). 12 The unfolded protein accumulates in ER and exceeds the load of ER, causes ER stress and ultimately leads to cell apoptosis. 13 Some studies have shown that ER stress exists in the brains of diabetic mice. 14,15 In addition, our previous studies have proved that berberine and quercetin can reduce the development of DE by alleviating ER stress in the brain of db/db mice. 16,17
The sirtuins family, a highly conserved family of NAD+ dependent deacetylases, plays an important role in regulating life span and aging, cancer, obesity, insulin resistance, inflammation, and energy metabolism. 18 Recent studies have shown that reduced sirtuin6 (SIRT6) activity is associated with obesity and diabetes. 19 SIRT6 can regulate pancreatic β-cell function to maintain normal blood glucose level. 20 Although the role of SIRT6 in DE has not yet been reported, some studies have shown that SIRT6 can alleviate cognitive impairment in neurodegenerative diseases. 21,22 Besides, some studies have shown that the activation of SIRT6 could alleviate cell damage caused by ER stress. 23,24
Carnosine is a dipeptide composed of β-alanine and
As a dietary supplement, carnosine can significantly inhibit hyperglycemia and pancreatic β-cell loss in obese type 2 diabetic mice (db/db). 30 In neurodegenerative diseases, carnosine can improve cognitive impairment by exerting antioxidation, reducing mitochondrial dysfunction, anti-inflammatory, relieving ER stress, and reducing the expression of β-amyloid. 31 –34 Besides, our previous study has shown that carnosine could increase the expression of SIRT6 to improve cognitive impairment in SAMP8 mice. However, whether SIRT6/ER stress pathway participated in carnosine on DE treatment is still unknown.
In this study, we used an animal model of type 2 diabetes (db/db mice) to explore the neuroprotective effect of carnosine on DE and its potential mechanism. We found that carnosine could reduce neuronal damage and ameliorate cognitive impairment in db/db mice by regulating SIRT6/ER stress pathway.
Materials and Methods
Materials
Primary antibodies of postsynaptic density 95 (PSD95, CAS:36233s), sirtuin-6 (SIRT6, CAS:#12486), binding immunoglobulin protein (BIP, CAS:#3177), protein kinase R-like endoplasmic reticulum kinase (PERK, CAS:3192s), phospho-protein kinase R-like endoplasmic reticulum kinase (P-PERK, CAS:3179s), inositol-requiring enzyme-1α (IRE1α, CAS:3294T), activating transcription factor 6 (ATF6, CAS:65880s), and C/EBP-homologous protein (CHOP, CAS:#2895) were purchased from Cell Signaling Technology, Inc. Primary antibodies of β-actin (ACTB, CAS:ab8227) and brain-derived neurotrophic factor (BDNF, CAS:ab108319) were purchased from Abcam, Inc. Primary antibody phospho-inositol-requiring enzyme-1α (P-IRE1α, CAS:NB100-2323) was purchased from Novus Biologicals. Secondary antibodies (goat anti-mouse IgG, CAS: S0002 and goat anti-rabbit IgG, CAS: S0001) were from Affinity Biosciences.
Animals and drug treatment
The db/db mice and age-matched wild-type C57BL/6J-db/m mice were supplied by Nanjing Biomedical Research Institute of Nanjing University, Nanjing, China (10-week-old, female). All mice were kept in a specific pathogen-free animal room under controlled temperature (24°C ± 1°C) and humidity (55% −70%) and get free access to water and food. After 6 weeks of adaptive housing and feeding, the mice were randomly divided into three groups (10 mice per group): control group (db/m, 0.9% saline), diabetic group (db/db, 0.9% saline), and carnosine treatment group (db/db + carnosine). The mice were intragastrically administered with 0.9% saline or carnosine (100 mg/kg) once daily at the age of 16 weeks and lasted for 8 weeks. The dose of carnosine was chosen according to our previous study. 31
Ethics statement
All experimental procedures were carried out according to the guidelines of the Administrative Panel on Laboratory Animal Care of Guangzhou University of Chinese Medicine and approved by the Committee for Animal Research of Guangzhou University of Chinese Medicine (Guangzhou, China). IRB approval number: 20160671.
Morris water maze test
After 8 weeks drug treatment, learning and short-term memory functions in mice were detected by Morris water maze through a water maze equipment (Guangzhou Feidi Biology Technology, Guangzhou, China). The water maze equipment consisted of a black circular pool, a black platform, and recording system. The circular pool was filled with nontoxic white coloring water and was divided into four equal quadrants.
