
Abstract
Parishin, a natural compound, has demonstrated significant potential in mitigating age-related phenotypes and improving outcomes in age-associated diseases. Given that aging is a major risk factor for numerous chronic conditions, including pulmonary fibrosis, we investigated parishin’s effects on cellular senescence and lung health. In our study, we treated mouse lung epithelial cells with parishin and observed a reduction in cellular senescence markers alongside an upregulation of sirtuin 1 (SIRT1). Building on these in vitro findings, we administered parishin to naturally aged mice. The treatment resulted in decreased pulmonary fibrosis and reduced DNA damage in lung tissue. Notably, we found that parishin treatment led to a reduction in Cluster of differentiation 38 (CD38) levels, concomitant with an increase in SIRT1 expression. These findings indicate that parishin may enhance lung function in aged mice, suggesting its potential as a therapeutic agent for treating age-related pulmonary disorders.
Introduction
Parishin, a phenolic glucoside and one of the major constituents of Gastrodia elata, is composed of three gastrodin molecules esterified with the terminal carboxyl groups of citric acid. Our research team has successfully patented the extraction and preparation process of parishin. Parishin has been shown to significantly extend the replicative lifespan of yeast by regulating the Sir2/Uth1/TOR signaling pathway. 1 Recent studies have explored the potential health benefits of parishin, particularly in aging and age-related diseases. Parishin has been found to ameliorate aging-induced “leaky gut” conditions by improving gut barrier function and reducing gut inflammation. 2 In addition, parishin upregulated Klotho, a protein of age-related declines, in the serum and vascular tissue of naturally aged mice, thereby alleviating vascular endothelial cell senescence and vascular aging. 3 Furthermore, parishin treatment improved cardiac function and ameliorated aging-induced cardiac injury, hypertrophy, and fibrosis in naturally aged mice. 4 More research is needed to fully understand its mechanism in age-related diseases.
Pulmonary fibrosis is a chronic lung disease that leads to a progressive decline in lung function, with its risk increasing with age. The disease is more prevalent in older adults. 5,6 Senescent cells secrete factors known as the senescence-associated secretory phenotype, which can promote tissue remodeling and fibrosis. 7,8 With age, mitochondrial function declines, leading to increased oxidative stress and inflammation in the lungs. Growing evidence suggests that mitochondrial dysfunction plays a significant role in the pathogenesis of pulmonary fibrosis. 9,10 CD38, an NADase enzyme, is induced by factors secreted from senescent cells. Increased CD38 expression in aging tissues is linked to age-related cellular Nicotinamide adenine dinucleotide (NAD+) decline. 11,12 Elevated CD38 activity can lead to reduced NAD+ levels, which contribute to oxidative stress and mitochondrial dysfunction. 13 CD38 also diminishes CD8+ T cell function by negatively affecting mitochondrial fitness through the inhibition of multiple steps of mitochondrial biogenesis and function. 14 CD38 inhibitors can block CD38 activity, thereby maintaining NAD+ levels and ameliorating aging-related phenotypes. 15 Elevated CD38 levels in pulmonary fibrosis downregulate intracellular NAD, leading to the impairment of NAD-dependent cellular and molecular activities, which likely promotes aging. Both genetic and pharmacological inactivation of CD38 have been showed to ameliorate pulmonary fibrosis. 16 Therefore, CD38 is a potential therapeutic target for treating pulmonary fibrosis.
In this study, we investigated whether parishin could improve pulmonary fibrosis and reduce age-related lung phenotypes. We found that parishin alleviated cellular senescence in mouse lung epithelial cells (MLE-12) and upregulated SIRT1 levels. In naturally aged mice, parishin reduced CD38 expression to ameliorate pulmonary fibrosis.
Materials and Methods
Parishin is provided by Qi Jianhua’s research team at Zhejiang University.
