
Abstract
We previously demonstrated that a 50% caloric restriction (CR) from birth improves several cardiometabolic risk factors in young rats. In this study, we investigated in middle-aged rats the consequences of a 50% CR from birth on cardiometabolic risk factors, heart function/morphology, ventricular arrhythmia, and fibrillation incidence, and cardiac intracellular proteins involved with redox status and cell survival. From birth to the age of 18 months, rats were divided into an Ad Libitum (AL18) group, which had free access to food, and a CR18 group, which had food limited to 50% of that consumed by the AL18. Resting metabolic rate, blood pressure, and heart rate were recorded, and oral glucose and intraperitoneal insulin tolerance tests were performed. Blood was collected for biochemical analyses, and visceral fat and liver were harvested and weighed. Hearts were harvested for cardiac function, histological, redox status, and western blot analyses. The 50% CR from birth potentially reduced several cardiometabolic risk factors in 18-month-old rats. Moreover, compared with AL18, the CR18 group showed a ∼50% increase in cardiac contractility and relaxation, nearly three to five times less incidence of ventricular arrhythmia and fibrillation, ∼18% lower cardiomyocyte diameter, and ∼60% lower cardiac fibrosis. CR18 hearts also improved biomarkers of antioxidant defense and cell survival. Collectively, these results reveal several metabolic and cardiac antiaging effects of a 50% CR from birth in middle-aged rats.
Introduction
Billions of dollars are spent every year to treat cardiometabolic diseases, the leading cause of morbidity and mortality worldwide. 1,2 The earlier the preventive measures are taken, the greater the delay in cardiometabolic disease onset. 3,4 Excessive calorie consumption is directly associated with a host of cardiometabolic diseases and aging. 5,6 On the contrary, calorie restriction (CR) is the most important nonpharmacological intervention to improve health and extend lifespan. 7 CR counteracts the cardiovascular aging process indirectly by reducing cardiovascular risk factors, such as elevated blood pressure, dyslipidemias, and hyperglycemia, 1 and directly by reducing the atherosclerotic process, improving cardiac function, reducing cardiac arrhythmia/fibrillation incidence, and inhibiting cardiomyocyte hypertrophy. 8 –10 Moreover, CR has been linked to enhancing several cell survival genes, making CR protocols a potential approach to elucidate heart aging mechanisms and develop CR mimetic agents, such as resveratrol. 8
Preclinical studies with calorie-restricted aged animals have often been used to examine the effects of CR on lifespan and diseases. In elderly rhesus monkeys, a 30% CR for 6 months delayed the onset of the aging process and age-associated diseases. 11 In aged rodents, 40% CR for 3 months exerted several antiaging effects on the heart, such as improvement of cardiac function, reduction in cardiac fibrosis, and enhancement of intracellular antiapoptotic signaling pathway. 12 Over the years, the decrease in resting metabolic rate and the consequent decrease in reactive oxygen species generation and cell damage has been strongly associated with CR's beneficial effects on health and longevity in both preclinical 13 and clinical studies 14 However, the long-term effects of CR on cardiometabolic health are not well understood, particularly if CR is initiated early in life.
We recently found a consistent reduction in multiple cardiometabolic risk factors in young rats' 50% calorie-restricted since birth, such as improvement in insulin tolerance and serum lipid profile and a robust reduction in visceral fat content and blood pressure. 15,16 Moreover, these young rats present improved cardiac function in basal conditions 16 and after ischemia/reperfusion insult. 17 Most of these cardiometabolic benefits were also found in young ovariectomized rats. 18 These previous studies suggest that an intense CR from birth may have long-term cardiometabolic beneficial effects. A key question is whether the beneficial cardiometabolic effects of an intense CR from birth seen in young rats extend to middle-aged rats. In addition, it is unclear if an intense CR from birth ameliorates typical adverse responses of the heart caused by the aging process, such as the increase in the prevalence of ventricular arrhythmia and fibrillation and the imbalance of redox state and cell survival signaling.
Since middle age is the period of life with the highest incidence of cardiac diseases, 19 we looked into the effects of a 50% CR from birth on the hearts of rats at 18 months of age, which is equivalent to middle age in humans (∼45 years). 20 In particular, we investigated in 18-month-old rats the effects of a 50% CR from birth on cardiometabolic risk factors, heart function/morphology, ventricular arrhythmia/fibrillation incidence, as well as cardiac biomarkers of redox status and cell survival.
Materials and Methods
Experimental protocols were approved by a local Ethics Committee on Animal Use (protocol 031/2016) and carried out in accordance with the Guide for the Care and Use of Laboratory Animals published by the U.S. National Institute of Health. 21
Animals and experimental design
The calorie restriction protocol is described in previous studies. 15 –18 In brief, pregnant female Wistar rats (n = 12), ∼90 days old, were housed in individual cages and maintained in a room with controlled temperature (22°C ± 2°C), in a 12 hours light/12 hours dark cycle, and with free access to tap water and commercial rat chow containing 22% protein, 55% carbohydrate, and 33% lipid (Nuvilab Nutrients LTDA, Colombo, PR, Brazil). Immediately after the birth of pups, half of the dams (n = 6) were assigned to the Ad Libitum (AL) group, while the remaining half (n = 6) were assigned to the CR group.
