
Abstract
Senile thymus atrophy is an important factor leading to decreased immune function. Repairing the atrophic thymus tissue structure, rebuilding immune function, and replenishing the number of exogenous stem cells may be ideal methods. In this study, bone marrow mesenchymal stem cells were intravenously infused into elderly macaques. We found that thymus volume was substantially increased, some thymus tissue regeneration was observed, the degree of thymus tissue fibrosis decreased, collagen fiber deposition decreased, cortical and medulla structures emerged gradually, the number of apoptotic cells decreased significantly, and the expression of apoptosis-related proteins decreased. For the effects of stem cell therapy on aging-related genes, we performed transcriptomic analysis of thymus tissue. The results show the expression pattern of the tissue transcriptome tended to be similar to the thymus expression pattern in young macaques compared with the elderly group, reverse aging-related proteins. Based on the results, it is suggested that stem cell therapy is an ideal method to prevent or reverse the aging of the thymus.
Introduction
The world is undergoing a rapid transition to aging. 1 Aging leads to the degeneration or loss of a series of functions at the level of tissues and organs. 2,3 The thymus is an important central immune organ of the human body. Degeneration of the thymus leads to weakened immune function, a decreased ability to resist infection, and an increased risk of tumors. 4 Therefore, the reconstruction of thymus tissue structure and immune function is necessary. 5
Since the end of the 19th century, more than 20 theories on aging have been proposed. 6 Based on the mechanism of aging caused by the decrease in the number of stem cells in the body, in this study, bone marrow mesenchymal stem cells (BMSCs) from juvenile rhesus monkey were infused intravenously into elderly rhesus monkeys to explore whether stem cells repair thymus tissue structure and function. BMSCs have become a popular trend in cell regeneration therapy due to their advantageous biological characteristics, such as easy access, easy amplification, low immunogenicity, and anti-inflammatory and immunomodulatory functions. 7,8
The rhesus macaque (Macaca mulatta), which is closely related to the evolution of humans, is a small primate model of human disease, development, and behavior that is widely used in infectious disease, pharmacology, and neuroscience research. In particular, the rhesus monkey model is an important tool for studying AIDS and the development of new drugs and vaccines against HIV. 9,10 Recently, some studies have described improvements in thymus function using stem cells. However, most studies were based on small and medium animal models, with few studies conducted in nonhuman primates. 11 Therefore, this study investigated the effects of BMSCs on the senescent thymus in aging rhesus monkeys. In addition, we investigated the molecular regulatory mechanism by which BMSCs reverse thymus senescence in rhesus monkeys. This study provides a theoretical basis for the treatment of thymus aging with BMSCs.
Materials and Methods
Experimental animals
Kunming Institute of Zoology, Chinese Academy of Sciences [2019-032 (Section)-01] provided 5 healthy female juvenile macaques with an average age of 3 years and body weight of 2.5 ± 0.5 kg and 10 healthy elderly female macaques with an average age of 25 years and body weight of 4.5 ± 0.5 kg. All macaques were housed in the animal room of the 920th Hospital of the PLA Joint Logistics Support Force. All aged macaques showed a senescence phenotype, dark hair color, limited mobility, obvious brain atrophy, pulmonary fibrosis, thymus, and ovarian atrophy. All animal studies were approved by the Laboratory Animal Ethics Committee of the supporting unit under animal certificate number: SYXK (Military) 2012-0039.
Preparation and identification of BMSCs from young macaques
Bone marrow was collected from healthy young female macaques with an average age of 3 years by bone marrow puncture under aseptic conditions. BMSCs were isolated and purified using the differential attachment method and the passage culture method. BMSCs were obtained after four passage amplifications. The morphology and growth status of BMSCs were observed under an inverted phase contrast microscope (Olympus, Tokyo, Japan).
Detection of the multidirectional differentiation ability of BMSCs from juvenile macaques
BMSCs were induced to differentiate into adipoblasts, osteoblasts, and chondroblasts in vitro, and the differentiation ability was detected. 12 P4 generation BMSCs were digested into single-cell suspension, inoculated at a density of 2 × 104 cells/cm2, added into the medium, and placed in an incubator with 5% CO2 at 37°C for culture until the cell growth and fusion reached >90%. Cells were incubated with adipogenic induction medium until the fat droplets become large and full; then, 1 mL of Oil Red O staining solution was added to each well, and cells were washed with phosphate-buffered saline (PBS) after staining.
When osteogenic induction medium was added and a large number of calcium nodules appeared under the microscope, 1 mL of alizarin red dye was added to each well at 14 days after induction, and cells were washed with PBS after staining. Chondrogenic induction medium was added to other cells. After 14 days, alcian blue staining solution was used for staining, and cells were washed with PBS. Adipogenic, osteogenic, and chondrogenic induction and differentiation were observed under an inverted phase contrast microscope, and cells were photographed.
Identification of surface antigens on BMSCs from juvenile macaques
The positive percentages of CD29 (lot no.: 11-0299-42), CD34 (lot no.: 11-0349-42), CD90 (lot no.: 11-0909-42), and CD105 (lot no.: 12-1057-42) cells were detected using flow cytometry; the above antibodies were purchased from eBioscience. P4 generation BMSCs were cultured. When the cells reached 80%–90% confluence at the bottom of the culture flask, 0.25% trypsin was added and cells were incubated in an incubator at 37°C. Rounding of the cells was observed under an inverted microscope. The cell density was adjusted to 1 × 107 cells/mL, and 100 μL of the cell suspension was separated into flow tubes. The cells were incubated at 4°C for 30 min in the dark, and the cell surface antigens were detected using a FACSCalibur flow cytometer.
