
Abstract
Our previous study showed that an exchange of blood between heterochronic parabionts for 3 months did not rejuvenate the immune system of the old partners. Moreover, the young immune system became more aged and began to function according to the “old” principle. Does this forced aging affect all systems of the organism in this model? We checked the levels of corticosterone, testosterone, insulin-like growth factor 1 (IGF-1), insulin, and thyroxine in the blood of heterochronic parabionts, but did not find significant changes compared with age-related controls. Since numerous data support the possibility of rejuvenation of the brain, muscles, and other tissues using the model of heterochronic parabiosis, as well as opposite data, we planned to assess the overall effect of this long-term blood exchange on the rate of organism aging. We measured the life span of animals whose blood was exchanged for 3 months and then were disconnected. Median and maximum life expectancy decreased in young heterochronic parabionts compared with the isochronic control. Old heterochronic parabionts showed only a small trend toward an increase in the median life span, but it was not statistically significant, and the maximum life span did not change compared with the isochronic parabionts. These data support our assumption that old blood contains factors capable of inducing aging in young animals. The mechanism of selective suppression of aging factor production in the organism could be a key research field for life extension.
Introduction
Heterochronic parabiosis is a widely used research tool to assess the effect of blood-borne factors/cells on functional recovery in aged animals. Parabiosis is a model of common circulation between two surgically connected animals, which was developed more than 150 years ago.
In 1864, Paul Bert first showed the presence of newly formed capillaries at the site of skin fusion—by introducing various substances into one animal and observing their appearance in a partner, 1 and this has been actively used for a long time. 2 During this time, the influence of heterochronic blood exchange on various systems and organs of animals was studied. However, the results depended heavily on the studied organ and term of coexistence.
In a number of studies that used the model of heterochronic parabiosis, blood factors from young blood rejuvenated the cells and tissues in old animals. Such results were obtained in the studies of the functions of satellite–liver progenitor cells as well as neural stem cells, 3,4 skeletal muscle regeneration, 3 scar formation and skin regeneration, 5 vascular remodeling in the brain and olfactory discrimination in aging mice, 4 age-related heart hypertrophy, 6 thymus regeneration, 7 bone repair, 8 and kidney functions. 9,10