The litter consisted of eight newborns per dam. The AL control dams were fed AL, and the CR dams were fed daily with 50% of the quantity consumed by the AL dams. Both groups had free access to water. Excess pups were removed after weaning (21 days) to form the experimental groups of eight male rats fed AL for 18 months (AL18) and eight male rats fed 50% of the amount consumed by the AL rats for 18 months (CR18). The pups were weighed, housed in individual cages, and received the same treatment as their dams. Figure 1 provides a comprehensive visual representation of the experimental design for this study.

Experimental setup. Diagram illustration describe the experimental groups, study design, and parameters under investigation.
Food intake, weight gain, and feed efficiency ratio
From 21 days to 18 months of age, weight gain was assessed weekly, food intake of the AL group was assessed daily, and this value was used to compute the offer to the CR18 group. The feed efficiency (ratio of weight gain to overall food consumption) was assessed for both AL18 and CR18 groups.
In vivo analyses
Resting metabolic rate
In vivo analyses were performed 1 week before euthanasia. Resting metabolic rate was recorded according to a previous study. 17 In brief, after a 12-hour overnight fast, the resting metabolic rate was estimated using the average oxygen consumption (VO2) and carbon dioxide production (VCO2) for 60 minutes. A computer-monitored indirect calorimeter (Oxyleptro; Harvard Apparatus, Spain) was connected to a metabolic chamber (airflow = 1.0/min) that housed the rats. A respiratory-based software program was used to measure the calorimetric parameters (software Metaoxy; Harvard Apparatus).
Oral glucose tolerance test
Immediately following the assessment of the basal metabolic rate, glucose was delivered through gavage (2 g/kg body weight; 50% solution). Blood glucose levels were evaluated by clipping a small portion of the rat's tail before (0 minutes) and 30, 60, and 120 minutes after the glucose challenge. An ACCU-CHEK analyzer (Advantage Glucose Analyzer; Roche Diagnostics Corporation, IN) was used to determine blood glucose levels. 15
Intraperitoneal insulin tolerance test
Forty-eight hours following the oral glucose tolerance test (OGTT), the animals fasted for eight hours before receiving an intraperitoneal injection of insulin (1 I.U./kg body weight). The same procedure of OGTT was used to determine blood glucose levels.
Systolic blood pressure and heart rate
Noninvasive tail-cuff plethysmography (MLT1020PPG IR Plethysmograph; ADInstruments, New South Wales, Australia) was used to assess systolic blood pressure and heart rate 16 48 hours following the intraperitoneal insulin tolerance test (IpITT).
Ex vivo analyses
After an 8-hour fasting period, the rats were decapitated 15–20 minutes after receiving an intraperitoneal injection of 400 IU heparin 24 hours after the systolic blood pressure and heart rate measurements. Blood was then taken, and heart, liver, and visceral fat were harvested and weighed.
Blood analyses
Blood was centrifuged to obtain plasma, which was aliquoted and stored at −80°C until analysis. Total plasma cholesterol (CHOL), high-density lipoprotein cholesterol (HDL-C), triglyceride (TG), IL-6, IL-10, and TNF-α levels were measured using commercial kits (LabTest; Lagoa Santa, Minas Gerais, Brazil). Using the Friedewald equation, the levels of very-low-density lipoprotein cholesterol (VLDL-C) and low-density lipoprotein cholesterol (LDL-C) were calculated.
22
The following equations were used to calculate the adiposity index,
23
atherogenic index,
24
and cardiovascular risk index
25
:
Isolated heart function
The Langendorff technique was used to record cardiac function. 15 In brief, the thorax was opened, the heart was carefully harvested and immediately cannulated, and then retrogradely perfused with Krebs–Ringer solution at 37.1°C, constant pressure of 65 mmHg, and oxygenation of 5% CO2 and 95% O2. A water-filled rubber balloon attached to a pressure transducer (MLT0380; ADInstruments) was introduced into the left ventricular chamber via the left auricle to record pressure. Using LabChart 8 software, the rate of pressure development (dP/dt) was calculated by differentiating the pressure variation records.
After stabilizing the perfusion system for 20 minutes, the baseline cardiac function was assessed for a further 20 minutes. The occurrence of ventricular arrhythmia was defined by the recording of at least five ectopic beats (irregular beats), while the occurrence of ventricular fibrillation was characterized by the recording of at least five fast, premature, and low contractions (low-pressure development). The records were performed visually, counted manually, and expressed as the frequency of events during the final 20 minutes of the recording period, as previously described. 3
Histological analysis
For assessments of cardiomyocyte diameter and cardiac collagen content, hearts were fixed in 4% Bouin, embedded in paraffin, and subsequently sectioned to a thickness of 5 μm. To access myocyte cross-sectional area, hematoxylin and eosin-stained heart slices were examined under 40 × magnification. Only longitudinally cut myocytes with the nucleus in the center and apparent cellular boundaries were evaluated. The cross-sectional diameter of the myocytes was calculated by tracing 50 to 100 individual cardiomyocytes within the ventricular free wall over five or six sections per heart using ImageJ software (National Institutes of Health). To evaluate cardiac fibrosis, sections were stained with Masson's trichrome to determine interstitial and perivascular collagen content. The extension of collagen deposition was quantified by percent area analysis at 20 × magnification using the ImageJ software. Seven fields in three distinct sections were evaluated per heart.