The black escape platform was placed 2 cm below the water surface in the center of the fourth quadrant. The mice participated in the navigation test for five consecutive days. Calculated the time it takes mice to find the platform every day. For the sixth day, the mice performed a space exploration experiment, calculated the time they spent on the original platform, the number of times they enter the original platform and their swimming speed.
Brain tissue collection
After the behavior test, the mice were anesthetized intraperitoneal with sodium pentobarbital (30 mg/kg). Then they were sacrificed by cervical dislocation. The brain tissues of the mice were rapidly removed. And the cerebral cortex and hippocampus were separated from the brains. Then the brain tissues were quickly removed to the ice and immediately stored at −80°C.
Oxidative stress and NOS level test
The brain tissues of the mice were made into a 10% tissue homogenate with ice-cold saline and centrifuged at 12,000 g 10 minutes at 4°C. Then the supernatant was collected to determine the total protein concentration by using a bicinchoninic acid (BCA) and detect the levels of MDA, SOD, CAT, GSH-PX, TNOS, iNOS, and cNOS according to the corresponding reagent instructions. The absorbance was measured at a wavelength of 532, 550, 405, 412, and 530 nm, respectively.
Western blot analysis
Brain tissues were made into a 10% tissue homogenate with ice-cold RIPA lysis solution and centrifuged at 14,000 g for 10 minutes at 4°C, the supernatant was collected to determine the total protein concentration by using a BCA. After that added appropriate volume of loading buffer and boiled for 10 minutes at 100°C. Separated the proteins with different concentrations of sodium dodecyl sulfate (SDS) polyacrylamide gels and transfered the proteins to the polyvinylidene fluoride (PVDF) membranes. The membranes were blocked by 5% skimmed milk. Then the membranes were incubated with the primary antibody (PSD95, BDNF, SIRT6, BIP, PERK, P-PERK, IRE1α, P-IRE1α, ATF6, CHOP, and ACTB) at 4°C overnight, and incubated with (goat anti-rabbit IgG or goat anti-mouse IgG) for 1 hour. Blots digital images were visualized with an Image Lab 3.0 (Bio-Rad).
The quantification of the results were analyzed with Image J software.
Statistical analysis
The experimental data were expressed as mean ± standard error of the mean. Data analysis was performed using SPSS 24.0 (SPSS, Inc.), and then data processing was performed using GraphPad Prism 8 software (GraphPad, Inc.). Statistical differences in data between groups were performed with analysis of variance (ANOVA) (one-way or two-way ANOVA), and followed by a post hoc test (Dunnett). p < 0.05 was presented as statistically significant.
Results
Carnosine ameliorates cognitive impairment in db/db mice
The cognitive impairment of db/db mice and the therapeutic effect of carnosine were evaluated by Morris water maze test. In Figure 1A, the swimming path of db/db mice were disordered, compared with db/m and carnosine treatment group. In addition, they could hardly find the target platform within 60 seconds. As shown in Figure 1B, in the Morris water maze test, the time for mice to find the target platform were decreased gradually in the five consecutive days in all groups. However, db/db mice needed more time to find the target platform compared with db/m, which demonstrated that cognitive impairment appeared in db/db mice.
