
Abstract
Oncogene-induced senescence (OIS), characterized by irreversible cell cycle arrest by oncogene activation, plays an important role in the pathogenesis of aging and age-related diseases. Recent research indicates that OIS is driven by activation of mitogen-activated protein kinase (MAPK). However, it is not apparent whether MAPK inhibition helps to recover senescence. In our previous study, we uncovered p38 MAPK inhibitor, SB203580, as an effective agent to reduce reactive oxygen species and increase proliferation in premature senescent cells. In this study, we evaluated whether SB203580 could ameliorate senescence in normal senescent cells. The senescence-improving effect was observed in the results that SB203580 treatment restored lysosomal function, as evidenced by a decrease in lysosomal mass and an increase in autophagic vacuoles. Then, SB203580-mediated lysosomal function restoration triggered the clearance of damaged mitochondria, leading to metabolic reprogramming necessary for amelioration of senescence. Indeed, p38 MAPK inhibition by SB203580 improved key senescent phenotypes. Our findings suggest a novel mechanism by which modulation of p38 MAPK activity leads to senescence improvement through functional restoration of lysosome and mitochondria.
Introduction
Somatic cells can divide only for a limited time and become senescent as the number of divisions increases, losing their reproductive capacity. 1 Thus, replicative senescence is a type of irreversible growth halt linked to decreased replication capacity. 2 Oncogene-induced senescence (OIS) is recently discovered to cause replication-independent senescence. Specifically, oncogene activation induces hyperproliferation followed by replicative senescence. 3
In human fibroblasts, OIS was primarily observed after oncogenic Rat sarcoma virus (RAS) activation. 4 In particular, oncogenic RAS initiates the RAS-rapidly accelerated fibrosarcoma (RAF)-MAPK/ERK kinase (MEK)-extracellular signal-regulated kinase (ERK) signaling cascade, activating p53, resulting in cell cycle arrest. 5 Numerous investigations have demonstrated that the RAS-induced OIS is driven by the mitogen-activated protein kinase (MAPK) pathway, which functions downstream of the RAS-RAF-MEK-ERK cascade. 6,7 Our prior research showed that BRAF inhibition successfully reduced senescence by inhibiting the RAS-RAF-MEK-ERK signaling cascade. 8 Given that the MAPK is the pathway that is downstream of the RAS-RAF-MEK-ERK signaling system, it is also important to examine if inhibiting the MAPK pathway appropriately will recover senescence.
MAPKs are serine and threonine protein kinases that play key roles in several signaling pathways, including those that control cell differentiation, division, and proliferation. 9 One of the main MAPK pathways is p38 MAPK, which was first revealed as a modulator of inflammatory and stress responses. 10 p38 MAPK has four isoforms consisting of p38α, p38β, p38γ, and p38δ, each encoded by a separate gene. These isoforms are distinct from one another regarding tissue distribution and regulation by upstream stimuli.
Cellular organelles that undergo the most obvious alterations during senescence are the lysosomes and mitochondria. 11 While mitochondria gradually change in both structure and function, lysosomes progressively alter the structure and function. 12 A buildup of dysfunctional mitochondria results in an overabundance of reactive oxygen species (ROS) production. The electron transport complex (ETC) in mitochondria is harmed by ROS as a result. 13 The ETC dysfunction lowers oxidative phosphorylation (OXPHOS), which lowers the effectiveness of adenosine triphosphate (ATP) synthesis. Senescent cells rely more on glycolysis than OXPHOS as a source of ATP energy to replenish ATP deficiency. 8,14 ROS make it more difficult for defective mitochondria to be cleared. 15 Lysosomes are also targeted by excess ROS, which compromises their functionality. 16,17 This harmful feedback loop between mitochondria and lysosomes accelerates and exacerbates senescence. These findings suggest that a ground-breaking therapy to recover senescence may involve the correct regulation of the feedback loop between them.
In a recent study, we discovered SB203580, a p38 MAPK inhibitor, which causes premature senescent cells to proliferate more and produce less ROS. 18 However, it is not yet known whether this drug promotes senescence amelioration even in normal senescent cells. In this study, we demonstrated that p38 MAPK inhibition by SB203580 can control the senescence process by regulating lysosomal and mitochondrial function.
Materials and Methods
Statement of informed consent
There are no human subject in this article and informed consent is not applicable.
Cell culture
The American Type Culture Collection (Manassas, VA, USA) provided the human diploid fibroblasts (PCS-201-010) for this work. Cells were grown as previously mentioned. 8 Cells were serially passaged at a 1:4 dilution during early passages and a 1:2 dilution during late passages. When the population doubling time of the cells was more than 14 days and less than 2 days, the cells were considered senescent and young, respectively. The final concentration of SB203580 (S1076; Selleck Chem, Houston, TX, USA) and DMSO (D8148; Sigma, St. Louis, MO, USA) in the culture medium was 0.5 μM and 1.4 mM, respectively. Cells were exposed to DMSO or SB203580 for 21 days before the functional test.