Cardiac redox status analysis
Cardiac Redox Status trials were conducted in accordance with a prior study. 17 In brief, left ventricular ventricle samples were immersed in phosphate-buffered saline (PBS) with a pH of 7.2, rigorously washed to remove blood, and kept at −80°C until the time of analysis. The samples were then homogenized for 3 minutes at 0°C–4°C at the time of the experiment. The homogenate was then centrifuged for 10 minutes at 10,000 g, and the supernatant was used for the biochemical analyses. According to previous procedures, the assays to measure superoxide dismutase 26 and catalase 27 activities were carried out. We used the production of thiobarbituric acid-reactive substances during an acid-heating reaction as an indicator of lipid peroxidation. 28 The ferric-reducing antioxidant power assay was conducted in accordance with previous procedures. 29
Western blot analysis
Using the Bradford method, the total protein content of left ventricular ventricles was determined. 30 A 10% polyacrylamide gel was loaded with 50 μg of protein for electrophoresis. Following electrophoresis, proteins were transferred to a PVDF membrane, blocked with a 5% solution of nonfat milk, and washed in PBS containing 0.1% Tween 20. Membranes were incubated overnight at 4°C with the primary antibodies: SOD1 (superoxide dismutase) (1:4000 dilution; Enzo Life Sciences), SOD2 (1:4000 dilution; Enzo Life Sciences), Hsp72 (1:5000 dilution; Enzo Life Sciences), Phospho-AMPK (1:5000 dilution; Enzo Life Sciences), AMPK (1:3500 dilution; Enzo Life Sciences), Phospho-P38 (1:4000 dilution; Enzo Life Sciences), P38 (1:4000 dilution; Enzo Life Sciences), Phospho-ERK1/2 (1:4500 dilution; Enzo Life Sciences), ERK1/2 (1:4000 dilution; Enzo Life Sciences), caspase 3 (1:500 dilution; Enzo Life Sciences), and glyceraldehyde 3-phosphate dehydrogenase (GAPDH, 1:5000 dilution; Cell Signaling).
Thereafter, Cell Signaling's monoclonal anti-rabbit or anti-mouse secondary antibody coupled with peroxidase (1:7500 dilution) was used. Enhanced chemiluminescence (Amersham Biosciences) was used for immunodetection, and protein levels were represented as a ratio of optical densities.
Statistical analysis
Data are presented as mean ± standard deviation (SD). Data normality was verified by the Shapiro–Wilk test. A two-way ANOVA followed by the Bonferroni post hoc test was used for the analysis of body weight, food intake, and feed efficiency ratio, while the remaining results were analyzed using the Student's t-test. The STATISTICA package version 10.0 (Aliso Viejo, CA) was used for statistical analyses. A blinded, trained researcher performed all analyses. Figures were produced using GraphPad Prism 9.1 (San Diego, CA).
Results
Figure 2 shows weight gain (Fig. 2A), food intake (Fig. 2B), feed efficiency ratio (Fig. 2C), and final feed efficiency (Fig. 2D) of AL18 and CR18 groups during the 18 months of CR protocol. As expected, compared to AL18, the CR18 group presented ∼43% lower weight gain (p = 0.001) and ∼18% higher feed efficiency (p = 0.01) from 21 days to 18 months of age.