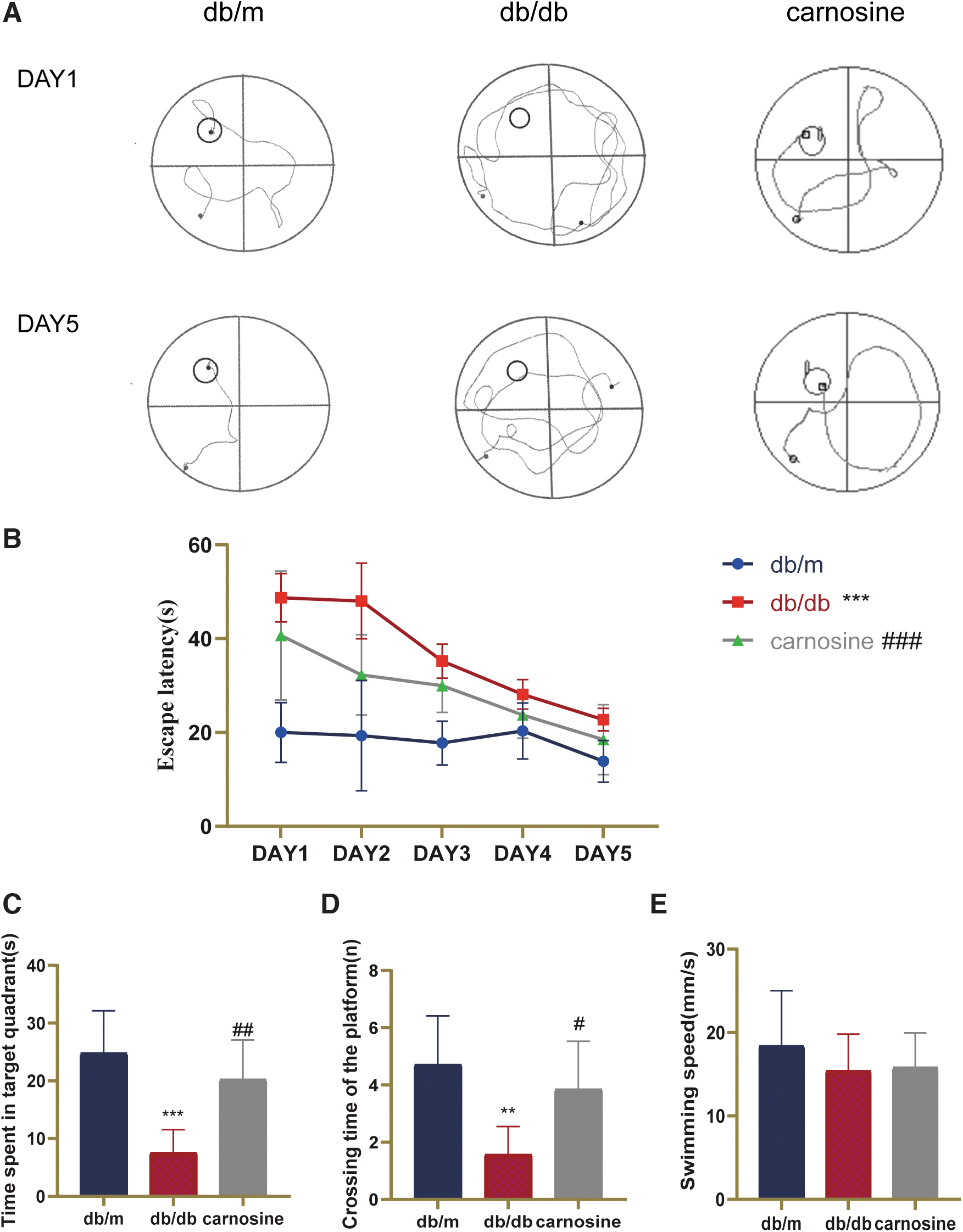
Carnosine ameliorates cognitive impairment in db/db mice.
After treatment with carnosine, db/db mice shortened the time to find the target platform. In Figure 1C and D, db/db mice spent less time in original platform position and made fewer attempts to arrive the original platform position than db/m mice. Carnosine treatment significantly increased the time of db/db mice spent on original platform and increased the number of times for crossing the original platform position. There is no significant difference in swimming speed among these groups. Therefore, these data demonstrated that carnosine could ameliorate cognitive impairment in db/db mice.
Carnosine reduces neuronal oxidative stress and inflammation in db/db mice
To determine whether carnosine has the ability of reducing oxidative stress damage and inflammation, we measured the level of MDA, the activities of SOD, CAT, and GSH-PX and the levels of TNOS, iNOS, and cNOS. In Figure 2, the level of MDA was higher, and the activities of SOD, CAT, and GSH-PX were lower in the db/db mice. After carnosine treatment, the level of MDA was decreased, the activities of SOD, CAT, and GSH-PX were increased. As shown in Figure 3, in the brain of db/db mice, the level of pro-inflammatory factor iNOS was increased and the level of cNOS was decreased compared with db/m mice. After administration of carnosine, these changes were reversed. These data demonstrated that carnosine could reduce neuronal oxidative stress damage and the expression of pro-inflammatory factor in db/db mice.