Cell proliferation assay
Cell proliferation assay was carried out in 96-well plates (353072; Corning, Glendale, AZ, USA) at a cell density of 2000 cells per well. To precisely count the number of cells, a cell proliferation assay based on a DNA content-based approach was used. 19
Western blot analysis
Western blot analysis was performed as described previously. 20 The primary antibodies used in this study included the p38 MAPK antibody (9212; 1:500 dilution; Cell Signaling Technology, Danvers, MA, USA), AKT antibody (9272; 1:500 dilution; Cell Signaling Technology), phospo-AKT antibody (4058T; 1:500 dilution; Cell Signaling Technology), and HRP-conjugated β-actin (sc47778; 1:1000 dilution; Santa Cruz). The secondary antibodies included the HRP-conjugated anti-rabbit antibody (sc-2357; 1:1000 dilution; Santa Cruz).
Measurement of ROS and mitochondrial mass
To determine the level of ROS, cells were incubated at 37°C for 30 minutes with media containing 5 μM MitoSOX (M36008; Life Technologies, Carlsbad, CA, USA). Then, cells were processed for FACS analysis as previously explained. 21 To determine the level of mitochondrial mass, cells were incubated at 37°C for 30 minutes with media containing 50 nM MitoTracker Deep Red (M22426; Life Technologies).
Flow cytometric analysis of cellular lipofuscin levels
Most unsaturated fatty acids are attached to proteins, where they are then peroxidized to create lipofuscin. Lipofuscin exhibits autofluorescence due to the Schiff base created by the interaction between carbonyl and amino molecules. 22 The widely used technique for lipofuscin quantification is autofluorescence assessment. 23 –26 A 530/30 nm bandpass filter with excitation at 488 nm was applied in flow cytometry to measure autofluorescence. The outcomes were examined using the program Cell Quest 3.2 (Becton Dickinson).
Measurement of lysosomal mass and autophagic vacuoles
Lysosomal mass was measured as described previously. 27 The level of autophagic vacuoles was assessed using Cyto-ID as previously mentioned. 28
Immunofluorescence
On chamber slides (30104; SPL, Pocheon, Korea), cells were plated for immunofluorescence. Cells were permeabilized for 15 minutes with 0.1% Triton X-100 in phosphate-buffered saline (PBS) after being fixed for 15 minutes with 4% paraformaldehyde in PBS. A blocking procedure using 10% fetal bovine serum in PBS was carried out at room temperature for 1 hour. The rabbit anti-LC3B antibody (2775; 1:200 dilution; Cell Signaling Technology) and mouse anti-total OXPHOS antibody (AB110411; 1:1000 dilution; Abcam, Cambridge, United Kingdom) were used to incubate cells at 4°C overnight. Subsequent procedures in immunofluorescence were performed as described previously. 8
Analysis of the oxygen consumption rate (extracellular acidification rate)
The XFe24 flux analyzer (Seahorse Bioscience XFe24 Instrument; Seahorse Bioscience, Billerica, MA, USA) was used. Briefly, 5 × 104 cells were placed in each well of an XFe24 cell-culture plate (100850-001; Seahorse Bioscience) and examined as previously described. 8
Senescent-associated β-galactosidase staining
Senescent-associated β-galactosidase (SA-β-gal) staining was performed according to the manufacturer's protocols (9860; Cell Signaling Technology, Beverly, MA, USA).
Statistical analysis
Statistical analysis was carried out using SigmaPlot 12.5 (Systat Software, San Jose, CA, USA). The Student's t-test and two-way analysis of variance (ANOVA) followed by Bonferroni's posttest were used to determine the significance of differences.
Results
Confirmation of senescence amelioration by p38 MAPK inhibition
SB203580, a p38 MAPK inhibitor, has been identified as a candidate reagent to increase cell proliferation and decrease ROS in premature senescent cells. 18 In this study, we investigated whether this reagent would be effective in ameliorating senescence in normal senescent cells.
Optimal levels of p38 MAPK inhibition were assessed by treating normal senescent cells with different concentrations (0–2 μM) of SB203580. The effect on cell proliferation was significantly higher at the 0.5 μM concentration (Supplementary Fig. S1). Based on this result, the 0.5 μM concentration was chosen for the subsequent tests.
The specificity of SB203580 as a p38 MAPK inhibitor was investigated before further experiments. SB203580 inhibits p38 MAPK catalytic activity and activation of protein kinase B (known as AKT). 29 Recent studies have shown that p38 MAPK plays an upstream signaling role regulating AKT at the protein and transcription level, suggesting a cross talk between them. 30 Therefore, to identify the specificity of SB203580 as a p38 MAPK inhibitor, we evaluated the expression levels of p38 MAPK, AKT, and phospho-AKT. SB203580 did not reduce the expression levels of p38 MAPK and AKT, whereas SB203580 reduced phospho-AKT levels (Supplementary Fig. S2). These data suggest that SB203580 specifically inhibited p38 MAPK catalytic activity, resulting in reduced phosphorylation levels of AKT, which function as a downstream signal pathway of p38 MAPK, without inhibiting the expression of p38 MAPK and AKT.