At the same time, numerous publications have shown that heterochronic blood exchange can have mutually opposite effects—to rejuvenate the functions/age markers of old animals, and contrariwise—to induce senescence in young ones. The study of transcriptome landscapes of different tissues after heterochronic blood exchange showed changes for different cell types. For example, mesenchymal stromal cells, endothelial cells, hematopoietic stem cells, and hepatocytes were most susceptible to changes in gene expression after 5 weeks of heterochronic parabiosis.
Moreover, the transcriptomic effects were opposite—aged blood, as a rule, accelerates age-related changes in young partners, while young blood simultaneously rejuvenates old ones. 11 Similar results have been found for visceral adipose tissue, 12 metabolic changes, and oxidant stress in mouse red blood cells 13 and in several other studies.
Another block of studies showed that heterochronic blood exchange predominantly accelerated aging effects in young partners. These studies demonstrated that reciprocal blood exchange leads to significantly lower total mitochondrial content and significantly smaller mitochondria in muscles compared with young isochronic controls 14,15 and accelerated aging of immune functioning in young heterochronic partners without any changes in old ones. 16,17
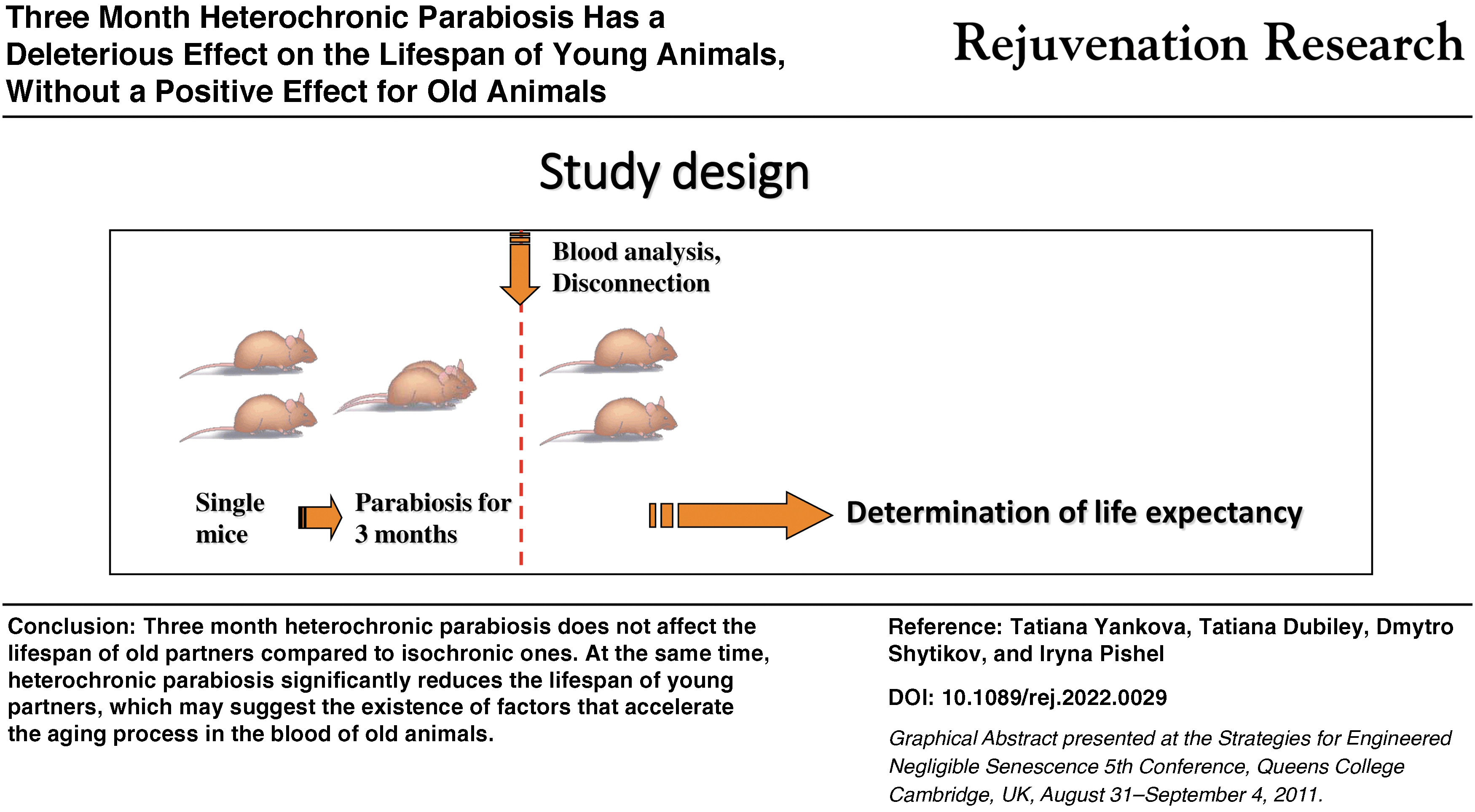
Indeed, the mammalian organism is a complex self-regulating system, in which intervention can lead to multidirectional results. Do we really register the processes of rejuvenation or accelerated aging on the model of heterochronic parabiosis? To answer this question, we conducted a study of the life span of parabionts after a 3-month reciprocal blood exchange and subsequent disconnection.
Materials and Methods
Study design
We used a generally healthy common strain of long-lived mice, CBA/Ca. Male mice, aged 2–3 (young) and 18–19 (old) months, were randomly divided into six groups of equal age distribution and size and underwent surgery to generate sham-operated single animals or parabionts. The experimental groups included young single (sham operated) mice (n = 20), old single (sham operated) mice (n = 25), young isochronic parabionts (n = 44; 22 pairs in total), old isochronic parabionts (n = 56; 28 pairs in total), and young and old heterochronic parabionts (n = 45 for both; 45 pairs in total).
Three months after initiation of parabiosis, mice were disconnected, and 5–10 pairs were subjected to blood plasma analysis. The rest of the animals were used to estimate life span. Mouse survival was checked every 1 to 3 days.
Parabiosis
Male CBA/Ca mice were bred and maintained at the D.F. Chebotaryov State Institute of Gerontology Animal Facility. For parabiosis, animals were typically joined at 2–3 (young) and 18–19 (old) months of age. The parabiosis surgery was performed exactly as described previously 16 and in accordance with the guidelines established by the D.F. Chebotaryov State Institute of Gerontology Administrative Panel for Laboratory Animal Care. Control (single) mice were sham operated in the same way.