Animal grouping and BMSC transplantation of macaques
The 15 rhesus macaques displayed no abnormal changes after 1 week of normal feeding and were randomly divided into groups, namely 5 in the juvenile group (n = 5), 4 in the elderly group (n = 4), and 6 in the elderly treatment group (n = 6). 1 × 107 cells/kg BMSCs were transplanted into the elderly treatment group via the femoral vein once every other day for three consecutive infusions; the juvenile group and the elderly group were infused with an equal volume of normal saline at the same time. Routine feeding was performed; the macaques in each group were sacrificed 6 months after the last transplantation, and materials were collected.
Hematoxylin–eosin staining of the thymus
The thymus tissue was fixed with 4% paraformaldehyde (PFA) solution for 24 h, dehydrated, embedded in paraffin, and sectioned at a thickness of 4 μm. Sections were dewaxed with xylene and washed with anhydrous ethanol. Sections were stained with hematoxylin, rinsed with dilute ammonia to develop the blue color, and washed with distilled water for 1 min. Eosin staining was performed, and sections were sequentially washed with 80% ethanol, 95% ethanol, and anhydrous ethanol. After incubation with xylene for dehydration, sections were sealed with neutral gum, air-dried at room temperature overnight, baked in an oven at 37°C, and observed and photographed under a light microscope.
Masson's trichrome staining of the thymus
The thymus tissue was fixed with 4% PFA solution, embedded in paraffin, and sectioned at a thickness of ∼4 μm. The paraffin sections were dewaxed to water and stained with ferric hematoxylin. After dehydration with anhydrous ethanol and dewaxing with xylene, sections were sealed with neutral gum and analyzed under a microscope.
Immunofluorescence staining of the thymus
The paraffin-embedded thymus tissue from rhesus macaques was sectioned at a thickness of ∼4 μm. Paraffin sections were dewaxed to water, subjected to antigen retrieval, and placed in ethylenediaminetetraacetic acid antigen repair buffer. Next, the primary antibody (Cytokeratin 5 [CK5], NBP2-22194; Novus) (Cytokeratin 8 [CK8], LS-C383073; LSBio) was added and incubated overnight in a humid chamber at 4°C. Sections were washed with PBS, and the secondary antibody for the corresponding species of the primary antibody was added dropwise to cover the tissues and incubated at room temperature in the dark. Sections were then stained with DAPI (4′,6-diamidino-2-phenylindole). After washes with PBS, the sections were sealed with anti-fluorescence quenching media. The slices were observed under an inverted fluorescence microscope, and images were captured.
Terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling staining of the thymus
The thymus tissues of rhesus macaques were embedded in paraffin and sectioned at a thickness of ∼4 μm. The paraffin sections were dewaxed to water, subjected to antigen retrieval, and then, the working solution of protease K was added dropwise to cover the tissues. The working permeabilization solution was added dropwise to cover the tissues, and the tissues were incubated at room temperature. Slices were incubated with a 3% hydrogen peroxide solution in the dark and then washed with PBS. The diaminobenzidine (DAB) chromogenic solution was added dropwise, and brownish yellow staining of the nucleus indicated a positive result. The nucleus was restained, and ammonia returned the color to blue. After dehydration, sections were sealed with neutral gum, a microscopic examination was performed, and images were captured and analyzed.
Immunohistochemical staining of the thymus
After section preparation, dewaxing, and rehydration, the tissues underwent antigenic recovery and immunohistochemical staining. The primary antibody (P53, P21, and Sirtuin3 from CST, China; Sirtuin1 from Affinity) (Caspase3 from Proteintech; Bax from Novus; Bcl-2 from Biorbyt, United Kingdom) was added to sections and incubated overnight in a refrigerator. The secondary antibody was added to sections and incubated at room temperature for 1 h. DAB chemical staining and hematoxylin reverse staining were subsequently performed. Sections were dehydrated with alcohol, rendered transparent using xylene, and sealed with neutral resin. Microscopy, image acquisition, and image analysis were then performed.
Transcriptomic sequencing analysis of thymus tissue gene expression
Five milligrams of thymus tissue from different groups of rhesus macaques was used for transcriptomic analysis. After grinding the thymus tissue, RNA was extracted for detection, and the library was constructed and tested for quality. Raw data were obtained by high-throughput sequencing, processed by disassembly of adapters, and quality-controlled to obtain clean reads. Differential expression analysis between the two comparison sets was performed using DESeq2 R software for determining differential expression in digital gene expression data using a model based on the negative binomial distribution. The resulting p-values were adjusted to control for false discovery rates using the method of Benjamini and Hochberg.