To confirm the SB203580-mediated proliferation-inducing effect observed in premature senescent cells, cell proliferation was evaluated at various time points (0–16 days). Cell proliferation was significantly increased upon SB203580 treatment, suggesting that SB203580 abolished cell cycle arrest in normal senescent cells (Fig. 1A).
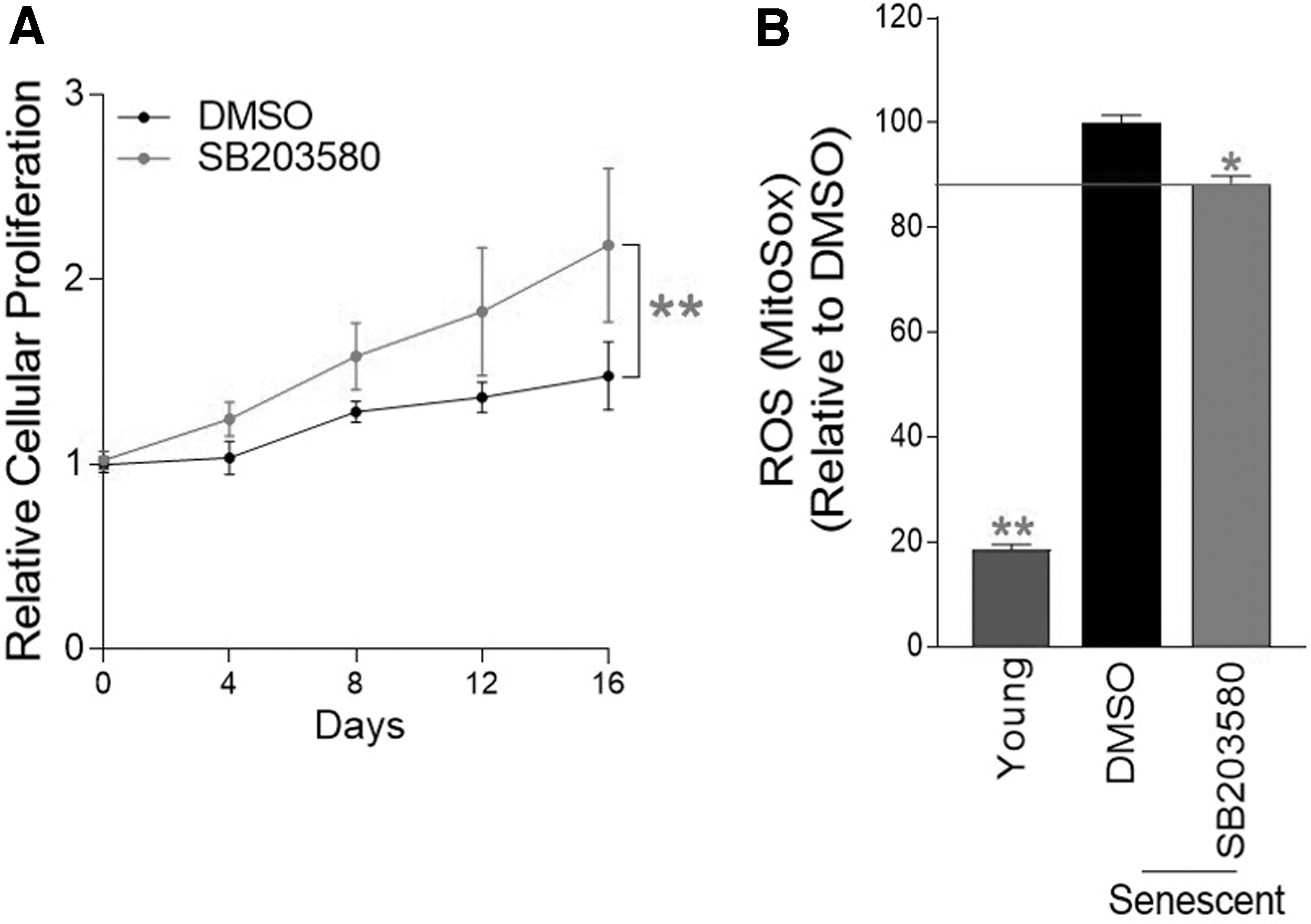
Confirmation of senescence amelioration by p38 MAPK inhibition.
We then investigated whether SB203580 would reduce ROS levels in normal senescent cells. To detect ROS, especially superoxide anions, cells were stained with a medium containing 5 μM MitoSOX. Senescent cells had higher ROS levels than young cells, confirming the senescence-associated ROS increase (Fig. 1B). 31 Treatment with SB203580 significantly decreased ROS levels in normal senescent cells (Fig. 1B).
p38 MAPK inhibition leads to the restoration of lysosomal function
Since we confirmed the increase in cell proliferation and the reduction in ROS in normal senescent cells, we investigated how SB203580 affects different senescent phenotypes. The accumulation of lipofuscin, one of the prominent senescent phenotypes, was examined. 32 The level of lipofuscin was determined by measuring the amount of intracellular autofluorescence. 33 After treatment with SB203580, autofluorescence levels significantly decreased, suggesting that SB203580 inhibited the buildup of lipofuscin in senescent cells (Fig. 2A).