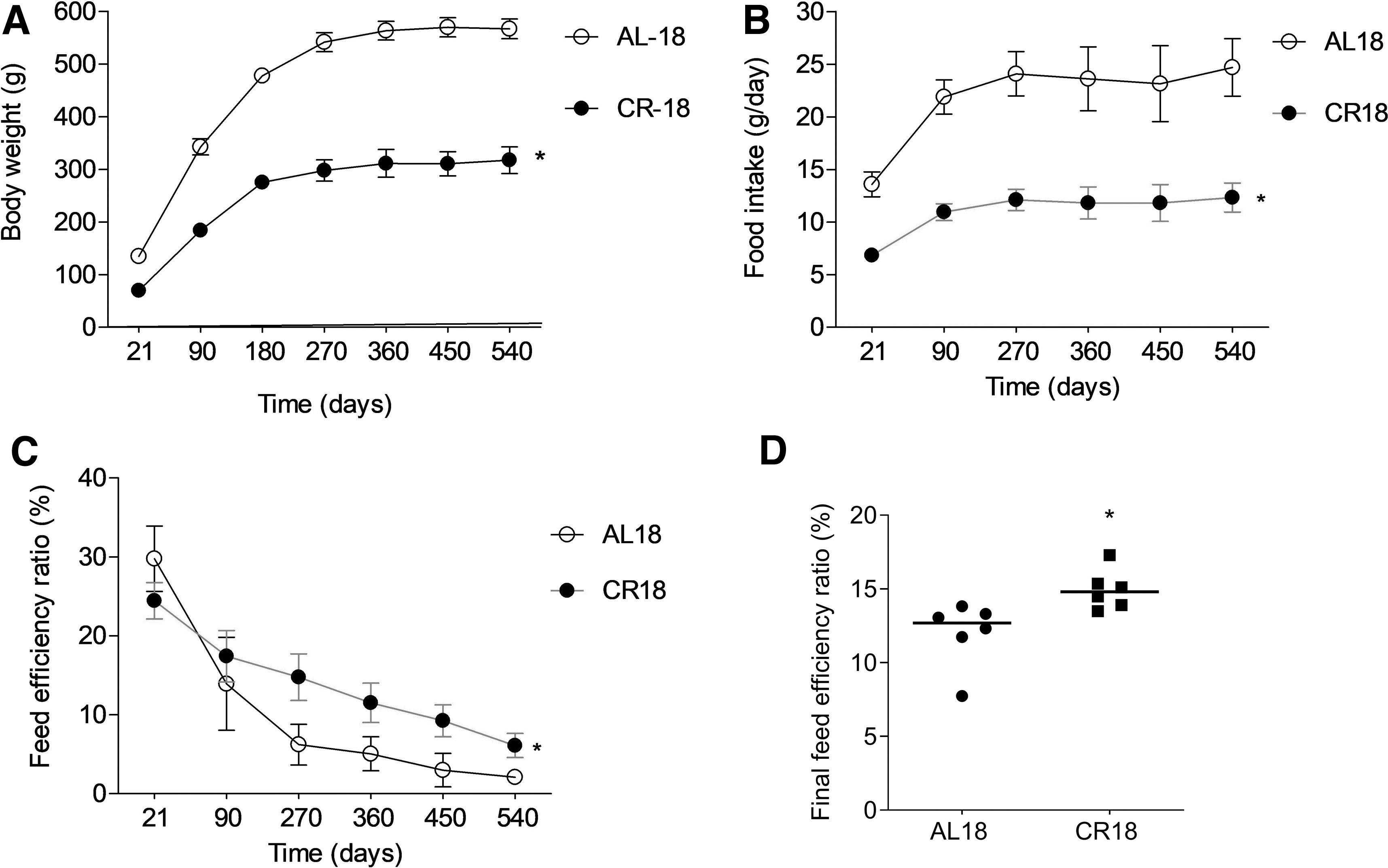
Food consumption and body weight. Food intake
Table 1 presents cardiometabolic risk factors and organ weight data. Compared with AL18, CR18 rats had ∼13% lower fasting glucose (p = 0.002), ∼17% lower OGTT (p = 0.001), ∼23% lower serum glucose levels during the IpITT (p = 0.001), ∼6% lower resting metabolic rate (p = 0.02), ∼10% lower blood pressure (p = 0.01), ∼15% lower heart rate (p = 0.001), ∼37% lower triglycerides (p = 0.003), ∼42% lower cholesterol (p = 0.04), ∼38% lower VLDL-C (p = 0.003) levels, and ∼32% (p = 0.01) lower atherogenic and cardiovascular risk indexes. AL18 and CR18 groups showed similar HDL and LDL-C levels. CR18 group also presented an anti-inflammatory profile compared to the AL18 group. Compared to AL18, CR18 rats had ∼13% (p = 0.01) and ∼12% (p = 0.04) lower IL-6 and TNF-α serum levels, respectively. IL-10 levels did not differ between the groups.
Effects of 50% Caloric Restriction from Birth on Cardiovascular Risk Factors in 18-Month-Old Rats
Data are presented as mean ± SD. N = 8/group. Student's T-test. AL18: 18-month-old AL group. CR18: 18-month-old calorie-restricted group.
p < 0.05 (AL18 vs. CR18).
AL, Ad Libitum; CR, caloric restriction; CRI, cardiovascular risk index; HDL, high-density lipoprotein; IpITT, intraperitoneal insulin tolerance test; OGTT, oral glucose tolerance test; SD, standard deviation; TG, triglyceride; VLDL, very-low-density lipoprotein.
Moreover, the IL-6/IL10 and TNF-α ratios were, respectively, ∼17% (p = 0.009) and ∼18% (p = 0.008) lower in CR18 rats than in AL18 rats. The most striking result was the effect of calorie restriction on visceral fat weight. The CR18 group had from six to seven times lower visceral fat weight than the AL18 group (p = 0.0001), and, even after calculating the fat/body weight ratio, CR18 presented from three to four times lower visceral fat weight than AL18 (p = 0.0001). CR18 also had lower liver (p = 0.0001) and heart (p = 0.0001) weights than AL18, but the liver/body weight and heart/body weight ratios were similar between the groups.
To investigate the basal heart function (without neurohormonal influence), we used the Langendorff technique. Figure 3 shows cardiac function data of the AL18 and CR18 groups. The cannulation of four hearts from AL18 rats and two hearts from CR18 rats had technical pitfalls; thus, these records were discarded. Compared with AL18 group, the CR18 group showed a ∼50% increase in cardiac contractility (Fig. 3A; +dP/dt; AL18: 979.5 ± 197.4, CR18: 1959.0 ± 384.5 mmHg/(s(g); p = 0.0004) and a ∼57% increase in cardiac relaxation (Fig. 3B; −dP/dt; AL18: 512.6 ± 144.5, CR18: 900.3 ± 236.1 mmHg/(s(g); p = 0.008). The intrinsic heart rate was similar between the groups (Fig. 3C; AL18: 185 ± 49, CR18: 200 ± 21 b.p.m.).