Cell culture
MLE-12 cells were obtained and cultured using an Epithelial Cell Medium (SCIENCELL, California, USA) containing 10% fetal bovine serum. The culture medium was changed every 24 hours. When MLE-12 were 70%–80% confluent, the cells were trypsinized, resuspended in the culture medium, and seeded into 6-well microplates for each assay. Cell senescence was evaluated by senescence-associated beta-galactosidase (SA-β-gal) staining, and the protein levels of SIRT1, PGC-1α, γH2AX and CD38 were assessed by Western blotting.
Drug administration
Parishin powder was dissolved in Dimethyl Sulfoxide(DMSO) and diluted into the final concentration of 10 and 30 μM by medium for cell intervention. A control group was set to add the corresponding dose of DMSO. The cells were randomly divided into a blank control group, parishin middle-concentration intervention group (10 μmol/L parishin), and parishin high-concentration intervention group (30 μmol/L parishin). MLE-12 cells underwent a 2-week parishin treatment, followed by cellular senescence induction using a 24-hour exposure to 10 µg/mL bleomycin, while parishin treatment was sustained throughout this induction. A wound-healing assay was conducted using bleomycin-induced senescent MLE-12 cells. Initially, MLE-12 cells underwent a 2-week treatment with parishin, followed by the induction of cellular senescence through a 24-hour exposure to 10 µg/mL bleomycin, while parishin treatment was maintained. After the bleomycin exposure, a scratch was created to wound the cells, with parishin treatment continuing throughout the process. Cells were incubated for 3–4 weeks using the corresponding preformulated media as described above.
Animals
Parishin was provided by Qi Jianhua’s research team at Zhejiang University (Patent No. of Zhejiang University CN201610061288.2). Nineteen-month-old specific pathogen free healthy male C57BL/6 mice (n = 30) were purchased from Zhejiang Laboratory Animal Center. All mice were fed with a normal diet and housed under a 12‐hour light and 12‐hour dark cycle (7 am and 7 pm, 25°C, and ∼70%–80% humidity) at the Laboratory Animal Center of the Medical Department of Zhejiang University. After a 2-week acclimation period, the mice were randomly divided into three groups (n = 10 each group) as follows: aged control group (aging), aged middle-dose parishin treatment mice (Par[M], parishin 20 mg/kg/day), and aged high-dose parishin treatment mice (Par[H], parishin 30 mg/kg/day]. The parishin was dissolved in 0.9% saline (containing 0.5% DMSO) and administered orally by gavage to the parishin group daily, while the aging group was administered the same volume of 0.9% normal saline each day. At week 8, the mice were euthanized to collect lung tissues for further analysis. All experiments were conducted following “The Instructive Notions with Respect to Caring for Laboratory Animals” issued by the Ministry of Science and Technology of the People’s Republic of China. The study protocols were approved by the Committee on the Ethics of Animal Experiments of Zhejiang University (approval number: 2020 Experimental Kuaishen No. 1446).
SA-β-gal activity assay
SA-β-gal activity was measured using a commercially available kit (Thermo Fisher Scientific, K145501), following a protocol adapted for enhanced sensitivity and specificity in detecting senescence. Briefly, MLE-12 cells were washed thrice with phosphate-buffered saline (PBS) and then fixed with a fixative solution for 15 minutes at room temperature. Subsequently, the cells were incubated overnight at 37°C with fresh SA-β-gal staining solution adjusted to pH 6.0. To ensure an accurate assessment of senescence, we included a control group of untreated cells to differentiate between basal and senescent β-galactosidase activity. Positively stained cells were counted in a sample of 200 total cells at 100× magnification, and the percentage of SA-β-gal-positive cells was calculated. Any differentiation between senescent and nonsenescent cells was also noted, where necessary, based on staining intensity and morphology.