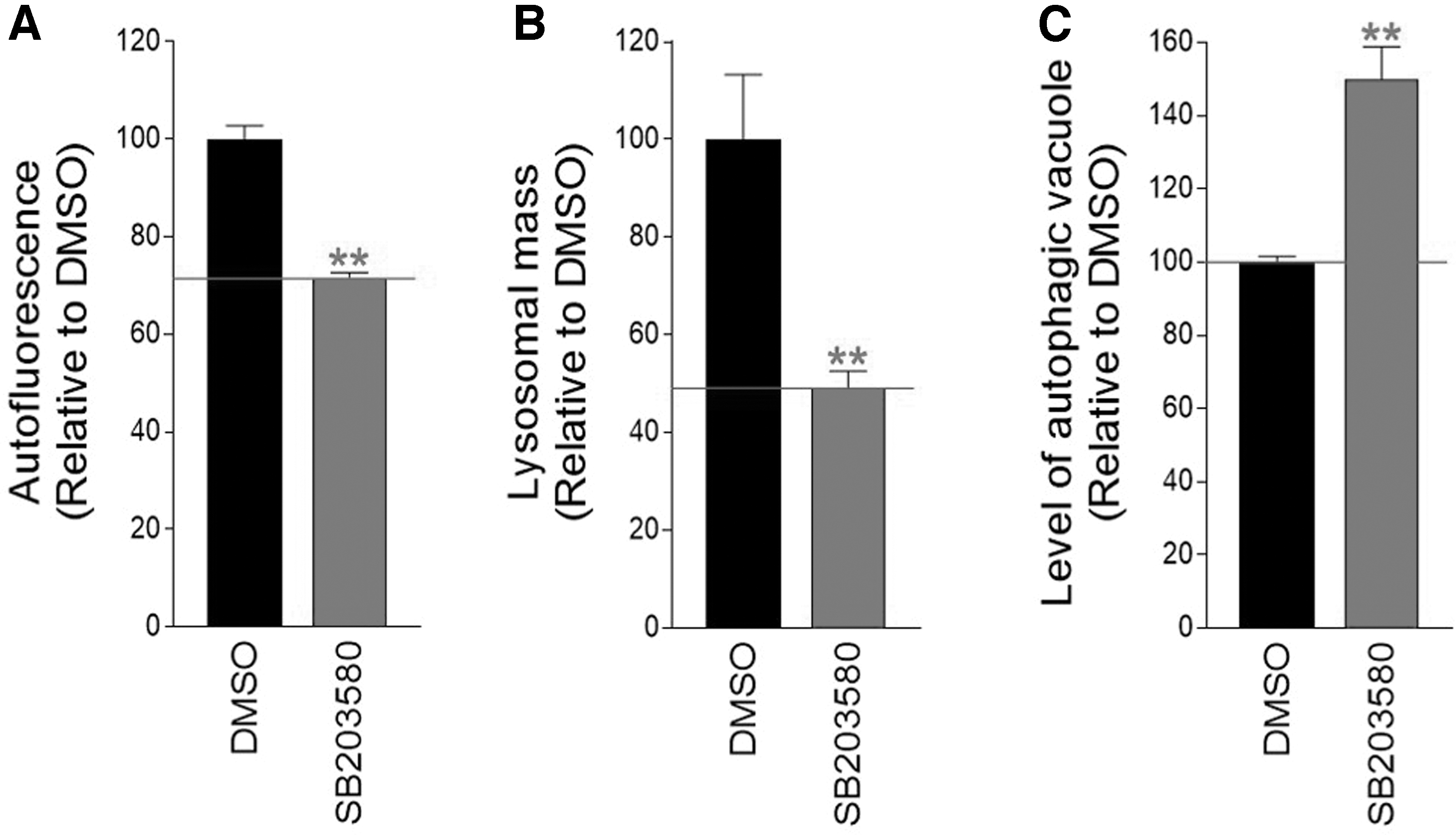
p38 MAPK inhibition leads to the restoration of lysosomal function.
An autofluorescent material called lipofuscin slowly accumulates in lysosomes. Lysosomes filled with lipofuscin act as sinks for newly formed hydrolases, increasing the lysosomal mass as nonfunctional lysosomes increase. 34 So, we checked to see whether SB203580 had any impact on the lysosomal mass. Indeed, SB203580 significantly decreased lysosomal mass, indicating a reduction in nonfunctional lysosomes (Fig. 2B).
Restoration of lysosomal function facilitates autophagic activity and initiates autophagy-dependent substrate clearance. 35 –37 Because we found a reduction in nonfunctional lysosomes, we further examined how SB203580 affects autophagic activity, measurable in the amount of autophagic vacuoles. 38 The level of autophagic vacuoles increased significantly after SB203580 treatment, demonstrating that SB203580 restored autophagy activity (Fig. 2C).