Genes with an adjusted p-value of <0.05 found by DESeq2 were assigned as differentially expressed genes. For each sequenced library, read counts were adjusted by the edge R package by a scale normalization factor. Using the Benjamini and Hochberg method to adjust the p-value, the corrected p-value and |log2foldchange| were used as the threshold for significant differential expression, and the aging-related genes were screened out using a linear model, and clustering and trajectory analysis were performed based on the aging-related genes. The Gene Ontology (GO) enrichment analysis of differentially expressed genes after BMSC transplantation was achieved by cluster Profiler R software, and GO terms with a corrected p-value of <0.05 were significantly enriched by differentially expressed genes. 13
Statistical analysis
All data were analyzed using SPSS 21.0 statistical analysis software. The statistical results are reported as
Results
Preparation and identification of juvenile BMSCs
Primary BMSCs showed colony growth, and the cells were evenly dispersed on the bottom of the culture plate. When the cell fusion rate reached 80%–90%, cells were subcultured and expanded. Following subculture, the P4 generation cells began to exhibit a uniform morphology, with fewer impurities, and grew in a long spindle shape resembling fibroblast-like cells, exhibited a high cell density, a whirlpool-shaped growth pattern, and good growth status (Fig. 1A). After in vitro adipogenic induction and staining with Oil Red O, the cell morphology became round, and red-stained lipid droplets were observed in the cell. Rhesus macaques BMSCs were induced to form osteoblasts in vitro. After staining with alizarin red, a large number of obvious dark red calcareous nodules were observed. Rhesus macaques BMSCs were also induced to form cartilage in vitro and stained with alcian blue; morphological changes were obvious, and proteoglycan synthesis in cartilage tissue was visible as blue staining (Fig. 1A).
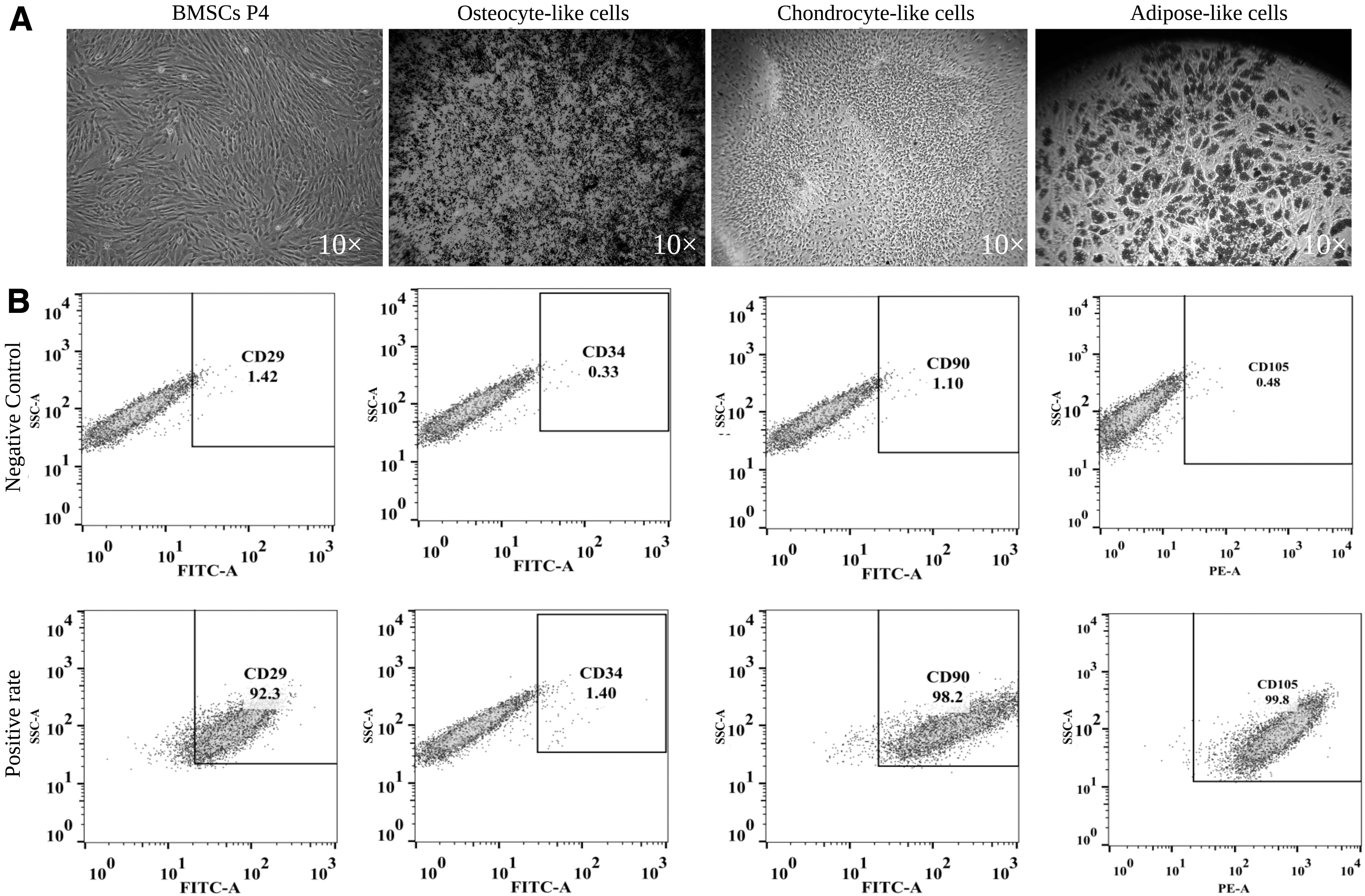
BMSC differentiation potential and molecular markers.
Flow cytometry analysis of the surface markers CD29, CD34, CD90, and CD105 on P4 generation BMSCs revealed positive expression rates of 92.3%, 1.40%, 98.2%, and 99.8%, respectively, consistent with the phenotypic characteristics of BMSCs (Fig. 1B).