Western blot analysis
Protein concentration was determined using a BCA Protein Assay Kit (Beijing Kangwei Century Biotechnology Co., Ltd.). Equal amounts of proteins (30 µg) were placed in 4%–20% Sodium Dodecyl Sulfate-Polyacrylamide Gel Electrophoresis (SDS‐PAGE) gel (GenScript Biotechnology) and transferred to Polyvinylidene Fluoride (PVDF) membrane (Bio-Rad) by semiwet transfection for 15 minutes. The membranes were blocked with the QuickBlock fast blocking solution (Beyotime Biotechnology) for 15–20 minutes and incubated with γH2AX (ab 11174) and GAPDH at 4°C overnight. Then, membranes were washed in Tris-Buffered Saline with Tween (TBST) and incubated with the appropriate secondary antibodies at room temperature for 1 hour. Subsequently, membranes were washed in TBST and visualized using the Omni-ECL Basic Chemiluminescence Detection Kit (Shanghai Ya Enzyme Biomedical Technology Co., Ltd.). The housekeeping protein GAPDH was used as a loading control.
Masson’s trichrome staining
The mouse vascular tissues were fixed in 4% paraformaldehyde (Beijing Solarbio Technology Co., Ltd.) at room temperature for 24 hours and then embedded in paraffin and cut into 4-μm thick sections. Masson’s trichrome staining (MS) was used to visualize the lung fibrosis. The slices were examined under an inverted light microscope (Leica, Berlin, Germany). The degree of vascular fibrosis was quantified by ImageJ software. The degree of vascular fibrosis is expressed as a percentage of the fibrotic area of the whole region.
Immunohistochemistry
Four-micrometer thick paraffin sections were deparaffinized in xylene and sequentially rehydrated using a graded series of ethanol at 100°C Ethylenediaminetetraacetic Acid (EDTA) (PH = 9) alkaline repair for 20 minutes. After rinsing in 1× PBS (Leica) for 15 minutes (three times, 5 minutes each time), the sections were blocked with 3% hydrogen peroxide for 10 minutes. The sections were rinsed again with 1× PBS for 15 minutes. The sections were incubated with α-smooth muscle actin (α-SMA) and CD38 antibody (1:100) overnight at 4°C. Then, the sections were rinsed with PBS for 15 minutes and incubated with the appropriate secondary antibodies (Leica, Berlin, Germany) for 15 minutes at Room Tempreture (RT). After rinsing with 1× PBS for 15 minutes, immunoreactivity was detected with 3,3′-diaminobenzidine substrate (Leica) for 10 minutes, and the samples were washed with 1× PBS for 15 minutes. Hematoxylin dye solution was added to the stain at room temperature for 3 minutes, and the slides were mounted with neutral gum. All the above steps were performed on Leica Bond RX (Leica, Sydney, Australia). The sections were then viewed using microscopy (magnification: ×400) (Leica).
Statistical analysis
All data are reported as mean ± SD. Each experiment was repeated at least thrice independently. The differences in mean values between the two groups were assessed by the Student’s t-test. Statistically significant differences were evaluated at p < 0.05.
Results
Parishin alleviated the senescent phenotypes in MLE-12 cells
Our previous studies reported that parishin alleviated vascular aging in mice by upregulation of Klotho. 3 In this study, we investigated the function of parishin in MLE-12, which were induced to cellular senescence by bleomycin. 17 Briefly, MLE-12 cells underwent a 2-week parishin treatment, followed by cellular senescence induction through a 24-hour exposure to 10 µg/mL bleomycin, while parishin treatment was sustained throughout this induction. Parishin notably reduced bleomycin-induced cellular senescence compared with cells exposed to bleomycin without parishin (Fig. 1A and B). We conducted a wound-healing assay using bleomycin-induced senescent MLE-12 cells. Initially, MLE-12 cells underwent a 2-week treatment with parishin, followed by the induction of cellular senescence through a 24-hour exposure to 10 µg/mL bleomycin, while parishin treatment was maintained. After the bleomycin exposure, a scratch was created to wound the cells, with parishin treatment continuing throughout the process. 18 Images of the cells in the scratched area were captured at 0- and 24-hour postwounding. Parishin-treated cells from the bleomycin-induced senescent MLE-12 group showed substantially elevated proliferation in both the Par(M) and Par(H) groups compared with the BLM group (Fig. 1C), suggesting that parishin-treated cells had a stronger repair ability than control cells.