Activation of mitophagy through p38 MAPK inhibition to eliminate dysfunctional mitochondria
A functional lysosomal/autophagy system is essential to clear dysfunctional or damaged mitochondria. 39 We therefore investigated whether the clearance of malfunctioning mitochondria is triggered by repair of the SB203580-mediated lysosomal/autophagy system. Mitochondrial morphology was viewed using antibodies to total OXPHOS proteins, and autophagy was viewed using antibodies to the autophagic protein, microtubule-associated protein 1 LC3B.
Compared with young cells, senescent cells showed a more branching mitochondrial morphology (Fig. 3A). LC3B and mitochondria colocalized in young cells, but this colocalization was infrequent in DMSO-treated senescent cells (Fig. 3A). However, SB203580 treatment caused colocalization of LC3B and mitochondria to reappear (Fig. 3A; white arrows). Senescent cells had a higher level of mitochondrial mass than young cells, supporting the senescence-associated increase in mitochondrial mass (Fig. 3B). 40 The simultaneous reduction in mitochondrial mass was shown in SB203580-treated senescent cells, suggesting that mitophagy, an autophagic mechanism that selectively degrades defective mitochondria, was triggered (Fig. 3B).
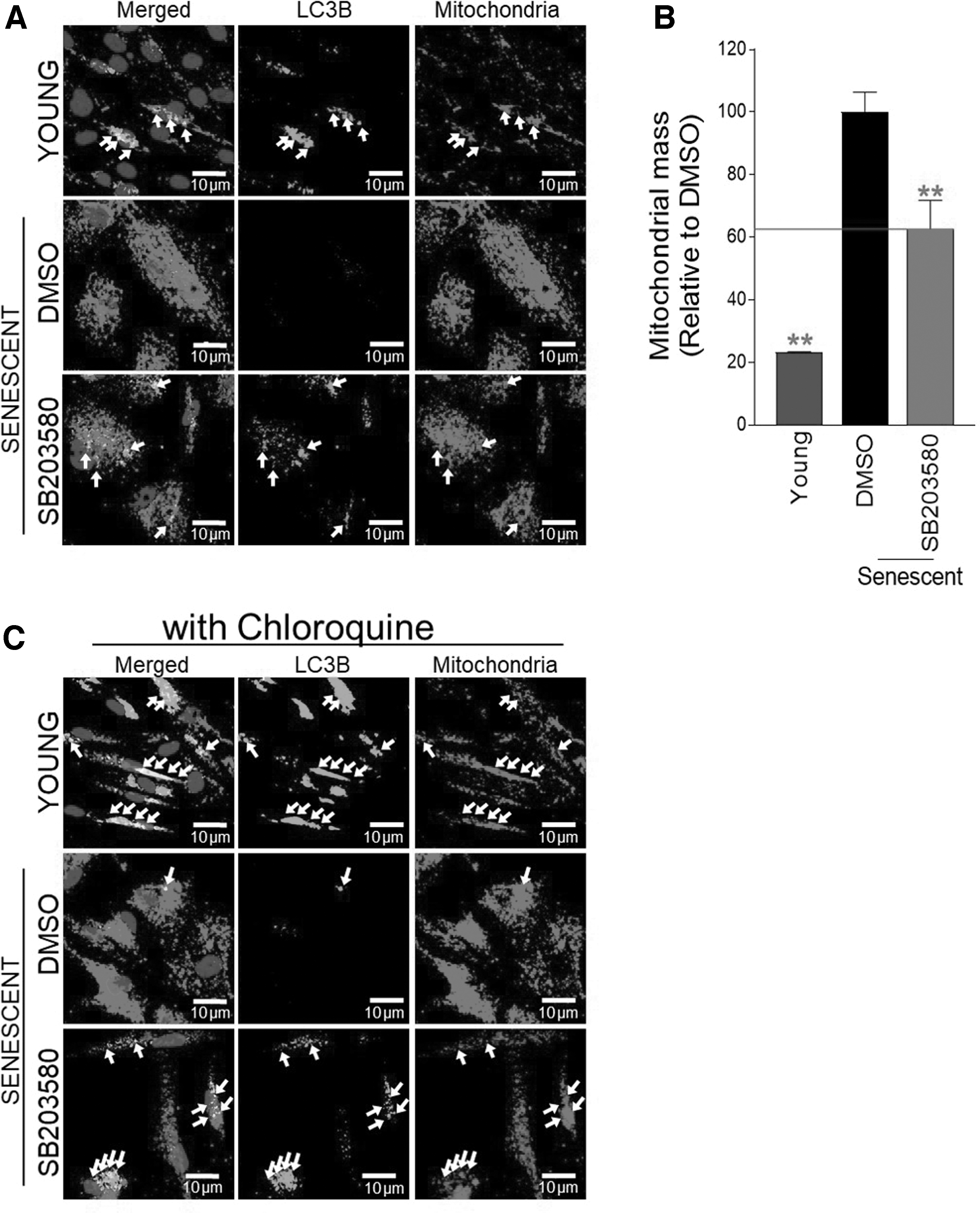
Activation of mitophagy through p38 MAPK inhibition to eliminate dysfunctional mitochondria.