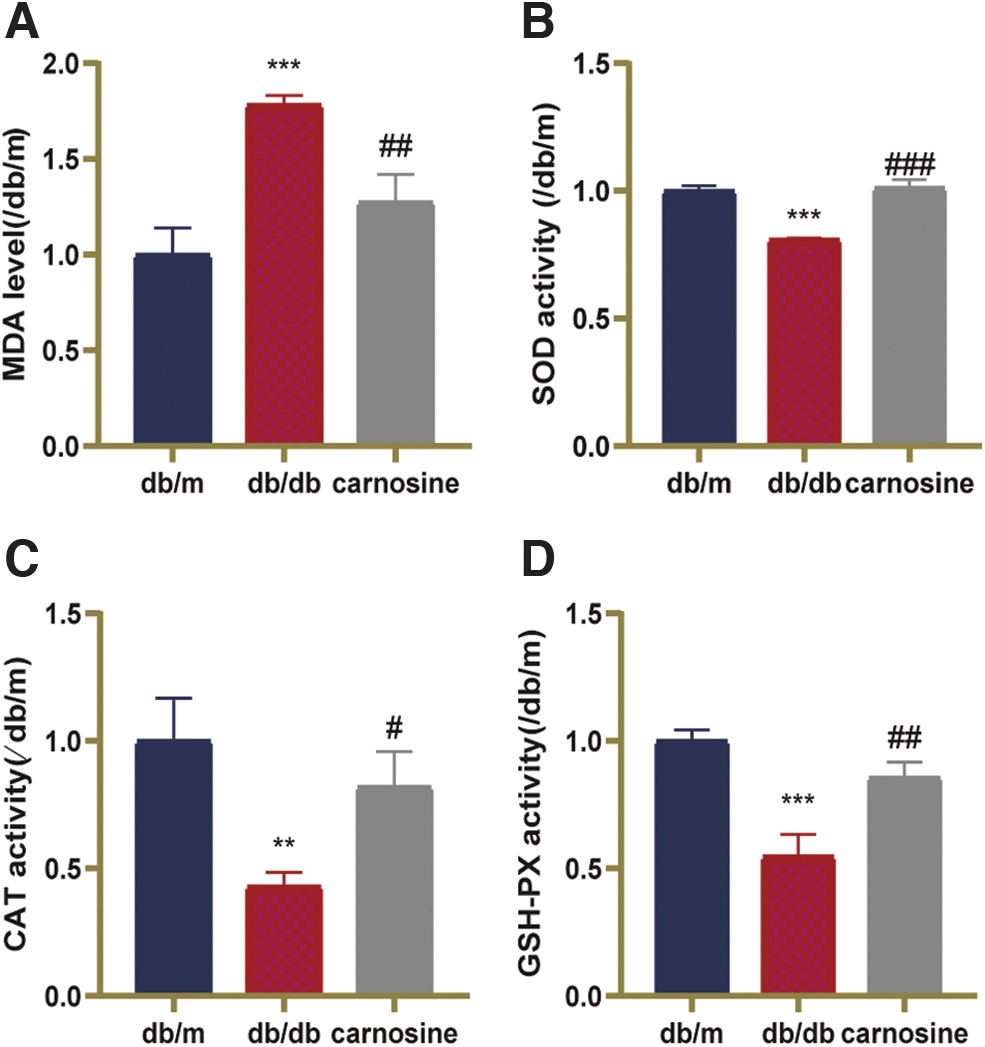
Carnosine reduces neuronal oxidative stress damage in db/db mice.

Carnosine reduces the expression of pro-inflammatory factor in db/db mice.
Carnosine reduces neurodegeneration in the hippocampus of db/db mice
To test the neuroprotection effect of carnosine, we detect the expressions of the synaptic marker, PSD95, and the neurotrophic factor, BDNF. The results showed that compared with db/m, the protein expressions of PSD95 and BDNF were significantly reduced in the db/db group (Fig. 4). Carnosine administration increased the expressions of PSD95 and BDNF. These data demonstrated that carnosine could reduce neurodegeneration in the hippocampus of db/db mice.