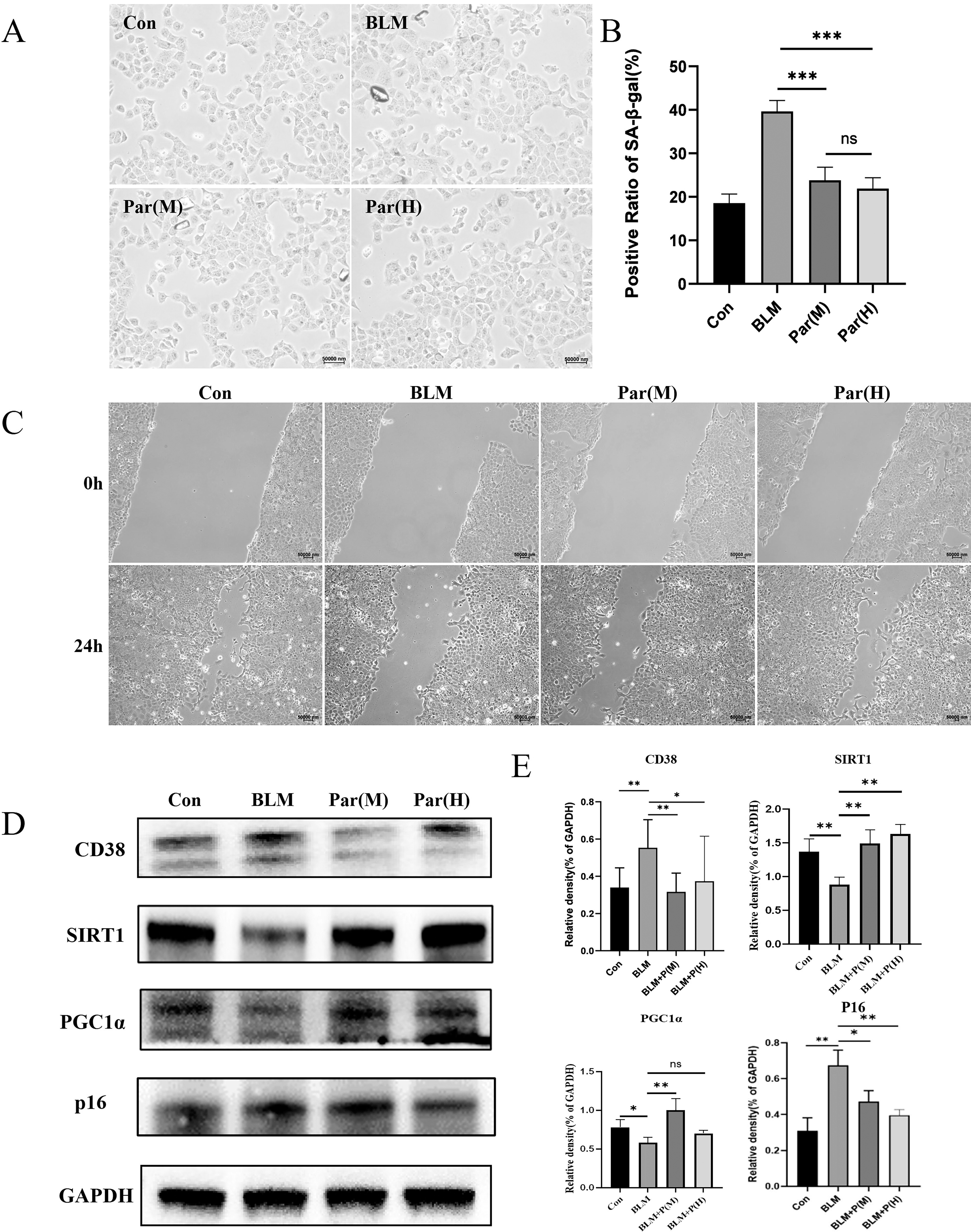
Parishin alleviated the senescent phenotypes in MLE-12 cells.