Structural changes in the thymus
According to the observation of the appearance of thymus tissue, the thymus tissue of the juvenile group was fresh and full, whereas that of the elderly group was atrophic and deformed. After BMSC infusion, the thymus tissue volume of the elderly treatment group increased (Fig. 2A).

Structural changes in the thymus.
By hematoxylin––eosin staining, observed images from the juvenile group showed that the thymus tissue envelope was relatively intact, with a clear junction between the cortex and the medulla, and the parenchymal area of the thymus was large and full. In the elderly group of macaques, the thymus was significantly atrophied and degenerated, the parenchymal and medullary junction were blurred, and the cortical area was filled with adipose tissue. In the elderly group treated with BMSCs, an increased parenchymal area of the thymus was observed compared with the elderly group, the structure of the cortex and medulla areas improved and clear boundaries began to appear, the number of thymocytes increased, and the amount of fat tissue filling the thymus decreased (Fig. 2B).
Masson's trichrome staining of tissue sections showed that tissue in the juvenile group was arranged neatly and densely, the collagen fibers were blue, the area of collagen deposition was small, and the degree of fibrosis was low. With aging, a large number of collagen fibers appeared in the thymus tissue of the elderly group, the arrangement was disordered, the area of collagen deposition increased, and fibrosis was obvious. The area of collagen fiber deposition in the elderly treatment group was significantly lower than that in the elderly group, and the degree of thymic fibrosis was significantly reduced (Fig. 2C).
Structural reconstruction of thymus cortex and medulla
Based on immunofluorescence staining (Fig. 3), the cortex and medulla junction structure gradually appeared in the elderly treatment group, and the thymus structure was dense. The expression of CK5 was the highest in the juvenile group and the lowest in the elderly group, and the expression of CK5 in the elderly treatment group was significantly higher than that in the elderly group (p < 0.05) (Fig. 3A). The expression of CK8 in the young group was the highest, and the expression of CK8 in the old group was the lowest, and the expression of CK8 in the old treatment group was significantly higher than that in the old group (p < 0.01) (Fig. 3B). The highest expression of CK5+CK8 was observed in the juvenile group, and the lowest expression was detected in the elderly group. The expression of CK5 in the elderly treatment group was significantly higher than that in the elderly group (p < 0.05), and CK8 expression showed an increasing trend compared with the elderly group (Fig. 3C).
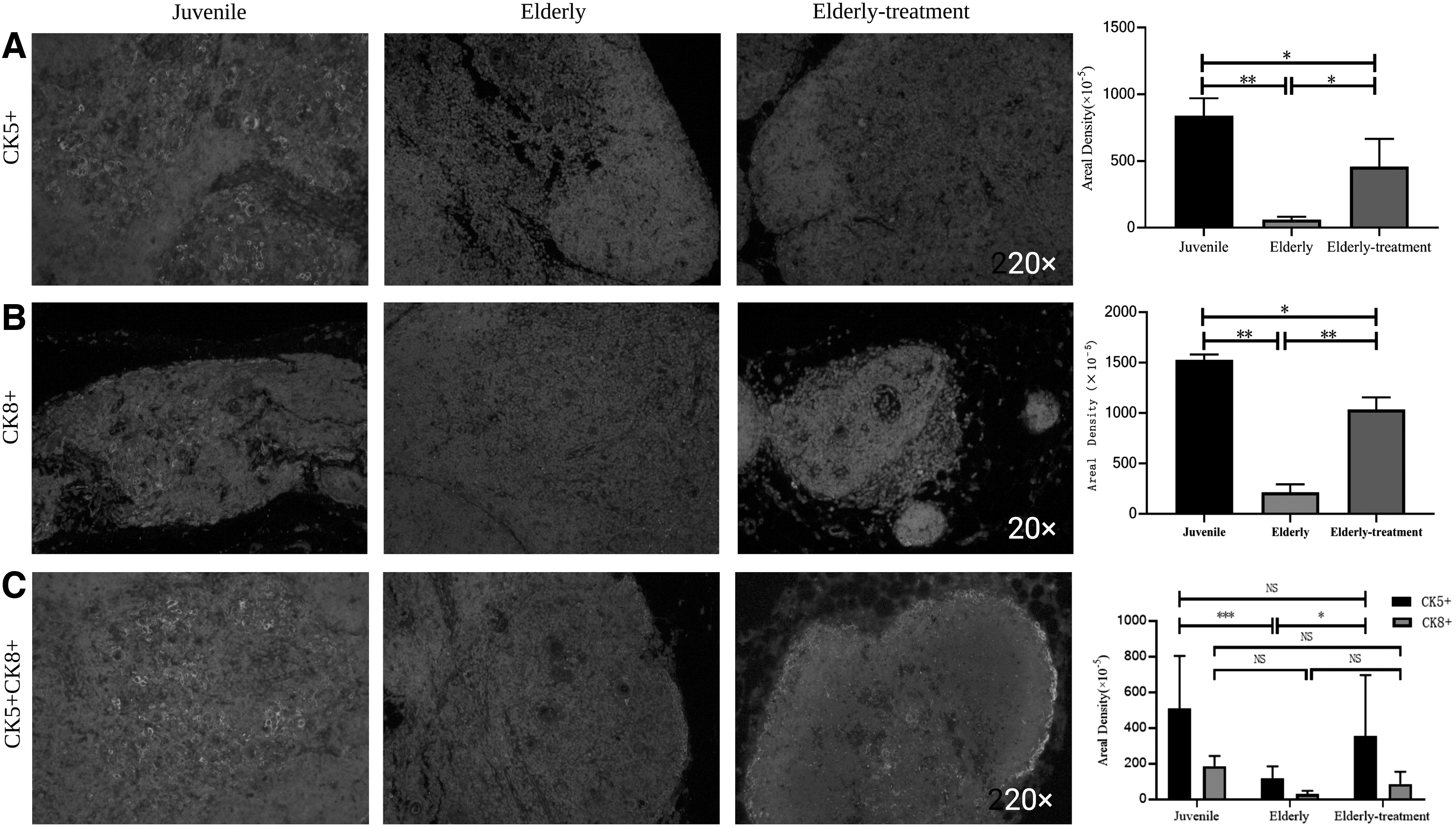
Structural reconstruction of thymus cortex and medulla. Immunofluorescence staining, structural changes in the thymic cortex and medulla of macaques.