To determine if the reconstituted lysosomal/autophagy system activates mitophagy, cells were cotreated with chloroquine, a drug that inhibits autophagy flux. 41 Young cells and SB203580-treated senescent cells increased colocalization, whereas DMSO-treated senescent cells did not (Fig. 3C; white arrow). These findings suggest that restoration of the lysosomal/autophagy system through SB203580 activated mitophagy.
Inhibition of p38 MAPK restores metabolism by reducing glycolysis dependence
The preservation of mitochondrial function is achieved by the efficient elimination of damaged mitochondria through mitophagy.
42
Consequently, the elimination of damaged mitochondria reduces ROS generation.
13
As we observed the appearance of mitophagy and reduction of ROS by SB203580, we evaluated whether mitochondrial metabolism was restored by SB203580. To indirectly monitor changes in mitochondrial metabolism, we measured the level of glycolysis, which can be determined by monitoring the extracellular acidification rate (ECAR). The substances (glucose, oligomycin, and 2-deoxy-
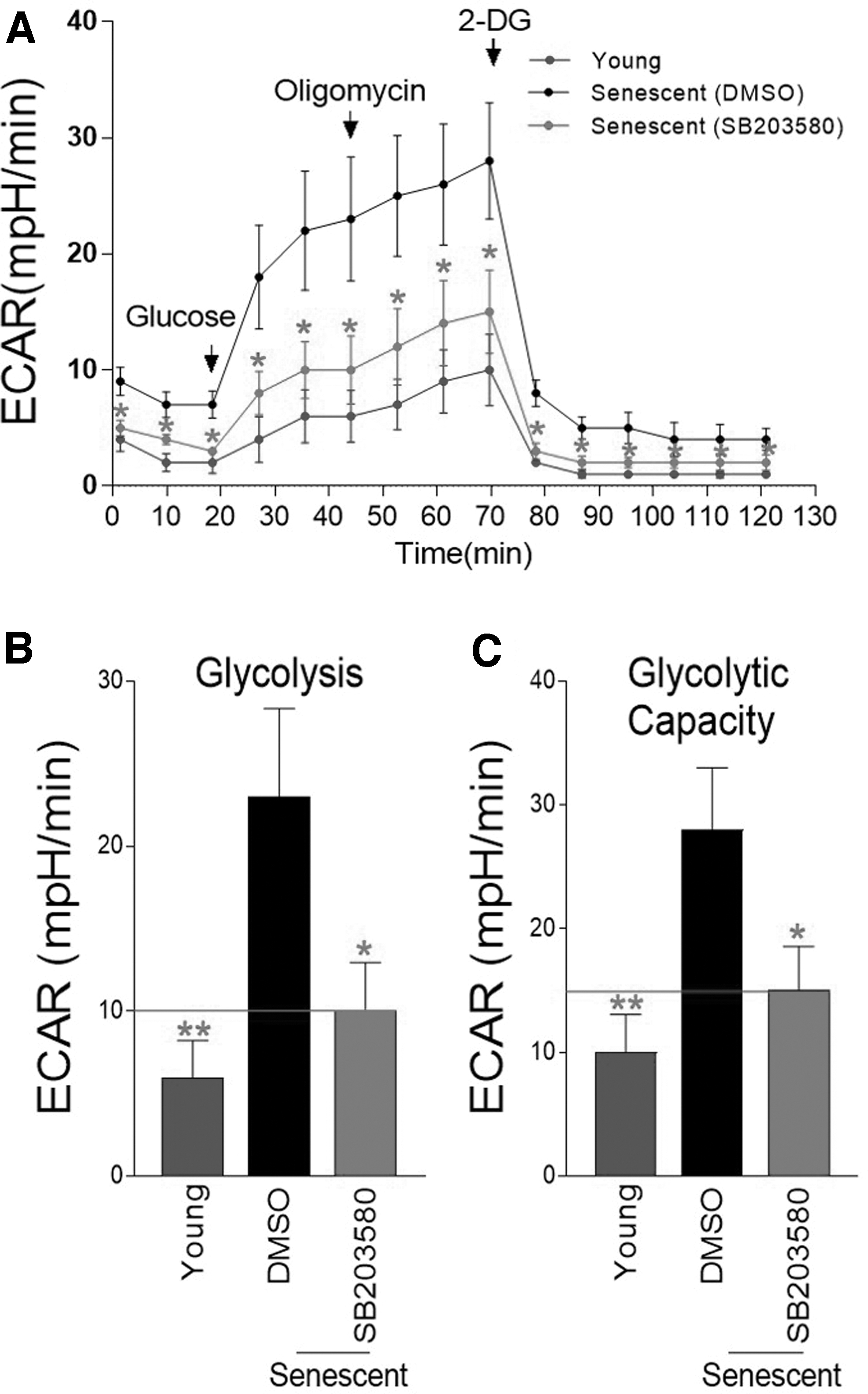
Inhibition of p38 MAPK restores metabolism by reducing glycolysis dependence.