SIRT1, a NAD+-dependent deacetylase, plays a crucial role in mitochondria function such as promoting mitochondrial biogenesis, protecting mitochondria from oxidative stress, and improving mitophagy. 1 To evaluate the effects of parishin on mitochondria function in MLE-12 cells, we performed Western Blot (WB) assays. We found that parishin treatment stimulated the expression of SIRT1 (Fig. 1D). SIRT1 promotes mitochondrial biogenesis by activating Peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α), a key regulator of mitochondrial function. In addition, PGC-1α expression was upregulated in parishin-treated groups compared with bleomycin-induced cells (Fig. 1D). Taken together, these findings suggest that parishin treatment reduced SA-β-Gal signals and increased SIRT1 expression, thereby maintaining cellular health and mitochondrial function in MLE-12 cells.
Parishin improved pulmonary fibrosis in naturally aged mice
To examine the function of parishin in the lungs of naturally aged mice, we selected naturally aged C57B/L6 mice (19 months, 20–30 g), which were randomly divided into three groups (N = 10), aging control (aging), middle dose (Par[M], 20 mg/kg/day), and high dose (Par[H], 30 mg/kg/day). Three young mice served as young control (young). Parishin was dissolved in 0.9% saline and administered as a suspension by the gavage method. Treatment was continued for 8 weeks, once daily. MS was performed on lung sections for the young, aging, Par(M), and Par(H) groups. Collagen was stained and appeared blue, which visualized fibrosis in lung tissues. The data showed that collagen deposition was significantly increased in aged lungs compared with the young group, whereas it was slightly decreased in the Par(M) group and significantly decreased in the Par(H) group (Fig. 2A). Pulmonary fibrosis is characterized by myofibroblast expansion. alpha-smooth muscle actin (α-SMA) expressing cells, termed myofibroblasts, are expanded and are considered a major risk factor for pulmonary fibrosis. 19 To assess the effect of parishin treatment on pulmonary fibrosis, we examined α-SMA expression by immunohistochemistry. The results showed that parishin improved pulmonary fibrosis by reducing the expression of α-SMA (Fig. 2B and C).

Parishin improved pulmonary fibrosis in naturally aged mice. (
To assess the potential effect of parishin on DNA damage response factor, we collected lysates from untreated and treated lung tissues of young and aged mice. Immunoblot analysis indicated that γH2AX was markedly reduced upon parishin treatment. These results indicated that parishin mitigated senescent phenotype by decreasing γH2AX levels (Fig. 2D and E).
Parishin reduced CD38 expression in naturally aged mice
CD38 is a NADase that hydrolyzes NAD+ into nicotinamide and Adenosine Diphosphate (ADP)-ribose, which can reduce NAD+ levels. SIRT1 is a NAD+-dependent deacetylase enzyme that plays a crucial role in various cellular processes. 20 CD38 suppresses the activity of SIRT1 and negatively affects mitochondrial function and ability. 14,21 Both genetic and pharmacological inactivation of CD38 can ameliorate pulmonary fibrosis. 16 To assess the effect of parishin on the lungs of naturally aged mice, we examined the impact of parishin administration on lung tissues by WB. Parishin decreased the expression of CD38 and increased SIRT1 protein levels in the Par(H) group compared with the aging group (Fig. 3A and B). We also examined CD38 expression by immunohistochemistry. The results showed that parishin reduced CD38 protein levels in the parishin treatment group (Fig. 3C and D). These results suggested that the reduction of CD38 by parishin treatment may improve mitochondrial function and efficiency in lung tissues.