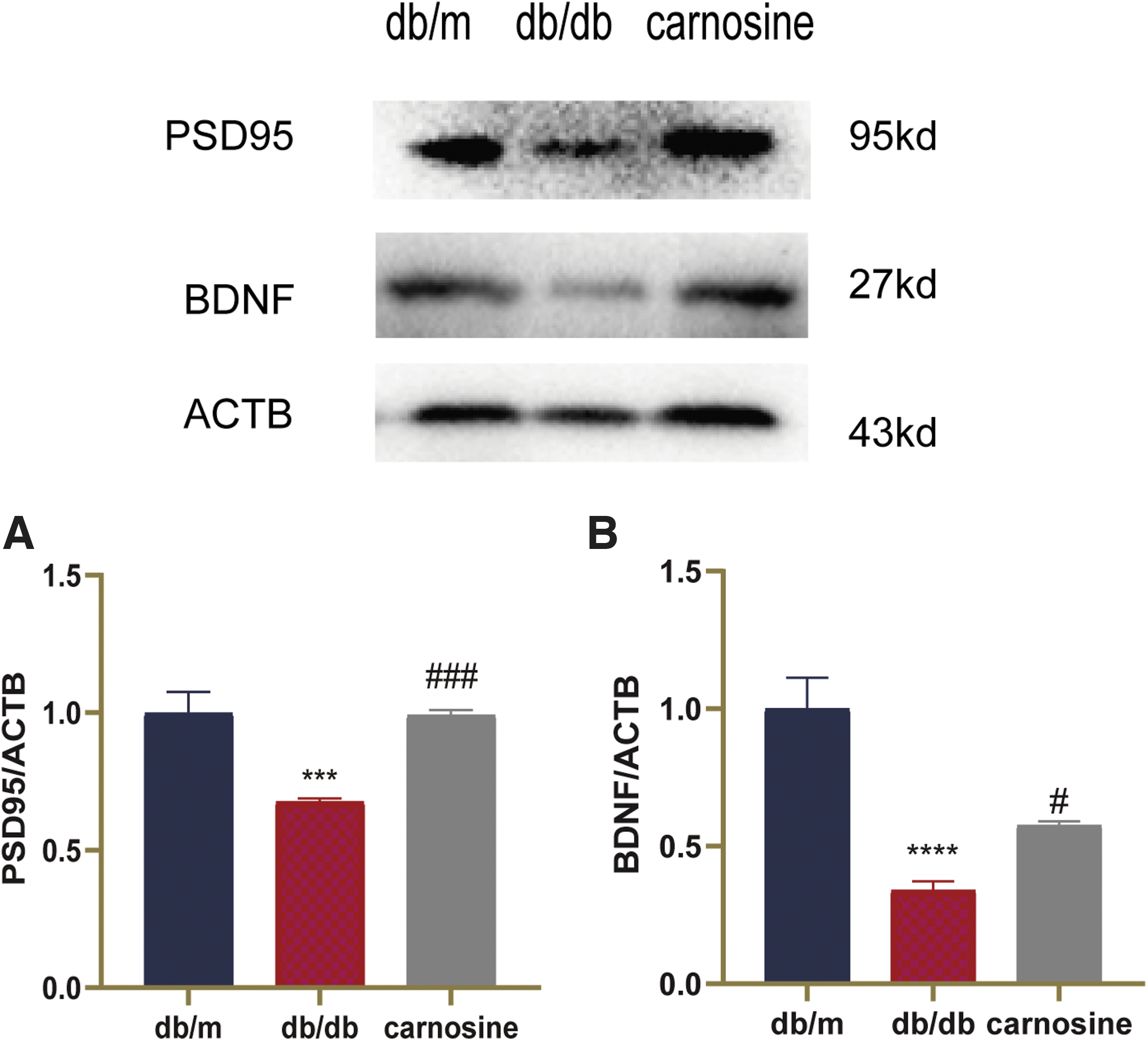
Carnosine reduces neurodegeneration in the hippocampus of db/db mice. Western blot analysis of target protein in the hippocampus of the mice.
Carnosine activates SIRT6 and relieves ER stress in the hippocampus of db/db mice
To determine whether SIRT6/ER stress pathway was involved in the protective effect of carnosine, SIRT6 and ER stress-related proteins were detected by western blot (Fig. 5). Results showed that the expression of SIRT6 decreased in the hippocampus of db/db mice. Whereas administration of carnosine increased the expression of SIRT6 in db/db mice. The ER stress-related proteins (BIP, P-PERK, P-IRE1α, ATF6, and CHOP) significantly increased in db/db mice. After carnosine treatment, the expressions of BIP, P-PERK, P-IRE1α, ATF6, and CHOP drastically reduced compared with the db/db group. Therefore, these data indicated that carnosine could activate SIRT6 and relieve ER stress in the hippocampus of db/db mice.