However, after SB203580 treatment, ECAR values were significantly decreased compared with a DMSO control, indicating a decrease in glycolysis dependence, a common phenomenon of mitochondrial metabolic recovery (Fig. 4A–C).
p38 MAPK inhibition ameliorates senescence phenotypes
Metabolic reprogramming was proven to be necessary for senescence amelioration. 8 As we identified metabolic reprogramming with a reduced dependence on glycolysis by SB203580, we examined whether SB203580 treatment would ameliorate senescent phenotypes. Since senescence is usually diagnosed using a surrogate measure of SA-gal activity, 43 the proportion of SA-gal-positive cells was examined. DMSO-treated senescent cells had a higher proportion of SA-β-gal-positive cells than young cells, whereas SB203580 treatment significantly reduced the proportion in senescent cells (Fig. 5A).
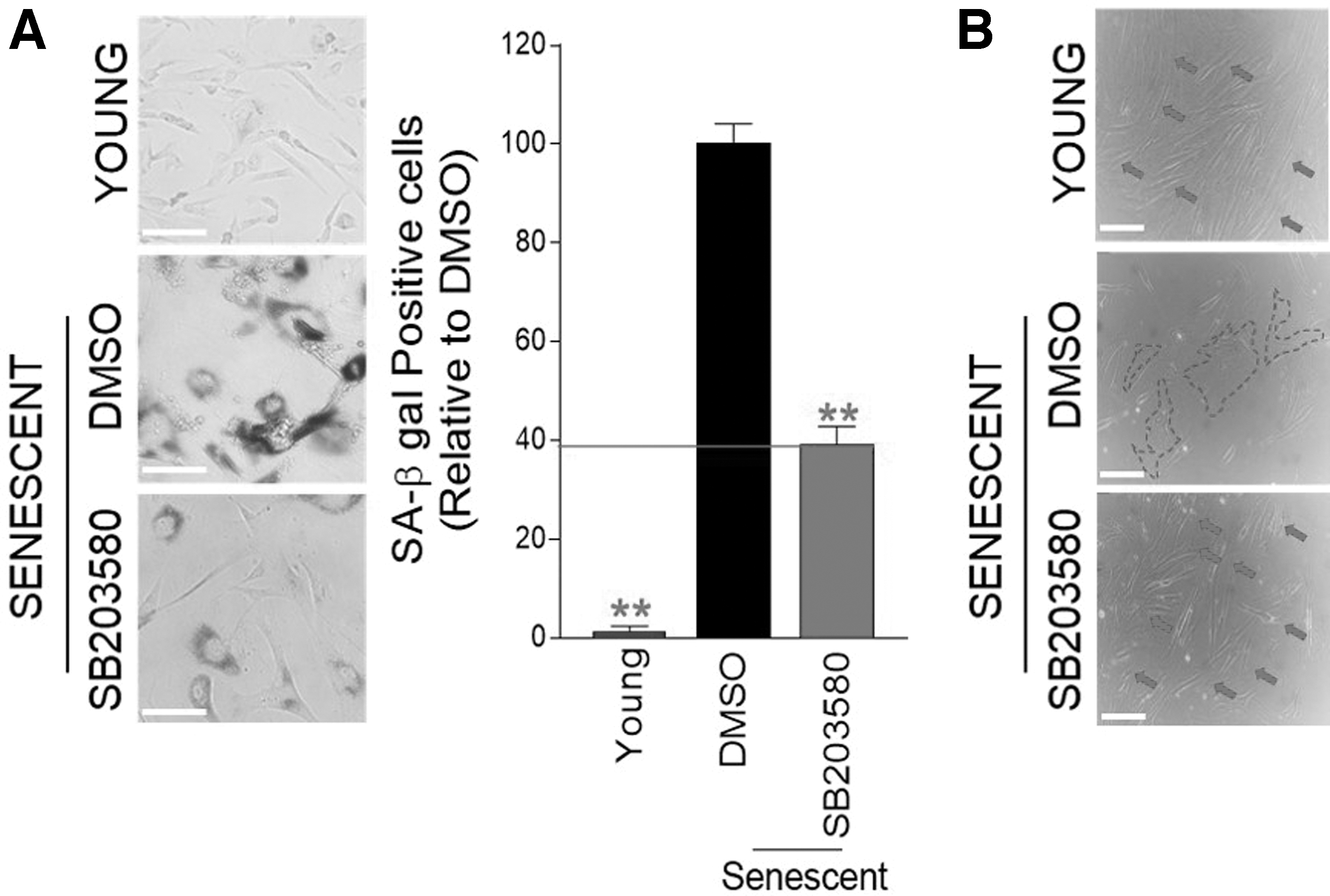
p38 MAPK inhibition ameliorates senescence phenotypes.
Finally, since the increase in cell surface area is one of the most noticeable senescent phenotypes, 44 the morphological changes following SB203580 treatment were examined. Young cells showed the tiny spindle-shaped structure (red arrows), whereas DMSO-treated senescent cells showed broad and flat structures (dotted line) (Fig. 5B). Treatment with SB203580 markedly recovered the morphology of senescent cells to a tiny spindle-shaped structure (red arrows) (Fig. 5B).