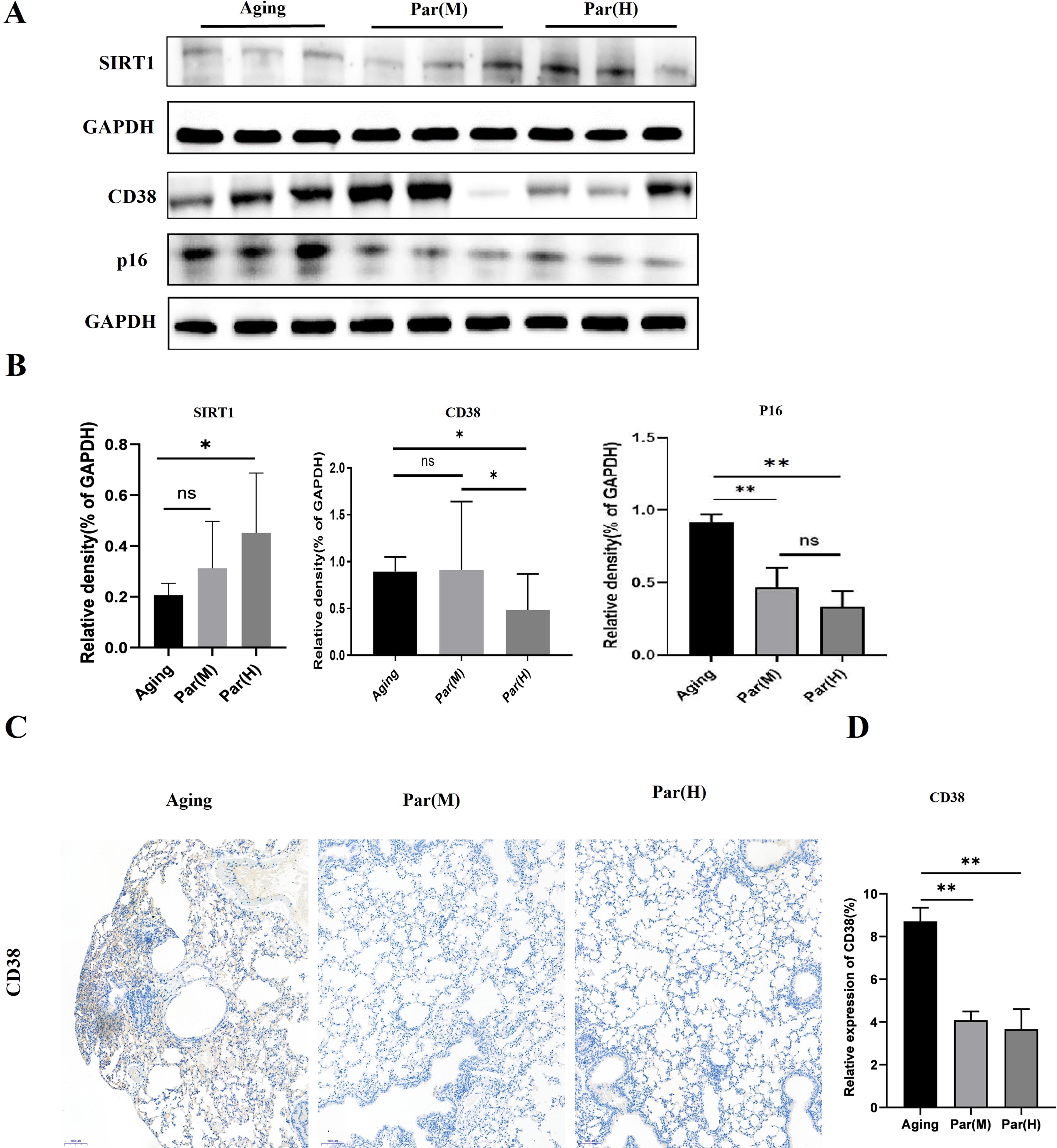
Parishin reduced CD38 expression in naturally aged mice.