The mice were housed in standard cages, each cage containing ∼2 to 3 pairs, which included one isochronic and two heterochronic pairs. Disconnection of mice was performed according to a standard procedure, which included disconnection of the peritoneal wall, clavicle, and skin of parabionts, followed by restoration of the integrity of the abdominal cavity and the skin of each individual animal.
Ninety-four percent of animals survived the surgery, on average, during the 1st week after surgery and it was only somewhat lower in the group of old mice (91%). Animals that died after separation surgery during the 1st week were not included in the study. Mice received standard rodent diet and acidified water ad libitum. Animals were maintained in a good conventional environment, with temperature of 21 ± 2°C and 12-hour light–12-hour dark cycles.
Clinical chemistry and enzyme-linked immunoassay
Blood collection was carried out after 6-hour fasting (from 8 am to 2 pm). Terminal bleeding and euthanasia for a part of the experimental animals were performed after 3 months of parabiosis. Animals were injected intraperitoneally with 2,2,2-tribromoethanol (250 mg/kg). Blood was collected from orbital sinuses into dry microtainers without an anticoagulant.
For serum collection, the tubes with collected blood were kept at room temperature for 15–60 minutes after blood drawing to form a fibrin clot and then centrifuged at 9000 rpm for 20 minutes at +4°C. The collected serum was frozen within 60–80 minutes after blood drawing and stored at −20°C for further clinical chemistry analysis.
The levels of some hormones in the serum of experimental animals were measured by enzyme-linked immunoassay according to the manufacturer's manual—total thyroxine (TT4; Diagnostic Systems, RF), insulin (ALPCO Diagnostics, USA), insulin-like growth factor 1 (IGF-1) (IDS, Great Britain), corticosterone (DRG, Germany), and testosterone (DRG). Glucose was determined in the serum using commercial kits (BioSystems, Spain) according to the manufacturer's instructions.
The levels of total cholesterol, triglycerides (TG), and high-density lipoproteins (HDL) were determined in the serum using commercial kits (Sentinel Diagnostics, Italy) according to the manufacturer's instructions.
Statistical analysis
Statistics were calculated using RStudio (v1.4.1717, RStudio Team 2021). Numerical parameters were analyzed through two-way factorial analysis of variance (ANOVA), with age and type of parabiosis as experimental factors, using the following packages: car, agricolae, and Fisheries Stock Assessment (FSA) (versions 3.0–12, 1.3–5, and 0.9.3, respectively).
Before the ANOVA, parameters were tested for normality using the Shapiro–Wilk test and equity of variance using Bartlett's test. If assumptions for parametric ANOVA were not violated, we used the factorial ANOVA, followed by the Newman–Keuls post hoc test. If the assumptions were violated, we used the Kruskal–Wallis nonparametric ANOVA, followed by Dunn's post hoc test.
Animal survival was analyzed by Kaplan–Meier plots using survival, survminer, and ggplot2 packages (versions 3.2–13, 0.4.9, and 3.3.5, respectively). The instant hazard rate was determined using the epiR package (version 2.0.43). Differences between groups were confirmed using the Mantel–Haenszel log-rank test. After confirming the presence of differences between the groups, we performed pairwise comparisons of all groups with each other using the survival package.
We used two-way factorial ANOVA with age and type of parabiosis as experimental factors to compare the average and maximal life span of animals. After confirming differences between groups, the ANOVA was followed by the Newman–Keuls post hoc test. To calculate the maximal life span, we chose 10% of the longest living mice (n = 3–5 mice in each group).
Results
In the first step, we studied the serum levels of hormones in experimental animals to determine possible systemic factors that may affect the life span and induce/restore age-related changes. We chose male CBA/Ca mice in continuation of our previous work, which revealed significant changes in the functioning of the immune system in parabiotic mice of this strain. 16
We selected several hormones for analysis, which are important for age-related disorders 18 : corticosterone, testosterone, insulin, IGF-1, TT4, total cholesterol, TG, and HDL. It should be noted that animal pairing for parabiosis leads to significant damage of the skin, abdominal muscles, and, in part, the bones of the scapula of experimental animals. Complete healing after surgery lasts 5–14 days for different tissues and results in chronic stress in the operated animals for that period of adaptation.