BMSC transplantation reduced cell apoptosis
Terminal deoxynucleotidyl transferase-mediated dUTP nick end labeling staining of rhesus macaque thymus tissue (Fig. 4A) showed that the juvenile group had the least thymocyte apoptosis, the elderly group had the most thymocyte apoptosis, and the elderly treatment group had significantly fewer apoptotic cells than the elderly group. In addition, immunohistochemical staining results of apoptosis-related proteins showed that the lowest expression of the Caspase3 and Bax proteins was observed in the young group and the highest expression was detected in the elderly group (Fig. 4B–C). The elderly treatment group exhibited significantly lower expression of these genes than the elderly group (p < 0.01). The lowest expression of the Bcl-2 protein was observed in the elderly group, and the elderly treatment group presented higher expression than the elderly group (p < 0.05), and the expression was slightly higher than that in the juvenile group (Fig. 4D).
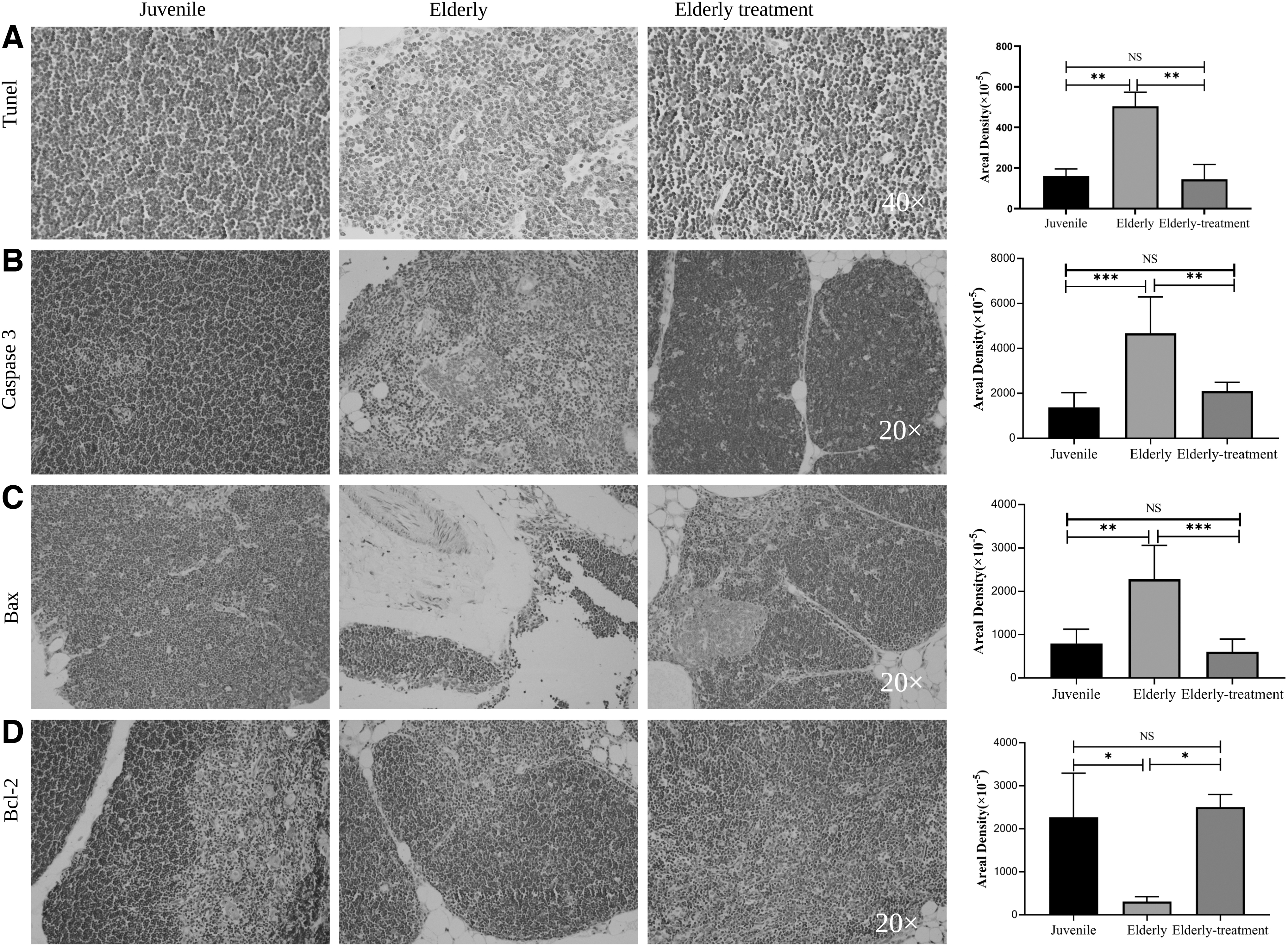
Apoptosis-related biomarker change.