Basal cardiac function and incidence of ventricular arrhythmia and fibrillation. Cardiac contractility
To further investigate the cardiac effects of intense calorie restriction from birth in middle-aged rats, we analyzed the ventricular arrhythmia and fibrillation incidence in 18-month-old rat hearts (CR18). Figure 3D and E illustrates representative traces of the pressure pulses from isolated heart data of CR18 and AL18 rats. Compared with the AL18 group, the CR18 group presented nearly three times less incidence of ventricular arrhythmia (Fig. 3F; AL18: 194.8 ± 45.49, CR18: 76.83 ± 51.19 U; p = 0.005) and nearly five times less ventricular fibrillation (Fig. 3G; AL18: 17.25 ± 9.50, CR18: 3.00 ± 6.98 U; p = 0.02).
Next, we delve into the consequences of the CR protocol on AL18 and CR18 rats' heart morphology by assessing the myocardium collagen content and cardiomyocyte morphometry. Figure 4 shows representative photomicrographs of the myocardium of AL18 and CR18 rats stained with hematoxylin/eosin (Fig. 4A, B) and Masson's trichrome (Fig. 4C, D). CR18 hearts presented ∼19% lower cardiomyocyte diameter (Fig. 4E; AL18: 18.97 ± 1.73, CR18: 15.41 ± 0.84 μm; p = 0.003) and ∼61% lower collagen deposition (Fig. 4F; AL18: 2.47 ± 1.07, CR18: 0.95% ± 0.37%; p = 0.03) than AL18 hearts.

Cardiac histologic data. Representative photomicrographs of the myocardium from AL18 and CR18 rats stained with hematoxylin/eosin
Finally, to further investigate the cardioprotection mechanism of CR, we analyzed the redox balance and key intracellular proteins involved with cell survival in the heart. Figure 5 highlights the cardiac redox balance from AL18 and CR18 animals. Redox biomarkers analysis revealed that, compared with AL18 rats, hearts from CR18 had ∼31% lower lipid peroxidation (Fig. 5A; AL18: 1.88 ± 0.41, CR18: 1.29 ± 0.10 nmol/MDA (mg de protein); p = 0.01), and ∼23% higher total antioxidant capacity (Fig. 5B; AL18: 22.48 ± 1.46, CR18: 29.41 ± 6.73 nM FeSO4/mg protein; p = 0.04).
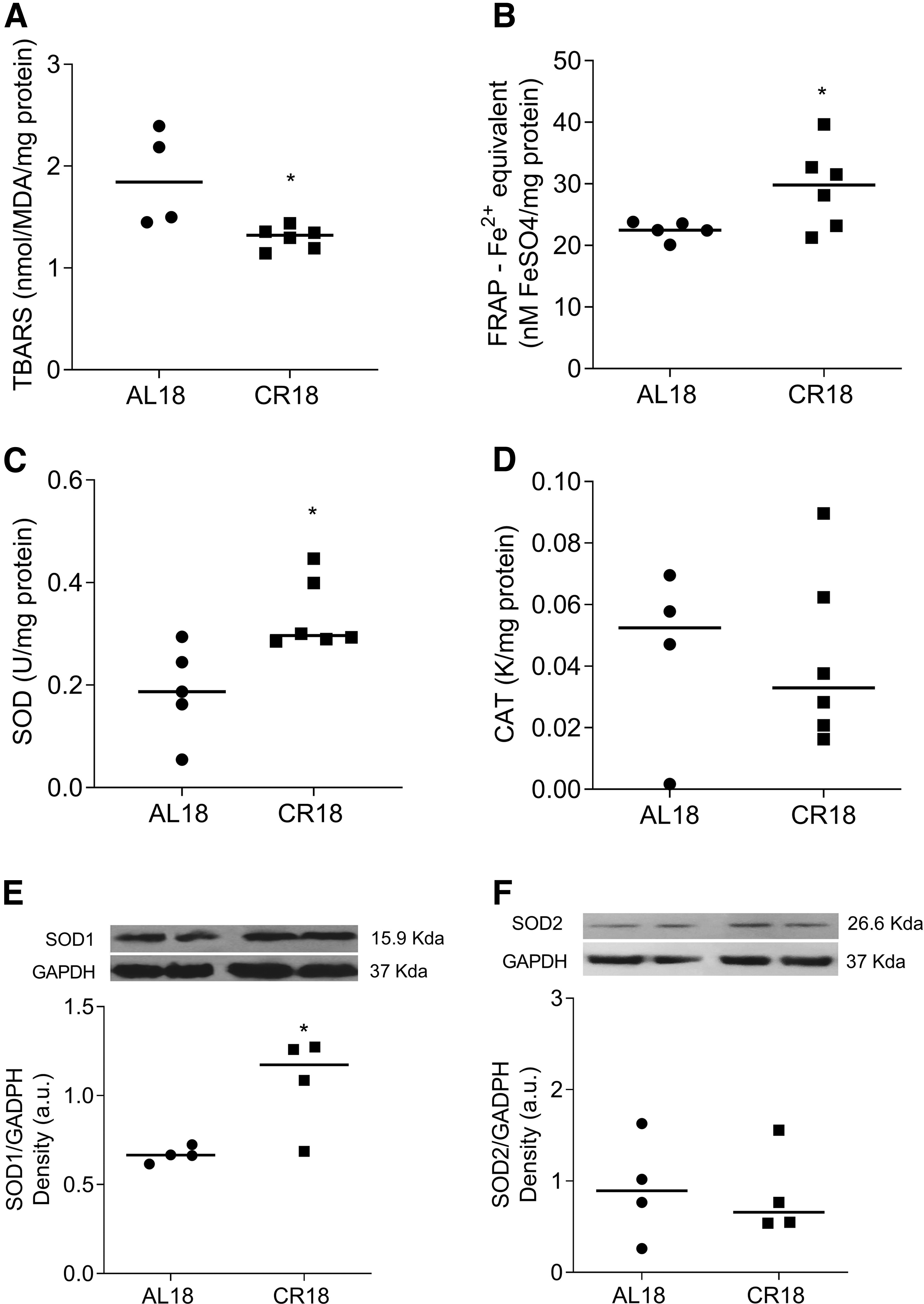
Cardiac redox balance. Lipid peroxidation showed by T-BARS levels
Compared with AL8, CR18 hearts presented a higher superoxide dismutase activity (Fig. 5C; AL18: 0.18 ± 0.09, CR18: 0.33 ± 0.06 U/mg protein; p = 0.01), but no differences were seen in catalase activity between groups (Fig. 5D; AL18: 0.04 ± 0.02, CR18: 0.04 ± 0.02 U/mg protein). The increase in SOD1 content (Fig. 5E; AL18: 0.66 ± 0.05, CR18: 1.07 ± 0.27 a.u.; p = 0.04) seems to explain the improvement of superoxide dismutase activity found in CR18 hearts since cardiac SOD2 content was similar between CR18 and AL18 rats (Fig. 5F; AL18: 0.91 ± 0.56, CR18: 0.85 ± 0.48 a.u.; p = 0.86).