As longevity studies require a period not confounded by ongoing stress, we tested corticosterone levels at the end of the coexistence period of parabiosis. The results of the study are shown in Figure 1. The data on corticosterone concentration in the serum are not significantly different. Isochronic parabionts tend to have somewhat higher values, and young single mice have relatively lower values. However, the effects are not statistically significant (Fig. 1D).
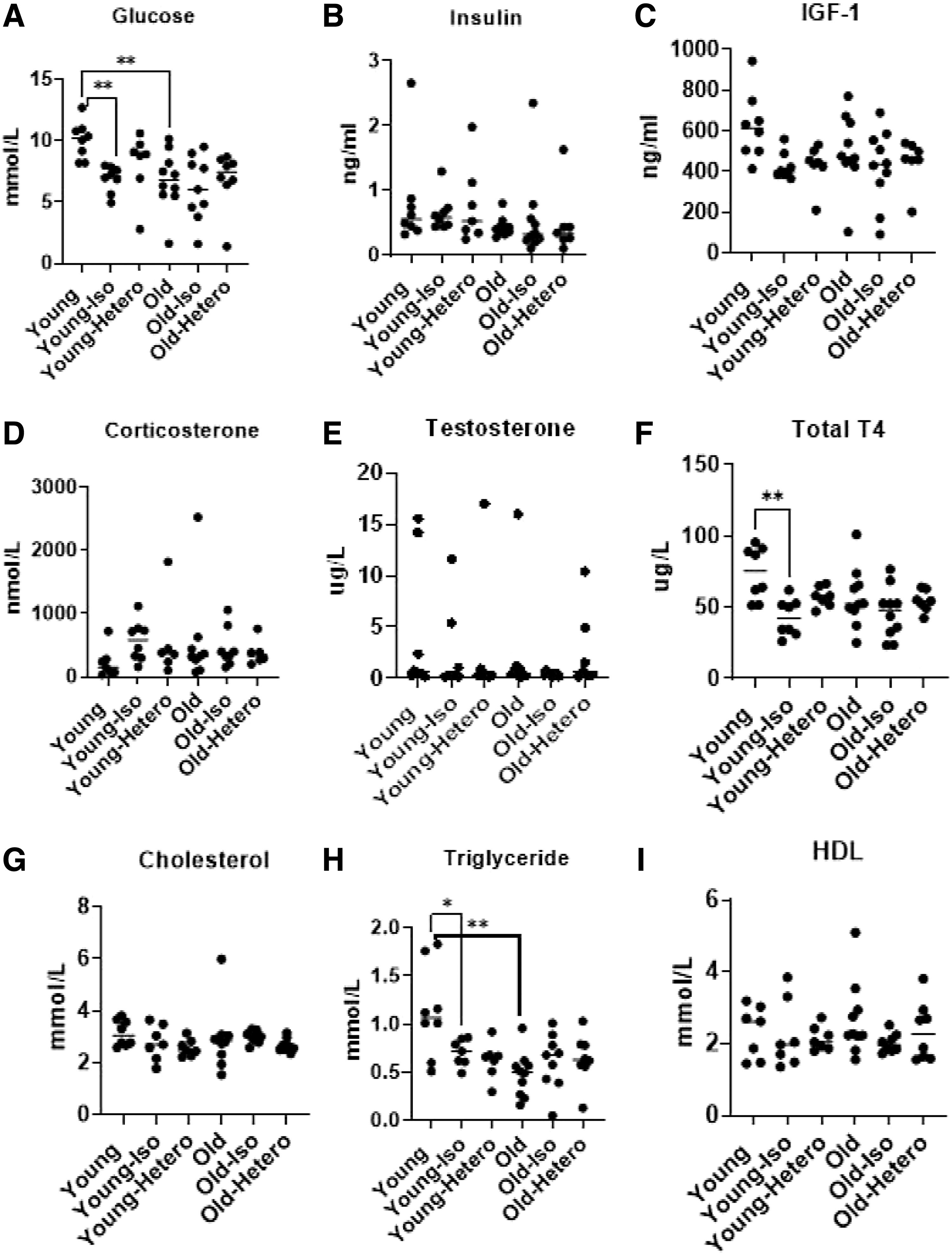
The levels of glucose
According to another investigation, the stress associated with the parabiosis surgery procedure, as well as general health, activity, and mouse behavior, normalizes 4 weeks after the operation. 19 In the current study, we found no difference in corticosterone levels 3 months after the surgery, both between old and young isochronic pairs and between old and young heterochronic partners compared with isochronic ones (Fig. 1D). Therefore, the effect of corticosterone should be comparable among the parabiotic cohorts and it could not have had a significant impact on differences in their physiological state.