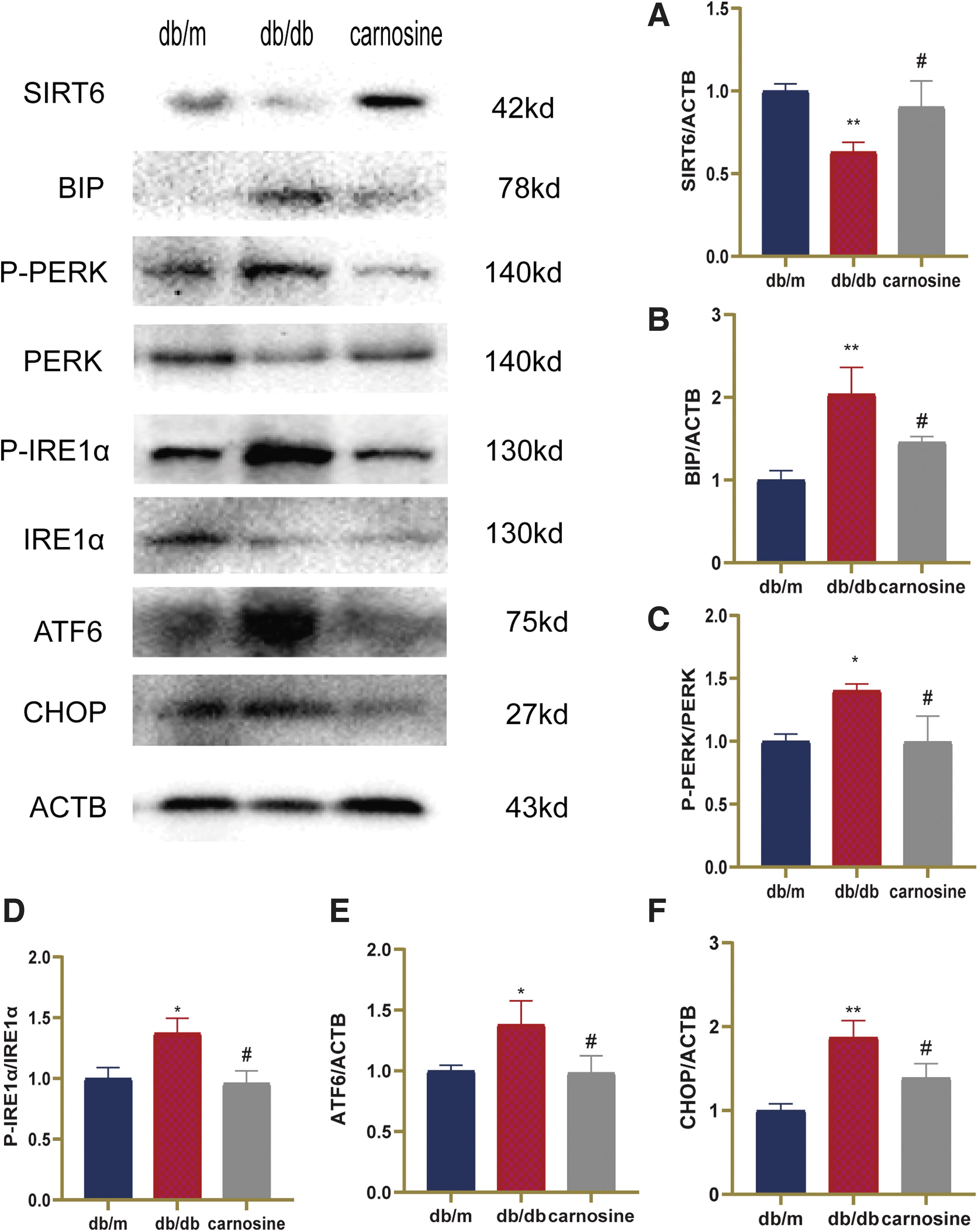
Carnosine activates SIRT6 and relieves ER stress in the hippocampus of db/db mice. Western blot analysis of target protein in the hippocampus of the mice.
Discussion
In this study, we found that carnosine could ameliorate cognitive impairment caused by diabetes mellitus in db/db mice. Eight-week administration of carnosine improved learning and memory dysfunction, relieved neuronal oxidative stress damage, and reduced the expression of pro-inflammatory factor in db/db mice. Meanwhile, carnosine reduced neurodegeneration in the hippocampus of db/db mice. Furthermore, carnosine activated SIRT6 and relieved ER stress in the hippocampus of db/db mice. Thus, our results demonstrated that carnosine could produce neuroprotective effect against DE, which might be related to SIRT6/ER stress pathway (Fig. 6).