Figure 6 shows expression levels of intracellular proteins involved with cell survival in the hearts of AL18 and CR18 rats. Despite similar content of phospho-AMPK (Fig. 6A; AL18: 0.94 ± 0.05, CR18: 0.83 ± 0.12 a.u.; p = 0.14), compared with AL18, CR18 hearts presented higher levels of HSP72 (Fig. 6B; AL18: 0.75 ± 0.10, CR18: 1.03 ± 0.18 a.u.; p = 0.03) and phospho-ERK (Fig. 6C; AL18: 0.45 ± 0.12, CR18: 1.06 ± 0.33 a.u.; p = 0.01), but lower levels of phospho-P38 (Fig. 6D; AL18: 1.19 ± 0.08, CR18: 0.89 ± 0.13 u.a.; p = 0.02) and caspase-3 (Fig. 6E; AL18: 0.77 ± 0.15, CR18: 0.48 ± 0.10 a.u.; p = 0.05). These results spotlight a prosurvival signaling pathway in the hearts of middle-aged rats, calorie-restricted since birth.
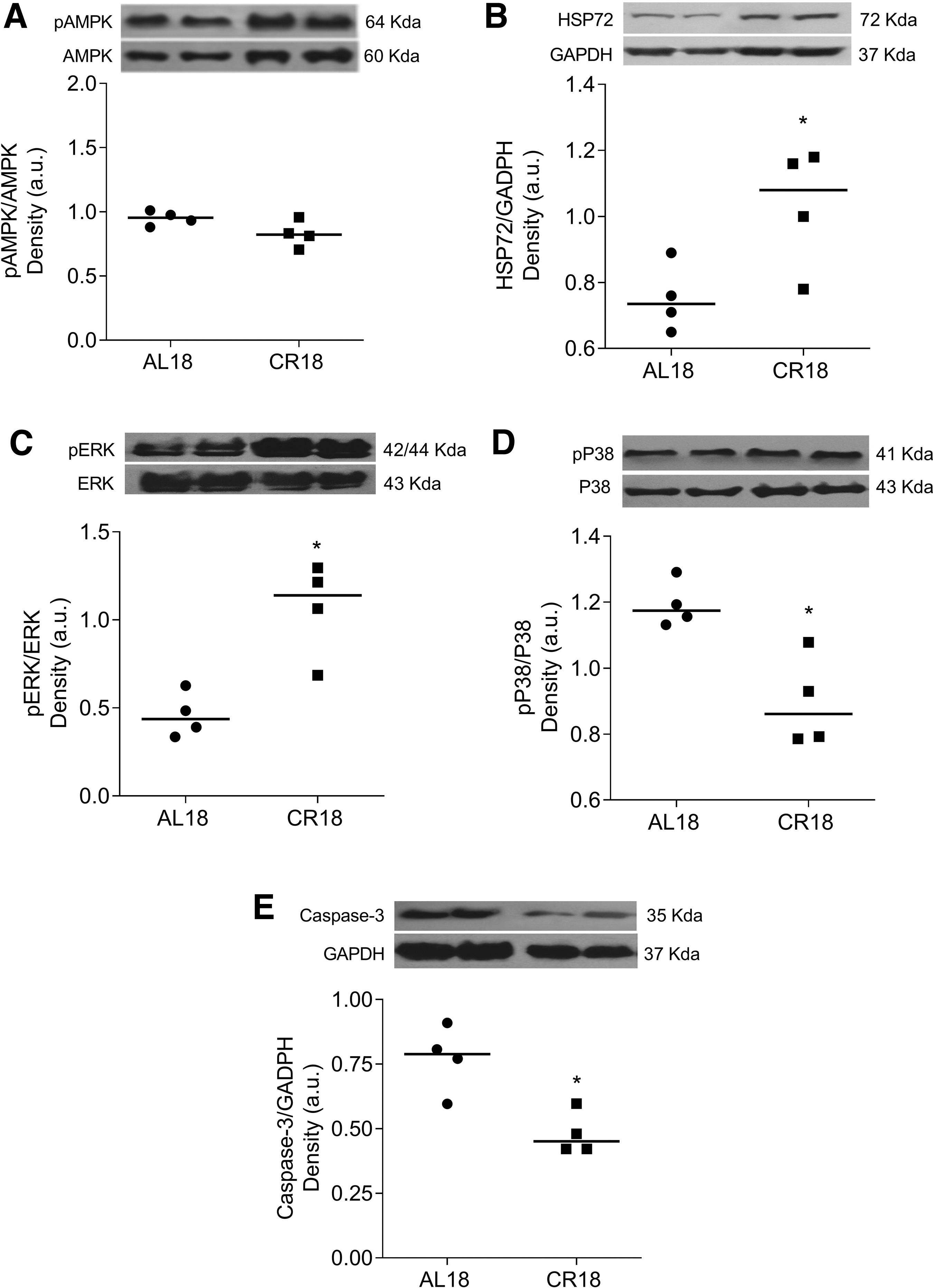
Protein levels of key proteins involved with cardioprotection. Protein levels of phospho AMK
Discussion
This is the first study to investigate the impact of a 50% calorie restriction from birth up to the age of 18 months on the cardiac function, morphology, and cardiac intracellular proteins involved with redox status and cell survival of rats. Our study provides evidence that a 50% CR from birth reduces multiple cardiometabolic risk factors, improves cardiac function and morphology, enhances antioxidative responses, and may activate prosurvival pathways in the heart (Fig. 7).

Key takeaways. Venn diagram summarizing the main findings of the study.
The decline in resting metabolic rate has been generally well accepted as a key determinant of the beneficial cardiometabolic effects of long-term CR. 31 In previous studies with young rats, a 50% CR from birth improved feed efficiency energy, an indirect measurement of energy efficiency improvement. In the current study in aged rats, we measured the resting metabolic rate and found that CR18 animals had a 6% lower resting metabolic rate than AL rats. Thus, it is tempting to speculate that this reduction in the metabolic rest rate triggers the improvement of cardiometabolic health in the CR18. We hypothesize that if animals are calorie-restricted from birth, their cells will adapt to energy deprivation, thus working more efficiently, producing less reactive oxygen species, resulting in less cell damage and increased lifespan. This hypothesis has frequently been used to explain the long-term beneficial effects of CR. 32,33