After 3 months, body weight was less in both young and old isochronic partners compared with single age-related animals. At the same time, this parameter did not differ in heterochronic partners compared with isochronic ones (Table 1). Thymus weight tended to be less in young parabionts compared with single animals [p(t) = 0.1], but did not change significantly in old parabionts. Testicle weight tended to be lesser in young heterochronic parabionts compared with isochronic ones [p(t) = 0.054], but the opposite effect was observed in old heterochronic parabionts—testicle weight was significantly increased compared with old isochronic parabionts (Table 1).
Body, Thymus, and Testicle Weights in Experimental Animals 3 Months After Surgery
Values are given as mean ± SEM.
p(Z) < 0.01 compared with young single animals.
p(Z) < 0.01 compared with old single animals.
p(Z) < 0.05 compared with old heterochronic parabionts.
§p(Z) < 0.01 compared with old isochronic parabionts.
p(t) <0.01 compared with old single animals.
p(t) < 0.06 compared with young heterochronic parabionts.
p(t) < 0.01 compared with old isochronic parabionts.
p(t) < 0.01 compared with old heterochronic parabionts.
SEM, standard error of the mean.
We observed decreased levels of glucose and TG in old single animals and in isochronic and heterochronic parabionts, compared with young single animals (Fig. 1A, C, F, H; p < 0.01). Therefore, these parameters of energy and metabolism are changed by parabiosis and the age of the mouse itself, but not attributed to the effects of heterochronic parabiosis. The blood level of the growth hormone, IGF-1, showed a similar trend, where young single mice have high values, while old single mice and parabionts of all age combinations have similar lower values (Fig. 1C).
However, the effect is relatively weak as young single mice differ from the other groups with only p < 0.1. Total thyroxine measures thyroid function and showed a significant drop in young mice from parabiosis, regardless of partner age. Parabionts and old mice had similar lower levels of this hormone (Fig. 1F). The mechanism of these changes is still not well understood, but may indicate a significant effect of the parabiotic coexistence on hormonal levels in the animals, perhaps from changes in activity or eating behavior.
For other parameters, such as the concentration of insulin, testosterone, cholesterol, and HDL, any influence of the age of mice and type of parabiosis was not statistically significant (Fig. 1A–I).