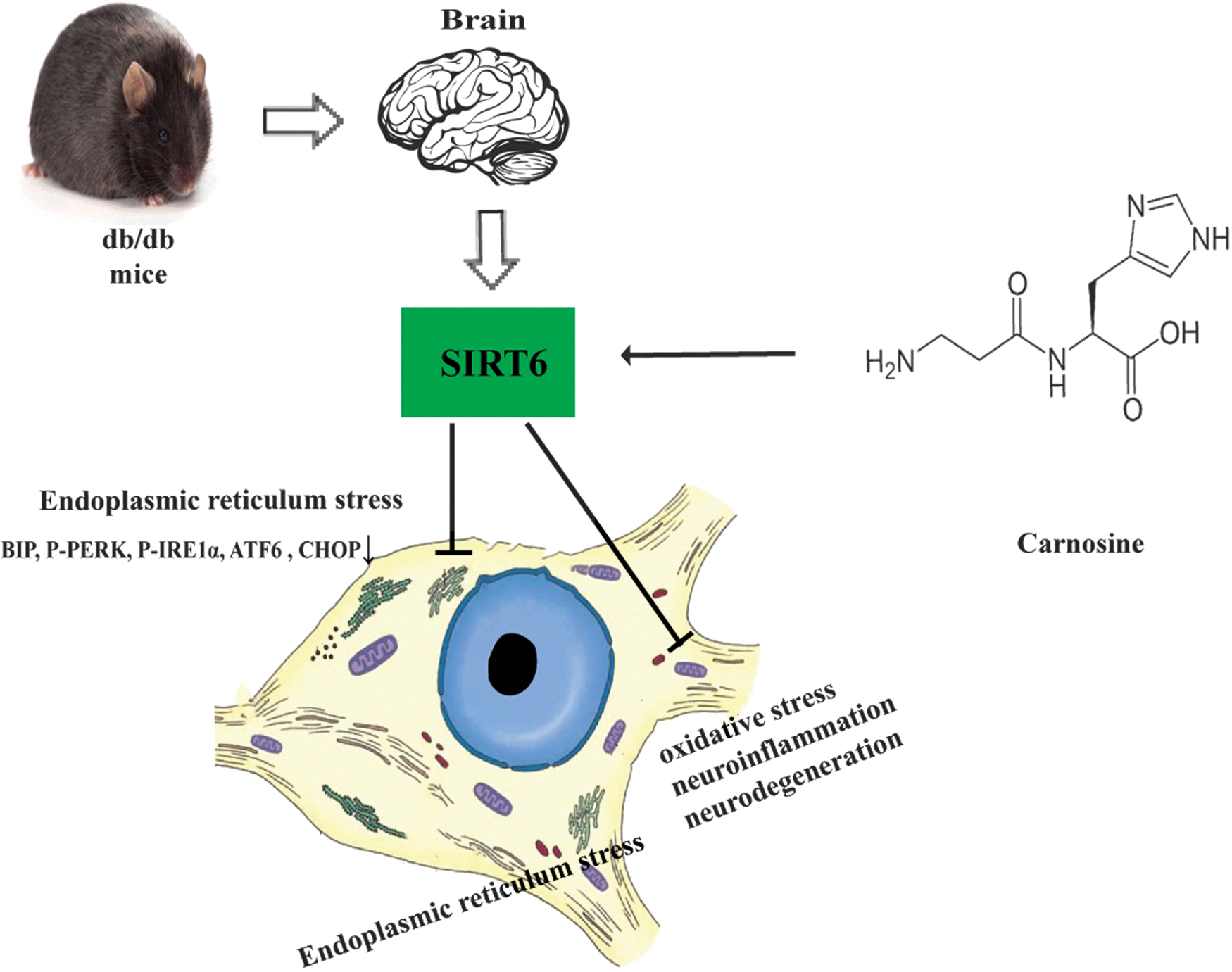
Carnosine can reduce oxidative stress, neuroinflammation, and neurodegeneration in the hippocampus of db/db mice by activated SIRT6 and relieved ER stress.
In the brain, insulin mediates multiple effects, such as glucose utilization and energy metabolism, oxidative stress, neurotrophic factor regulation, and cholinergic gene expression. 5,35 The weakening of insulin signaling inhibits synaptic plasticity in the hippocampus, leading to memory impairment. 36 The db/db mouse was used as a model of type 2 diabetes. Their phenotype consists of obesity, insulin resistance, diabetes, and cognitive impairment, which is similar to type 2 diabetes in humans. 37 Recently, carnosine has been used to treat cognitive impairment and diabetes mellitus. 38,39 In this study, cognitive impairment appeared in db/db mice, oral administration of carnosine significantly improved the learning and memory impairment in db/db mice. Therefore, we suggested that carnosine could ameliorate cognitive impairment in db/db mice.