This study revealed a marked increase in cardiac function and a decrease in cardiomyocyte hypertrophy and cardiac fibrosis in CR18 rats, similar to what was observed in young rats 50% calorie restricted from birth. 17 Melo et al. found that the improvement in cardiac contractility/relaxation produced by the 50% CR from birth in 3-month-old rats was independent of alterations in cardiomyocyte calcium handling. Instead, the improvement in cardiac function in CR rats was associated with cardiac morphological alterations, such as a decrease in cardiomyocyte growth and cardiac fibrosis. As in the current study, other authors also found that CR prevents cardiomyocyte hypertrophy and cardiac fibrosis deposition in middle-aged rats. 34 –36
A novel finding from this study was that a 50% CR from birth prevented ventricular arrhythmia and fibrillation in aged rat hearts. Interestingly, these beneficial cardiac effects did not occur when rats were subjected to an intense CR during a short-term period. In a recent study, Almeida et al. 3 found that, compared to AL rats, isolated hearts from 4-month-old rats subjected to a 60% CR for 14 days presented an increase in ventricular arrhythmia and fibrillation. Moreover, hearts from these CR rats revealed a pathologic profile such as myofibrillar disarray and cardiomyocyte apoptosis. Three main points distinguish this current study from the studies by Almeida et al. 3 Almeida et al. 3 used younger rats; their CR protocol was more intense, and, most importantly, CR was imposed in a short-term period.
Another difference was that Almeida et al. 3 obtained the data from ischemia/reperfusion (I/R) experiments, while in the current study, cardiac function records were obtained at basal conditions. Thus, one may argue that if the hearts of CR rats in our study were subjected to the insult of I/R, they may exhibit a higher incidence of ventricular arrhythmia/fibrillation than the hearts of rats in the AL group. Although we did not perform I/R experiments, we believe that CR18 hearts would be protected from I/R insults. In our previous study, we found that the hearts of 6-month-old rats 50% calorie restricted from birth were significantly protected from the cardiac function decline induced by I/R insult. Nevertheless, the absence of I/R experiments is an important limitation of the current study. Another limitation was not accessing cardiac electrophysiological data since the majority of arrhythmia and fibrillation events in aged rat hearts are caused by electrophysiological abnormalities.