The survival curves beginning 1 week after surgery as well as survival characteristics are presented in Figure 2 and Table 2. Keep in mind that the time on the x-axis of Figure 2 represents the recorded days after separation from surgery, not animal age in days. Young animals are already 165 ± 15 days old at the start of the curves, while old animals are already 645 ± 15 days old.
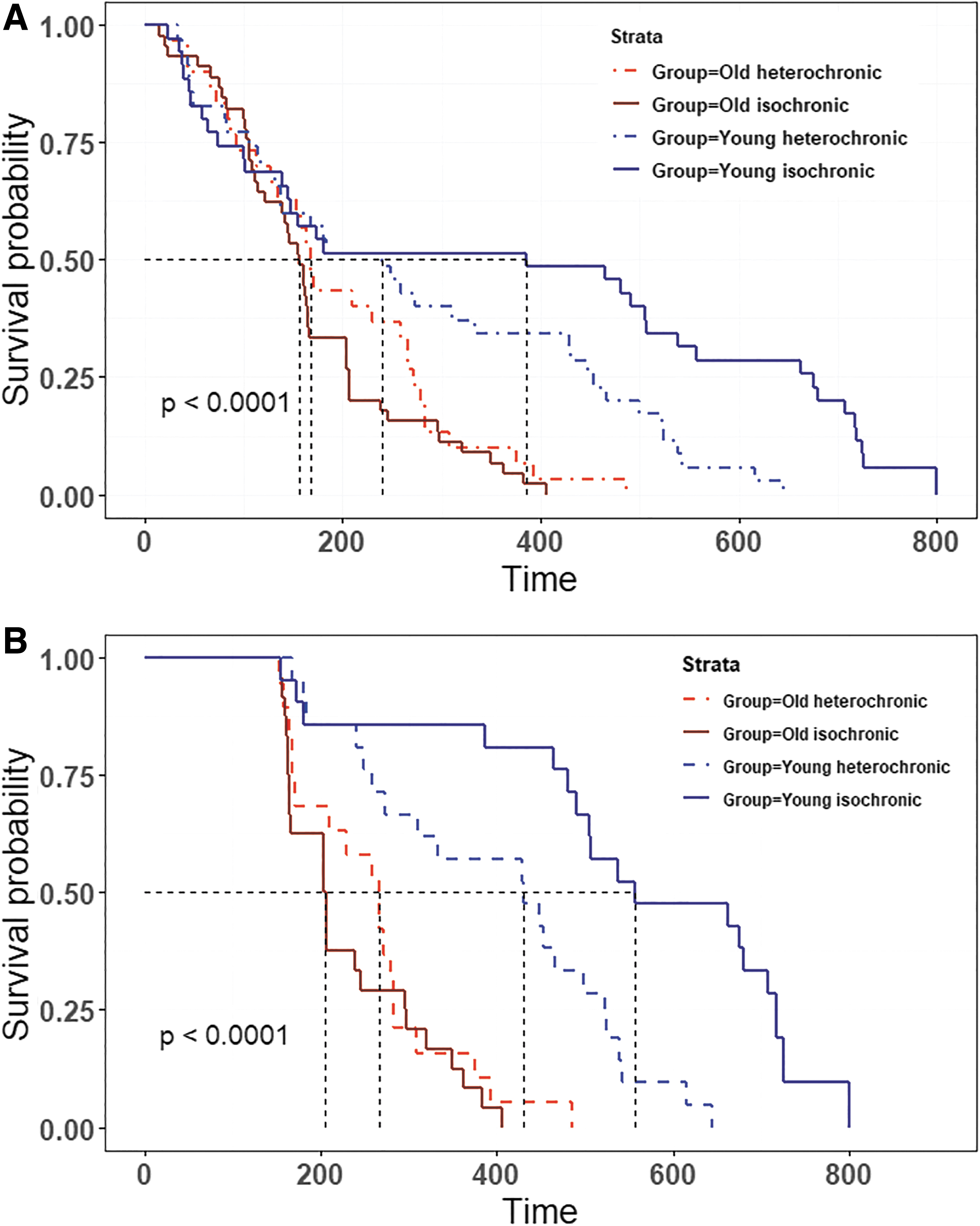
The survival rate (life span in days after disconnection) of male CBA/Ca young and old isochronic and heterochronic parabionts for total mice
Life Span Statistics of the Experimental Animals
Values given are days of animal age. Mean and maximal life span as well as 25th, 50th, and 75th percentiles are presented.
p(t) < 0.01 compared with young isochronic parabionts.
p(t) < 0.01 compared with young single animals.
p(t) < 0.01 compared with young heterochronic parabionts.
Heterochronic parabiosis for 3 months did not significantly increase the life span of old heterochronic parabionts compared with old isochronic ones (p χ2 > 0.1; Fig. 2A). Moreover, heterochronic parabiosis significantly decreased the life span of young heterochronic parabionts compared with the young isochronic ones (p χ2 = 0.01; Fig. 2A).
Survival of mice was not different during the first 150 days, which might be attributed to the recovery of animals from separation surgery. To remove the effects of surgery in our analysis, we decided to have a closer look at mice that survived over 150 days (Fig. 2B). Again, we observed a highly significant reduction in the life span of young heterochronic parabionts compared with young isochronic parabionts (p < 0.005; Fig. 2B) without any significant increase in the life span of old heterochronic parabionts.
Assessment of the average life span using factorial two-way ANOVA revealed a strong effect of age in mice (F (1, 141) = 19.45, p < 0.001); there was evident, although statistically not significant, interaction of age with the type of parabiosis [F (1, 141) = 3.58, p = 0.06]. The mean life span of young heterochronic parabionts was significantly lower than that of young isochronic parabionts (p t < 0.05). The mean life span of old heterochronic parabionts was not significantly increased due to the contact with young animals.
The maximal life span of animals was affected by the age of mice [F (1, 12) = 146.2, p < 0.001], type of parabiosis [F (1, 12) = 6.98, p < 0.05], and interaction of these two factors [F (1, 12) = 24.33, p < 0.001]. Old mouse cohorts did not differ significantly from each other and their life span was significantly longer than in any type of young animal (p t < 0.001; Table 2). At the same time, it was significantly lower in young heterochronic parabionts than in young isochronic ones (p t < 0.001; Table 2).