Discussion
SIRT1 directly interacts with and activates PGC-1α by deacetylating it. PGC-1α is renowned for stimulating the production of new mitochondria, which extends the health span and protects against age-related metabolic decline. 22,23 Parishin stimulates the expression of SIRT1 and PGC-1α, promoting mitochondrial biogenesis in MLE-12 cells. These observations suggest that parishin may have a potential role in improving mitochondrial function in MLE-12 cells.
Lung fibrosis, characterized by excessive collagen deposition and stiffening of lung tissue, is a serious condition with limited treatment options for older people. We explored the potential of parishin in improving lung fibrosis using MS and α-SMA as key indicators. Parishin treatment reduced the intensity and extent of collagen deposition compared with the aging group. α-SMA is a marker of myofibroblasts, which play a crucial role in fibrosis by producing an excessive extracellular matrix. 24 Parishin decreased α-SMA expression compared with the control group, indicating reduced myofibroblast activation and fibrosis progression. This study provides a strong basis for further investigations into the therapeutic potential of parishin for lung fibrosis treatment.
Mitochondrial dysfunction is associated with the aging process and age-related diseases, including neurodegenerative diseases, metabolic syndrome, diabetes, cardiovascular diseases, and cancer. 25 CD38 modulates cellular metabolism and energy balance by regulating NAD+ levels. CD38 activity contributes to the age-related decline in NAD+ levels. Lower NAD+ levels are linked to impaired mitochondrial function, increased oxidative stress, and reduced sirtuin activity, which are hallmarks of aging and associated diseases. 12 –15,21 The interplay between CD38 and mitochondria is significant due to their roles in NAD+ metabolism. SIRT1, being NAD+-dependent deacetylases, regulates mitochondrial biogenesis and function. CD38-mediated NAD+ depletion can impair SIRT1 activity, affecting mitochondrial health and cellular metabolism. To combat the age-related decline in NAD+ levels and associated mitochondrial dysfunction, parishin reduces CD38 expression, which could preserve NAD+ levels, thereby enhancing mitochondrial function and potentially ameliorating age-related conditions. Parishin offers a novel approach to reduce CD38 expression and enhance NAD+ levels. This study showed that parishin has potential to treat a wide range of age-related diseases. Ongoing research aims to fully elucidate its mechanisms, optimize its efficacy, and ensure its safety.
Conclusions
In this study, parishin has a significant capacity to alleviate senescent phenotypes and improve mitochondrial function in MLE-12 cells, primarily through the upregulation of SIRT1 and PGC-1α. In naturally aged mice, parishin treatment effectively mitigates pulmonary fibrosis and improves overall lung health by reducing collagen deposition, α-SMA expression, and γH2AX levels. Parishin reduces CD38 expression, thereby enhancing SIRT1 activity and promoting better mitochondrial function in lung tissues. These collective findings highlight the potential of parishin as a therapeutic drug against cellular senescence and age-related pulmonary conditions.
Footnotes
Acknowledgments
The authors thank all of the doctors and graduate students who were involved in the study.
Authors’ Contributions
X.X.Z. contributed to conceptualization (equal), data curation (lead), formal analysis (lead), investigation (lead), methodology (lead), project administration (equal), software (lead), validation (lead), visualization (lead), and writing—original draft (lead). S.X.Z. was involved in investigation (supporting), formal analysis (equal), methodology (equal), project administration (equal), resources (supporting), and software (supporting). Z.Y.S. was in charge of investigation (supporting), formal analysis (equal), methodology (supporting), project administration (equal), and resources (supporting). L.L.S. was in charge of software (supporting), methodology (supporting), writing—review and editing (supporting), validation (supporting), and visualization (supporting). Q.Z. was in charge of funding acquisition (supporting), resources (supporting), and writing—review and editing (supporting). Y.Q.L. was involved in conceptualization (lead), funding acquisition (lead), project administration (lead), resources (lead), supervision (lead), and writing—review and editing (lead).
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This study was supported by funding from