Changes in thymus aging-related biomarkers
The lowest expression of the P21 protein was detected in the young group, and the highest expression was measured in the elderly group. The elderly treatment group exhibited significantly lower expression than the elderly group (p < 0.05). The expression of the P53 protein was the lowest in the young group and the highest in the elderly group. The elderly treatment group showed lower expression than the elderly group (p > 0.05). The highest Sirt1 protein expression was detected in the juvenile group, and the lowest expression was observed in the elderly group. The elderly treatment group exhibited a trend toward higher expression than the elderly group (p > 0.05). The Sirt3 protein was expressed at lower levels in the elderly group than in the young group (p < 0.05), and the elderly treatment group exhibited a trend toward higher expression than the elderly group (p > 0.05) (Fig. 5).
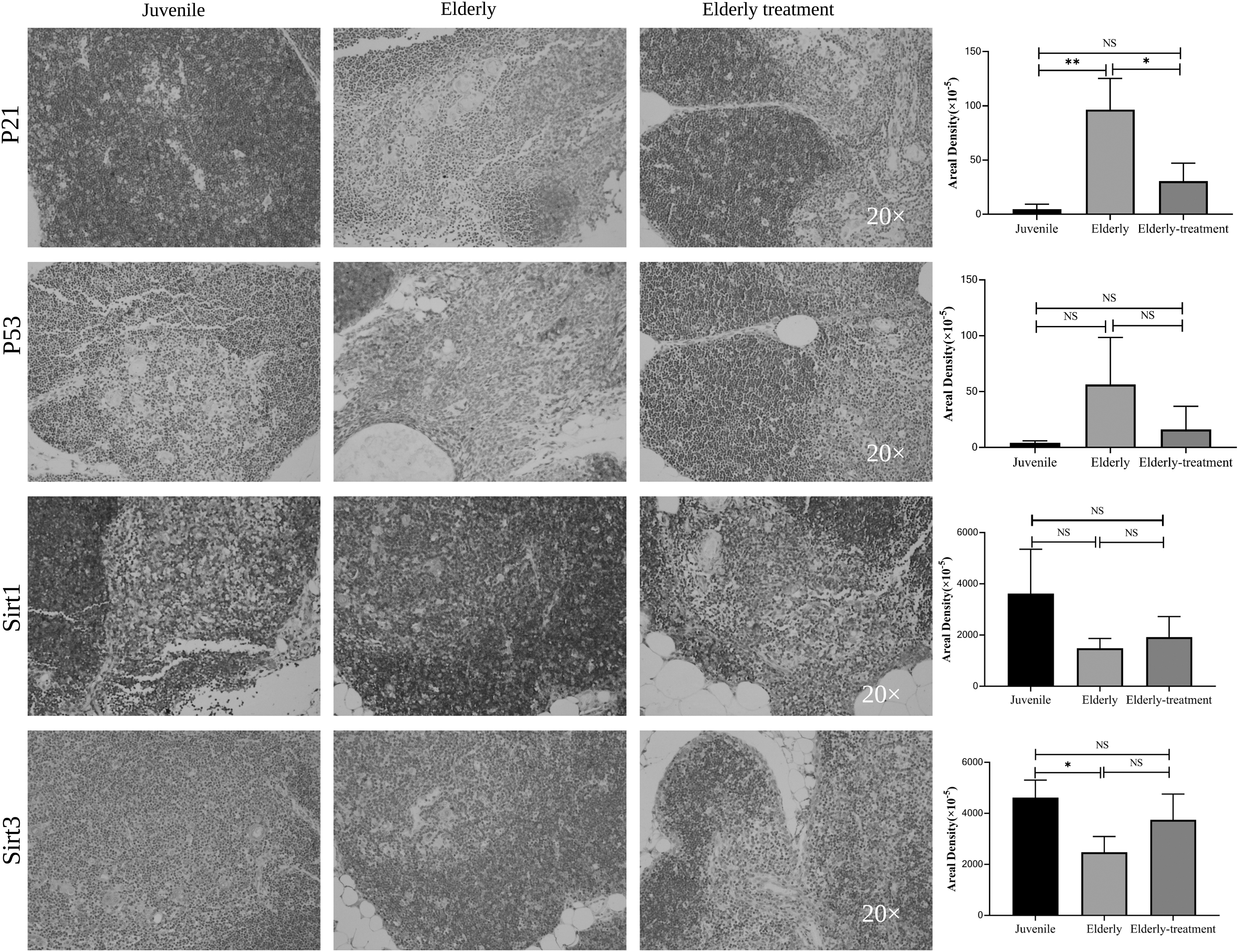
Changes in thymus aging-related biomarkers. For each thymus tissue, we performed immunohistochemical staining for aging-related biomarkers. P21-, P53-, Sirt1-, and Sirt3-positive expression was detected in multiple tissues using immunohistochemistry. Scale bars: 100 μm. Statistical method: one-way ANOVA. *p < 0.05, **p < 0.01, and NS p > 0.05.