Discussion
Parabiosis is a complicated model of the anatomical and physiological union of two organisms. Numerous studies have used this model, but there are still many questions to solve. As mentioned above, parabiosis is generally accepted as a model of blood exchange between partners through an extensive network of capillaries that are formed during regeneration of peritoneal muscle and skin after surgery.
However, more detailed studies showed that the transfer of cells and biologically active substances with blood has limitations for parabiosis and the physiological effects can be observed not only due to blood factors but also due to the adjacent tissue mutual reinnervation, 20 –22 as well as the influence on the size–control mechanisms of internal organs and their functioning. 23,24 These aspects are experimentally unexplored in parabionts and require further elucidation, especially when using old animals.
The dynamics of blood flow and the distribution of leukocytes between parabionts have been explored in detail by many researchers. In particular, Gibney et al. showed that all populations of neutrophils and lymphoid cells reach equilibrium in peripheral blood within 14 days. Fluorescence analysis of whole blood showed an average rate of 16 μL/hour or 0.66% of the circulating blood volume per hour. 25
At the same time, migration of lymphocytes into tissues (lymph nodes and spleen) showed variable distribution: equilibrium occurred at 14 days for most studied cell types, but some populations, such as B220+ and CD4+ cells, had not yet reached equilibrium in lymphoid tissues even after 28 days. 25 This factor was the main prerequisite for our choice of longer-term parabiosis—up to 3 months, which would provide a more complete mutual distribution of cells in partners' tissues.
Migration or transplantation of young lymphoid cells and stem cells has always been of interest in aging research. Using the parabiosis model, we observed an active exchange of lymphocytes between partners, which mutually repopulate the lymphoid organs, 17 while donor cells usually did not cross the blood–brain barrier in parabiosis. 26 Repopulation of muscle tissue by donor cells in the parabiont mdx muscle has also been shown. 27
However, the donor cells did not express myosin heavy chain, which indicates that they do not participate in muscle regeneration. 27 Liver tissues show similar effects when resident stem cells constitute the major source of stem cells. 28 However, most authors suggest that migrating cells can have an indirect effect on tissue remodeling processes.
In our study, blood exchange lasted 3 months, which allowed blood cells to effectively repopulate tissues between partners of different ages. However, this approach did not lead to an increase in the life expectancy of old partners.
How effective can the exchange and mutual influence of biologically active molecules between partners be? It is well known that the total blood volume is exchanged slowly in relation to the half-life of a number of biologically active substances—approximately from 0.17 (based on blood leukocyte exchange) to 20 times a day (based on Evans blue dye exchange) between parabiotic partners. 25,29 Therefore, factors that are rapidly cleared from the circulation do not reach equilibrium in the parabiotic union.
In particular, for the radiolabeled glucose analog, [ 18 F]-FDG, the equilibrium period in the brain did not occur up to 39 days of partners' coexistence. 26 Similarly, the blood glucose levels differed significantly between heterochronic parabionts up to 6 weeks of coexistence (our unpublished observations). At the same time, factors with a long half-life (including blood cells) can reach the same concentration and biological activity in both members of the parabiotic union.
We tested some clinical blood parameters and hormones in blood of parabionts with a short half-life (within 10–20 minutes), such as TG, glucose, insulin, IGF-1, testosterone, and corticosterone, and with a long half-life (>12 hours), such as cholesterol, HDL, and TT4. We found no significant age-related differences between parabiotic partners. In young isochronic parabionts, levels of corticosterone tended to increase compared with single mice.
In old isochronic mice, levels of corticosterone did not differ from those in old single animals. It has been shown that with aging in mice, the stress reactivity of the hypothalamic–pituitary–adrenal system decreases and behavioral manifestations of active resistance to a stressful situation become less pronounced. 30 These factors could lead to a relatively rapid recovery of hormonal parameters in both old parabiotic partners and young heterochronic mice.
The formation of parabiotic pairs itself significantly affected the animals, as is evident from the levels of glucose, TG, TT4, and IGF-1 in the blood. We can explain this by two possible hypotheses.
First, the decreased level of blood TT4, as well as the tendency of decrease in IGF-1 and thymus mass, observed in young isochronous partners could be caused by stress, 31,32 especially during the 1st month of parabiosis.