The imbalance of cardiac redox status has been extensively related to cardiac dysfunction in aged rats. 37 –39 Overall, a decrease in antioxidant defense or an elevation in oxidative stress is closely linked to cardiac dysfunction in aged rats. Numerous studies have reported that calorie restriction enhances antioxidant defense and reduces oxidative stress in the heart of aged. 1,8,35,36,40 In the present study, we revealed that even when the calorie restriction is imposed since birth, an improvement in redox homeostasis in the heart of middle-aged rats is found. The improvement in the antioxidant profile in CR18 hearts may be a consequence of an increase in SOD activity since catalase activity was similar between CR18 and AL18 hearts. Moreover, the CR18 hearts had an upregulation of multiple prosurvival pathways closely related to the improvement of cell antioxidant defense.
Previous studies showed an increase in AKT phosphorylation 16 and HSP72 content 17 in hearts of young rats 50% calorie-restricted from birth. This study also revealed an enhancement of intracellular HSP72 content in the CR18 hearts and revealed new findings showing that the CR18 heart presents an increase in ERK ½ phosphorylation and a reduction in P38 phosphorylation and Caspase-3 content. The improvement in cardiac HSP72 content has been considered a key pathway against cellular oxidative damage by increasing the SOD activity 17 and the resistance to the opening of mitochondrial permeability transition pores, reducing reactive oxygen species release. 41,42 In addition, the increased levels of HSP72 and phospho-ERK ½ trigger prosurvival pathways by reducing p38 and caspase-3 activation. 43 –45 However, these results must be interpreted with caution, since phospho-ERK ½ and p38 may activate various cell signaling pathways other than prosurvival pathways.
Study limitations
Several limitations should be acknowledged in this study. Since female rats severely calorie-restricted from birth may present cardiac function impairment in adulthood, 46 the results of this study should be applied just to male rats. The sample size was relatively small, especially in the cardiac function experiments, due to some outliers' values detected during statistical analyses. Moreover, we cannot assure that the results obtained from isolated hearts in this study would match those obtained from in vivo cardiac function analysis (e.g., echocardiographic analyses). The incorporation of electrophysiological assessments, such as patch clamp experiments, is also essential for accurately detecting and analyzing the ionic currents associated with the incidence of ventricular arrhythmia and fibrillation. Also, in vivo analyses of autonomic function may help understand the beneficial effects of our CR protocol on the heart, especially on ventricular arrhythmia and fibrillation incidence.
Although we analyzed isolated hearts, CR from birth may inhibit the increased sympathetic activity, which is commonly observed with aging, decreasing the susceptibility of the heart toward arrhythmia and fibrillation. However, the Langendorff technique provides highly reproducible results regarding cardiac function without humoral and neuronal influences, revealing the potential effects of different treatments and interventions of interest on the heart. 47 CR appears to increase longevity and reduce cardiac aging by different mechanisms beyond that investigated in the current study, such as by boosting sirtuins 31 and signaling pathways involved with autophagy, 40 and mitochondrial biogenesis. 48 Moreover, we did not measure other caspases besides caspase-3 that would interfere with survival and apoptotic signaling. Thus, additional studies are required into the intracellular mechanisms underlying the cardioprotection induced by CR from birth in aged hearts.
Implications for clinical practice
The results of this study cannot be extended to human studies; in fact, transferring this CR protocol to humans is complex. Adverse effects of this CR protocol, such as growth retardation, cannot be ruled out. Thus, the findings of this study should be interpreted with caution. From a clinical standpoint, it is currently unfeasible to implement a long-term, intense CR protocol starting from birth. However, the implementation of less intense CR protocols, conducted over an extended period of time, starting from early stages of life, such as from childhood to senility, may provide valuable insights into the impact of CR on risk factors for cardiovascular diseases. In addition, research into the intracellular mechanisms activated by long-term calorie restriction protocols may assist in the development of effective therapeutic strategies, such as CR mimics, target to reversing or preventing age-related cardiac diseases.
Future preclinical and clinical studies are warranted to evaluate the impact of more realistic calorie restriction protocols implemented from early life stages to senescence on the development and progression of cardiovascular diseases.
Conclusions
Collectively, our data reveal for the first time multiple positive effects of a 50% CR since birth in middle-aged rats. The CR protocol clearly prevented the decline of cardiac function, reduced the incidence of ventricular arrhythmia and fibrillation, inhibited cardiomyocyte hypertrophy and the deposition of collagen in the myocardium, enhanced antioxidant defense, and improved the intracellular levels of distinct proteins involved with cell survival in the heart.
Footnotes
Authors' Contributions
D.d.S.M., L.C.P., C.S.S., B.F.M., I.F.M.K., L.G.M., and B.C.C.G.: conducted research (hands-on conduct of the experiments and data collection). R.C.C., E.A.E., E.R.V., F.d.C.M., and L.d.S.A.C.: provided essential reagents and equipment for the research and hands-on conduct of the experiments and data collection. R.A.L.D.S., K.H.S., and I.P.Q.: analyzed data, performed the statistical analysis, and had primary responsibility for the final content. M.F.D.P.: designed research (project conception, development of overall research plan, and study oversight) and wrote the article. All authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
We thank the Centro Integrado de Pós-Graduação e Pesquisa em Saúde, (CIPq-Saúde) from the Universidade Federal dos Vales do Jequitinhonha e Mucuri (UFVJM) for providing equipment and technical support for experiments. We thank the Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG) (Nos. APQ-00214-21, APQ-01049-21, APQ-00938-18, APQ-03855-16), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) (No. 438498/2018-6), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES)—Finance code 001 for providing financial support.