The thymus reverses aging-related molecular mechanisms
Based on thymus senescence-related genes, 3D model principal component analysis of thymus tissue expression matrix results showed the overall transcriptional characteristics; the elderly treatment group (dark blue) were significantly more oriented toward the young group (light) than the elderly group (purple) (Fig. 6A). At the same time, cluster analysis was used to detect changes in thymus senescence-related gene expression before and after BMSC transplantation. The results showed that compared with the elderly group, the thymus expression characteristics of BMSC transplantation were more similar to that of young-adult macaques (Fig. 6B). Through a trajectory analysis, we determined the trajectory of thymic aging-related genes. Compared with the elderly group, the thymus expression patterns after BMSC transplantation tended to resemble the expression patterns in the thymus of the young rhesus macaque.
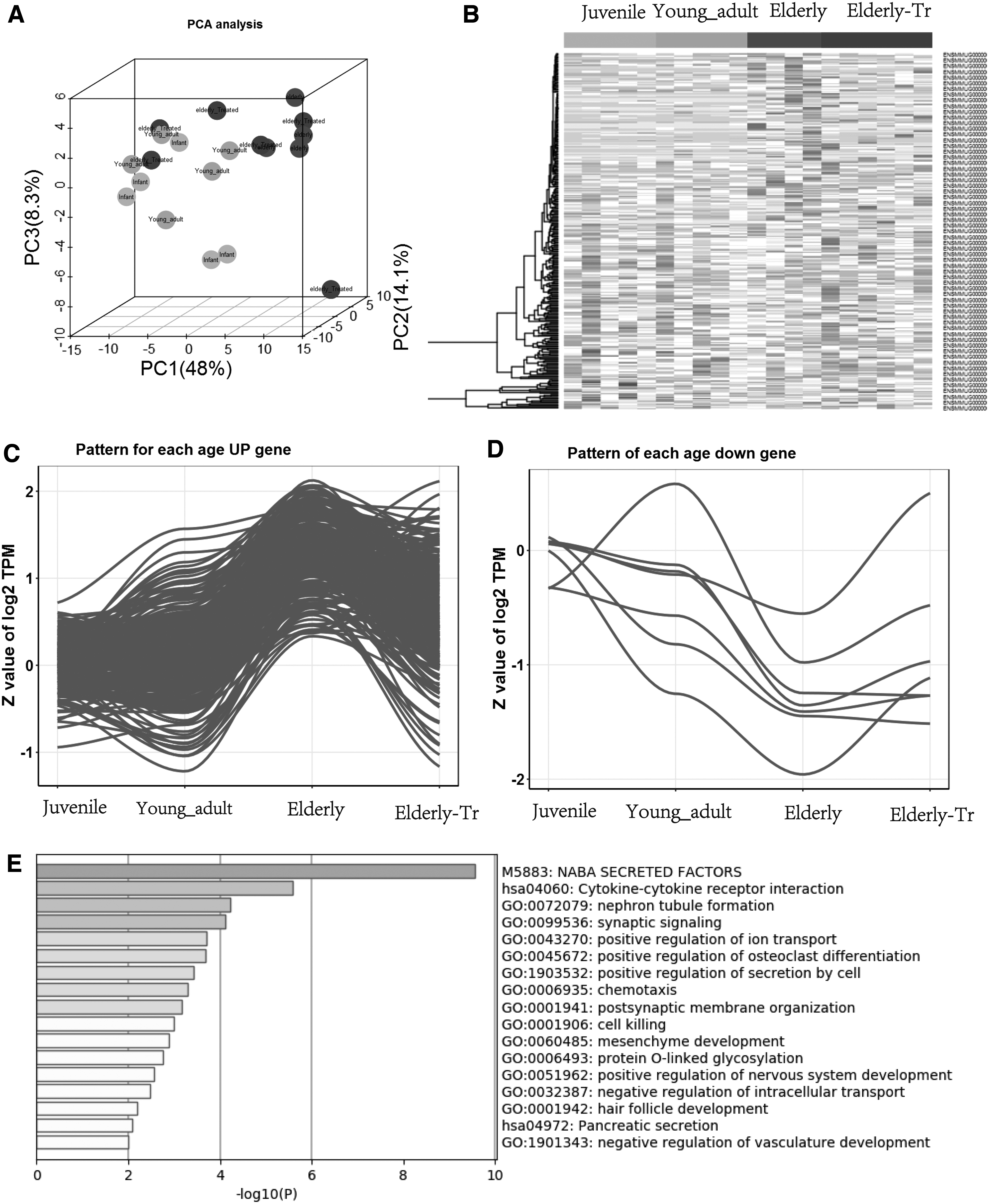
The thymus reverses aging-related molecular mechanisms.
Using linear model analysis, a total of 312 differentially expressed genes related to thymic tissue aging were detected based on p < 0.05, among which 305 genes were upregulated with age. Seven genes were downregulated with age (Fig. 6C–D). The GO enrichment analysis of genes that were upregulated with aging and decreased after treatment were sorted from large to small according to the -log p-value corresponding to each item to obtain 17 items. The GO enrichment analysis revealed that genes that were upregulated with aging and downregulated after treatment were mainly enriched in genes encoding secreted soluble factors (NABA), cytokine–cytokine receptor interaction, nephron tubule formation, synaptic signaling, positive regulation of ion transport, regulation of osteoclast differentiation, and other pathways (Fig. 6E).