Increased oxidative stress and the release of pro-inflammatory factors in the diabetic brain are considered to be important pathogenic factors for diabetic cognitive decline. 40,41 In the brain of diabetes mellitus, hyperglycemia enhances the release of pro-inflammatory factors and reduces antioxidant levels. 42,43 SOD, CAT, and GSH-PX are important antioxidant enzymes, which can effectively scavenge free radicals and resist free radical damage. 44 MDA, the product of lipid peroxidation, can indirectly reflect the level of oxygen free radicals in cells. 45 Studies have shown that inhibiting the expression of pro-inflammatory factor, iNOS, can alleviate neuronal apoptosis and cognitive impairment in diabetic animals. 46
In this study, neuronal oxidative stress and inflammation were occurred in the brain of db/db mice. Carnosine treatment reduced neuronal oxidative stress damage and the expression of pro-inflammatory factor in db/db mice.
Synaptic plasticity has been regarded as the basic neural mechanism of learning and memory. 47 Studies have shown that long-term potentiation in diabetic rats cannot be induced normally. 48,49 PSD-95 is crucial for synaptic plasticity. 50 BDNF, a member of the neurotrophic family, plays an important role in various neuronal activities, such as synaptic plasticity, neuron survival, neuron development, and differentiation. 51 The circulating BDNF levels of type 2 diabetes mellitus (T2DM) patients are reduced. 52 Interestingly, BDNF administration can significantly improve hyperglycemia and insulin sensitivity in T2DM mice. 53,54 In this study, the expressions of PSD95 and BDNF were significantly reduced in db/db mice. Carnosine administration reversed these changes. Therefore, carnosine could reduce neurodegeneration in the hippocampus of db/db mice.
SIRT6 plays an important role in diabetes mellitus. It can alleviate the pathological changes caused by diabetes by regulating the function of pancreatic β-cells, inhibiting gluconeogenesis, reducing adipogenesis, and preventing obesity. 19,55 Besides, study has shown that the expression of SIRT6 is reduced in the brain of db/db mice. 56 Our data also showed that the expression of SIRT6 was decreased in the hippocampus of db/db mice. The administration of carnosine increased the expression of SIRT6. Previous studies have shown that the pathological changes of hippocampal synapse and cognitive impairment in diabetes mellitus are related to ER stress caused by hyperglycemia. 17,57,58
When BIP and ER receptors (PERK, IRE1α, and ATF6) dissociate, this process regards as the beginning of UPR. 59 Three pathways of PERK, IRE1α, and ATF6 activate their downstream, CHOP, ER stress happened, and cell death. 13 Studies have shown that SIRT6 can deacetylate X-box binding protein-1 (XBP1) and inhibit the phosphorylation of PERK, which may help restore liver insulin sensitivity. 23,24 In this study, the ER stress-related proteins BIP, P-PERK, P-IRE1α, ATF6, and CHOP significantly increased in the hippocampus of db/db mice. Whereas carnosine administration decreased the expression of ER stress-related factors. Therefore, these data demonstrated that carnosine could activate SIRT6 and relieve ER stress in the hippocampus of db/db mice.
In conclusion, the neuroprotective effect of carnosine against DE was verified in db/db mice. In addition, it was found that carnosine ameliorated cognitive impairment by regulating the SIRT6/ER stress pathway in the hippocampus of db/db brain.
Ethics Approval
All procedures were approved by the Committee for Animal Research of Guangzhou University of Chinese Medicine.
Availability of Data and Materials
The data sets used and/or analyzed during this study are available from the corresponding author on reasonable request.
Footnotes
Authors' Contributions
S.-J.Z. designed the experiments. D.P. carried out the experiments and wrote the article. H.-Y.L. performed the animal study. L.G., L.-J.Q., and Y.-B.C. supported the materials. D.P., Q.X., and S.-J.Z. modified the article. Q.W. supported the experimental platform. Y.-F.C. and S.-J.Z. supervised this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by China Postdoctoral Science Foundation (No. 2021M690761); Natural Science Foundation of Guangdong Province of China (No. 2021A1515011478); Guangdong Provincial Key Laboratory of Research on Emergency in TCM (No. 2017B030314176).