Discussion
Lysosomal dysfunction reduces mitochondrial turnover, increasing the production of ROS, which attack the lysosome. 34 Lysosomes and mitochondria are thus trapped in a vicious feedback loop that exacerbates senescence phenotypes. The extent of senescence has been regulated by fine-tuning the interaction between lysosomes and mitochondria. The importance of this cross talk on the regulation of senescence is supported by the discovery that lysosomal reacidification through the attenuation of ataxia telangiectasia mutant activity accelerated the clearance of defective mitochondria, which in turn reversed the senescent phenotypes. 14 Furthermore, these findings are highlighted by other studies that the trehalose, a mitophagy inducer, improved mitochondrial quality and reduced age-related atherosclerosis in mice. 45
Therefore, learning how to regulate the interactions between lysosomes and mitochondria could be the key to establishing therapeutics for age-related diseases and aging itself. In this study, we demonstrated that modulation of cross talk by p38 MAPK inhibition mediates a unique strategy to alleviate senescence. Notably, p38 MAPK inhibition promoted the restoration of lysosomal function, triggering the clearance of defective mitochondria. Efficient removal of damaged mitochondria restored mitochondrial function by reducing dependence on glycolysis, a phenomenon known to be necessary for improving senescence. 8 Our study provides the first evidence that p38 MAPK inhibition triggers functional recovery of lysosomes and mitochondria, demonstrating that modulating cross talk between lysosomes and mitochondria by controlling p38 MAPK activity is an effective therapeutic strategy for senescence.
Mitochondria function as intracellular ATP energy producers. They are now understood to be a vast signaling platform that regulates a number of significant cellular and physiological activities. 46 Recently, there is increasing evidence that mitochondria are not only an energy-producing unit, but also support a more important function in the regulation of senescence. 14,18,47,48 In particular, reducing the activity of certain protein kinases restored mitochondrial function to the status of young cells and postponed the progression of senescence. For example, suppression of the rho-associated protein kinase (ROCK) lowered ROS generation while also restoring mitochondrial function. 18,47 This inhibition also changed the energy source's dependence from glycolysis to OXPHOS. In addition, BRAF inhibition caused metabolic reprogramming and mitochondrial functional recovery, which are two crucial aspects that are disrupted in senescent cells. 8 Metabolic reprogramming through BRAF inhibition served as a prerequisite for senescence improvement.
Consistent with these findings, we found that inhibition of p38 MAPK induced metabolic reprogramming accompanied by improved senescence. In particular, p38 MAPK inhibition blocked the malignant feedback loop between lysosomes and mitochondria, reducing mitochondrial damage while reducing reliance on glycolysis. Given that modulating protein kinase activity can improve aging while achieving metabolic reprogramming, controlling the activity of protein kinases such as ROCK, BRAF, and p38 MAPK in various combinations may be a more efficient approach.
MAPK controls cell survival, proliferation, and differentiation through downstream signaling molecules, and cellular senescence is also one of the factors controlled by MAPK. 49 In the presence of senescence-inducing stimuli, MAPKs act as sensors that determine whether to induce cells to apoptosis, senescence, or other responses. When cells are determined to adopt a senescence response, MAPK participates in the implementation of gene expression programs that allow indefinite growth arrest, including increased synthesis of p53, p21, and p16. 49 Furthermore, MAPKs transcriptionally regulate secretion of senescence-associated secretory phenotype through nuclear factor-κB pathways. 50 Given that MAPK signaling is highly connected with senescence, the regulation of MAPK activity might uncover new tools for therapeutic benefit. In this study, a beneficial effect on senescence was found using SB203580, an ATP-competitive pan-p38 MAPK inhibitor. 51 However, we acknowledge that we are unable to identify the specific p38 MAPK subtype that controls senescence.
Therefore, to further investigate the underlying mechanism of SB203580-mediated senescence amelioration, it should be warranted to identify p38 MAPK subtypes that permit effective senescence improvement.
Conclusion
In conclusion, we presented a novel mechanism for how p38 MAPK inhibition by SB203580 controlled senescence. In particular, p38 MAPK inhibition had a restorative effect by recovering the lysosomal and mitochondrial functions, two critical aspects that were compromised in senescent cells. All things considered, our findings suggest that effective modulation of p38 MAPK activity might be a therapeutic strategy for senescence.
Footnotes
Authors' Contributions
H.W.K., Y.B., and J.T.P. conceived of and designed the experiments. J.Y.P., H.L., E.S.S., Y.H.L., M.U.K., and G.K. performed the experiments. J.Y.P., H.W.K., Y.B., and J.T.P. analyzed the data. J.Y.P., H.W.K., Y.B., and J.T.P. wrote and edited the article.
Author Disclosure Statement
No funding was received for this article.
Funding Information
This research was supported by the Technology Innovation Program (Alchemist Project, 20019181) through the Korea Evaluation Institute of Industrial Technology (KEIT) funded by the Ministry of Trade, Industry and Energy. This research was also supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (NRF-2021R1A2C1004298) and by the Ministry of Education (NRF-2019R1A6A103031807).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.