Discussion
An important cause of aging is a series of functional and structural changes in the immune system. The thymus is the first immune organ to degenerate. 14 Elderly individuals are prone to infections, tumors, and autoimmune diseases due to degeneration of the thymus, resulting in immune senescence and reduced disease resistance. 15 In contrast, if the body can maintain a certain threshold of thymus hormone levels, it can rebuild and maintain the function of the earliest degraded organ in the human body, which may be used as an alternative therapeutic drug to treat and prevent many secondary diseases and achieve an antiaging effect.
Our previous study found that intravenous infusion of stem cells can improve the secretory function of the thymus. Based on the biological properties of stem cell immune regulation and anti-inflammatory and regenerative repair, this study used stem cell therapy to explore the structural and functional repair of the aging thymus. The structure of the thymus tissue was repaired, and biomarkers of thymus aging also changed. Finally, the transcriptome was used to reveal the mechanism of thymus function reconstruction. In summary, our findings may provide effective data for stem cell therapy of thymus-related diseases and molecular mechanisms.
When the thymus degenerates, its cortex and medulla are blurred and gradually replaced with adipose tissue. 16 After treatment with BMSCs, the senile rhesus macaques in the elderly treatment group exhibited an increased volume, cortex and medulla structures improved and boundaries began to appear, the number of thymocytes increased, and the amount of adipose tissue decreased compared with the elderly group. Organ fibrosis gradually increases with age. Organ fibrosis is the mechanism of repair of damaged parts of the organ itself, resulting in excessive secretion of extracellular matrix proteins and the formation of permanent scars, which ultimately leads to organ degeneration and functional failure. 17,18 We observed a large amount of collagen fiber deposition in the thymus tissue from the elderly group, whereas collagen fiber deposition in the treatment group was significantly reduced.
CK8 and CK5 are marker proteins for the cortex and medulla of thymic epithelial cells (TECs), so we used them to recognize the right location of TECs. In normal thymus, the K5 positive (K5+) TECs were observed in the center medullary, whereas the K8 positive (K8+) TECs were observed around the cortical areas. 19 We performed immunofluorescence staining to further confirm the structural changes in the thymus after treatment. The expression of CK5 and CK8 was significantly higher in the elderly rhesus macaque treatment group than that in the elderly group; BMSCs promoted the proliferation of thymus epithelial cells. 20
Similarly, the number of apoptotic cells in the treatment group was significantly lower than that in the elderly group. 21 We found that the levels of the apoptosis-related markers, Caspase3 and Bax, were significantly reduced in the elderly treatment group, 22 and Bcl-2 level was significantly increased in the elderly treatment group 23 and inhibit the expression of apoptosis-related proteins. Based on these results, BMSCs reconstructed the normal structure of thymus gland in aging rhesus monkeys. The results showed that BMSCs could repair the thymus structure of aging rhesus monkeys.
Thymus atrophy and decreased immune function are manifestations of aging, but the relationship between the thymus and aging is unclear. We tested aging-related proteins to reveal the mechanism by which stem cells repair the structure and function of the degenerated thymus. We found that the levels of the aging-related markers, P21 was significantly increased in the elderly treatment group, 24 and Sirt1 and Sirt3 levels were increased in the elderly treatment group. 25,26 To further confirm our conclusion, we performed a transcriptome sequencing analysis of thymus tissue. Based on p < 0.05, 312 differentially expressed genes related to thymic tissue aging were detected, which increased with age: 305 genes were upregulated, and 7 genes were downregulated with age.
By analyzing genes related to thymic aging, we further found that the elderly treatment group generally shifted to the pattern in the young group. By GO enrichment analysis, we found that genes that were upregulated with aging and downregulated after treatment were mainly enriched in cytokine–cytokine receptor interaction, nephron tubule formation, and synaptic signaling. The transcriptome results indicate that thymus degeneration and aging-related genes are regulated, and the repair of degenerated thymus is also inseparable from the expression of aging-related genes. These genes may provide references for precision clinical treatment of thymus-related diseases, and further exploration is needed.
In summary, juvenile BMSCs improve the structure and function of the thymus of elderly macaques. Stem cells improve the structure and function of the thymus, mainly by regulating aging-related genes and signaling pathways. Thymus degeneration and aging are inseparable, and the thymus may be the first beneficiary in the process of antiaging treatment.
Footnotes
Acknowledgments
Prof. Xing-Hua Pan and Prof. Xiang-Qing Zhu of the 920th Hospital of the PLA Joint Logistics Support Force and all the teachers are acknowledged for their assistance.
Authors' Contributions
Y.-Y.W., X.-Q.Z., and X.-H.P. designed the study. Y.-Y.W., L.C., Y.L., C.T., Q.W., and X.-Q.Z. performed the experiments. Y.-Y.W. analyzed the data. Y.-Y.W., X.-Q.Z., and X.-H.P. drafted and revised the article. All authors read and approved the final article for publication.
Availability of Data and Materials
All data generated or analyzed during this study are included in this published article.
Ethical Approval and Consent to Participate
Experimental protocols were approved by the Experimental Animal Ethics Committee of the 920th Hospital of the PLA Joint Logistics Support Force.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the project entitled “Transformation of subtotipotent stem cells based on the tree shrew model of multiple organ dysfunction syndrome” (SYDW[2020]19) and by Major Special Projects of Yunnan province (2018ZF007).