Stress has numerous effects on physiology. It can decrease the level of TT4 and suppress the immune system, 33 which can adversely affect the viability of an individual. In this regard, it should also be noted that mortality increased among young isochronic partners in the first few months after the pair separation, which is evidenced by the lowest value of the 25th percentile of life expectancy among all the groups of parabionts.
Second, we suppose that such effects of parabiosis, especially in young partners, may be mediated by factors that control body and organ size. 24 It is well known that animals progressively lose weight after parabiotic surgery. In our studies, weight loss was ∼26.5% in young isochronic partners at 3 weeks postoperatively compared with sham-operated animals. Later on, weight loss was only 13.3% at week 6 of parabiosis in animals of this group and remained at this level until 12 weeks after surgery. The same pattern was observed for the weight of the liver and spleen (unpublished data).
It is well known that the main mechanisms that control body and organ size are mediated by insulin or insulin-like growth factors in mammals. 34 Therefore, a sharp decrease in IGF-1 concentration in the blood of young isochronic parabionts can be explained by a compensatory mechanism aimed at regulating the size of body/tissues of the “common” organism.
The size of internal organs can be regulated by feedback factors such as transforming growth factor beta (TGFβ), growth differentiation factor 11 (GDF11), and mammalian target of rapamycin (mTOR). 24 Some of these factors are also involved in age-related disorders. In particular, this mechanism may be potentially responsible for the decrease in cardiomyocyte hypertrophy seen during heterochronic parabiosis. 6 Changes in levels of the studied hormones were not found in the serum of old isochronic parabionts when compared with single animals, which may suggest a decrease in the activity of this mechanism with age.
On the other hand, we did not find any difference in the studied blood parameters between the groups of young isochronic and heterochronic parabionts. Therefore, this mechanism and blood hormone level cannot be involved in such a drastic reduction in the life span of young mice coexisting with old partners.
Stress can not only negatively affect life expectancy but also be beneficial due to the effect of hormesis. 35 In our experiment, the manifestation of hormesis could be an increase, compared with single sham-operated mice, in the median and 75th percentile of the life span in groups of young isochronic (+55% and +37%, respectively) as well as old iso- (+51% and 31%, respectively) and heterochronic (+63% and +72%, respectively) parabionts.
The effect of hormesis is weaker in old animals, 35 which is consistent with a less pronounced increase in these parameters in the group of old isochronic mice compared with young ones. It is noteworthy that old heterochronic mice showed a slight increase in the median and 75th percentile of the life span, and the maximum life span was higher than in the group of isochronic animals, which can be considered suggestive of “rejuvenation,” although statistically insignificant. In this context, the increase in testis mass in old mice joined with a young partner should also be noted.
The most robust and interesting result of this study is the significant decrease in the life span of young mice from heterochronic parabiotic pairs. Due to the complexity of the studied hormonal and metabolic parameters, the mechanisms of this phenomenon were not sufficiently elucidated, and the solution of this problem requires further research.
In addition, we should mention our previously published data on accelerated aging of the immune system of a young parabiont under the influence of an old partner. One of the possible mechanisms may be the accelerated aging of the immune system of young heterochronic parabionts, which we observed in previous studies. 16,17 In addition, aging of the immune system may be one of the factors causing a decrease in the life expectancy of young animals in heterogeneous pairs.
Conclusions
Mutual blood exchange by parabiosis between animals of different ages for 3 months did not lead to an increase in life expectancy of old partners as after separation, their average and maximum life expectancy did not differ from the animals of the isochronic group.
There may be several assumptions about the reason for such immunity of the old organism to the rejuvenating effects of young blood: first, the inability of circulating stem cells to occupy niches of resident tissue stem cells in a parabiosis partner; and second, the inability to reach equilibrium concentrations for the many biologically active blood factors, which have a short half-life, as a result of a long period of blood exchange between partners.
In addition, even such a factor as double dilution of old blood with young blood did not lead to an increase in life expectancy in old heterochronic parabionts.
Is it possible to prolong old life by transfusion of young blood in principle? Perhaps the reason for the absence of the rejuvenation effect is the elusive “aging factors,” which easily overcome all the barriers listed above, migrate into the young organism, and intensively “age” it, leading to a sharp reduction in the life expectancy of young partners afterward.
This assumption has already been confirmed in other studies 14,36 and we hope this area will be successfully developed in the future.
Footnotes
Author Disclosure Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as potential conflicts of interest.
Funding Information
The study was supported by a grant from the Academy of Medical Sciences of Ukraine, 0109U001723.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.