
Abstract
Parkinson’s disease (PD) is accompanied by a complex array of nonmotor and motor manifestations. The exploration of anti-inflammatory and antioxidant active ingredient as potential therapeutic interventions in PD-associated mood alterations has gained significant attention. This study aimed to assess the antidepressant and anxiolytic properties of luteolin (LTN), a potent antioxidant and anti-inflammatory component, using a 6-hydroxydopamine (6-OHDA)-induced animal model of PD. Rats were administered LTN (10, 25, and 50 mg/kg, per oral) and fluoxetine (10 mg/kg/per oral) over a 28-day period. Behavioral tests were employed to estimate the depression- and anxiety-like behaviors. Rats treated with LTN exhibited significant improvement in 6-OHDA-induced mood alterations, as per behavioral tests. Additionally, LTN treatment led to increased hippocampal levels of catalase and superoxide dismutase, and a reduction in malondialdehyde. LTN downregulated the gene expression of nuclear factor kappa B (NF-κB)/nod-like receptor (NLR) pyrin domain-containing 3 (NLRP3) axis components, including NF-κB, NLRP3, ASC, and Caspase1 and reduced the protein level of pro-inflammatory cytokines, including interleukin (IL)-6, interleukin (IL)-1β, and tumor necrosis factor alpha (TNF-α), in addition to augmenting the protein levels of TNF-α, IL-1β, and IL-6. Furthermore, LTN exhibited an upregulatory effect on the anti-inflammatory cytokine IL-10 within the hippocampus of 6-OHDA-induced PD rats. Also, molecular docking showed higher affinity between LTN and NF-κB/NLRP3 axis components. These findings highlight the potential anxiolytic and antidepressant impacts of LTN through its antioxidant and anti-inflammatory mechanisms against 6-OHDA-induced alterations in a rat PD model.

Introduction
Parkinson’s disease (PD) is a complex and fast-growing degenerative condition of brain in the world. 1 The number of patients with PD has doubled to more than 6 million from 1990 to 2016. 2 The pathophysiological hallmark of PD is a profound by the loss of midbrain dopamine neuron and, to a lesser extent, of noradrenergic and serotoninergic neurons. 3 The histopathology of PD includes the presence of abnormal and misfolded alpha-synuclein protein (α-syn) aggregates in Lewy bodies and Lewy neurites. 4 The main characteristics of PD include the classical motor manifestations, including bradykinesia, rigidity, and tremor, but patients invariably also have nonmotor symptoms, such as depression, anxiety, cognitive impairment, altered sensations, constipation, or sleeping disturbances. 5,6 Nonmotor symptoms have been identified as substantial determinants that are linked to a decrease in the overall quality of life. Moreover, these symptoms impose a noteworthy economic burden on both the families of the patients and the wider society. 6 Depressive or anxious symptoms are prevalent among individuals diagnosed with PD and often coexist. 7 Research has indicated that approximately 92% of patients with PD diagnosed with anxiety exhibit depressive behavior, while approximately 67% of patients with PD and depression display anxious behavior. 8 In models of PD induced by neurotoxins, there is evidence of depression behaviors. These depression-like symptoms can appear before the onset of motor impairments typically related to PD. 9 There is a well-established association between depression and inflammatory processes. The administration of 6-hydroxydopamine (6-OHDA) has been demonstrated to induce neuroinflammation, which involves the progression of depressive-like symptoms along with PD in animal models. 10 This neuroinflammation is triggered by the secretion of inflammatory markers, including tumor necrosis factor (TNF) and interleukin 1-beta (IL-1β), from activated microglia and astrocytes. 11 The secretion of cytokines depends on the activation of intracellular complexes, such as the nod-like receptor (NLR) pyrin domain-containing 3 (NLRP3) inflammasome, and the maturation of procytokines to cytokines via the caspase 1 (Casp1) component of the inflammasome. 12 Consequently, an activation of cellular mechanisms results in mitochondria-associated cellular dysfunction, oxidative stress (OS), and ultimately leading to the apoptosis of dopaminergic cells. 11 The inflammation in the brain, known as neuroinflammation, occurs following the production of a wide range of pro-inflammatory mediators from activated microglia. 13 To address these interconnected processes, employing antioxidant and anti-inflammatory strategies that target multiple pathways may offer potential benefits in reducing anxiety and depressive behaviors in individuals with PD.
Luteolin (LTN, 3′,4′,5,7-tetrahydroxyflavone) is a flavonoid abundant in various medicinal plants, fruits, and vegetables. 14 It possesses a broad range of pharmacological and biological properties, including neuroprotective, antitumor, antioxidant, anti-inflammatory, and antiallergic effects. 15 Importantly, LTN can penetrate the blood–brain barrier, allowing it to exert its effects directly within the brain. 16 According to the literature, LTN is able to effectively reduce pro-inflammatory mediators, enhance short-term memory, and facilitate neural plasticity in animal models with cognitive deficits resulting from chronic cerebral perfusion decline. 17 LTN exerts anxiolytic and antidepressant effects through multiple pathways and can ameliorate anxio-depressive-like behaviors by modulating neuroinflammation and enhancing hippocampal neurogenesis. 18 –20 Furthermore, LTN exhibits its anxiolytic and antidepressant effects by regulating the nuclear factor kappa B (NF-κB)/NLRP3 inflammasome axis and modulating cytokine release, especially in the context of sleep disorders. 21 These findings underscore its anti-inflammatory mechanisms, which contribute to its therapeutic potential in treating anxiety and depression.
The aim of this research was to investigate the potential of LTN in alleviating anxio-depressive symptoms in a rat model of PD. This study is novel in its approach, as it not only evaluates the anxiolytic and antidepressant effects of LTN but also explores its underlying mechanisms related to OS and inflammation—factors that are critically implicated in the pathophysiology of PD. To achieve this, the animals were initially subjected to intracerebral (IC) administration of 6-OHDA into their substantia nigra pars compacta (SNpc), which is a key region affected in PD. Subsequently, the rats were divided into different groups and administered either LTN or fluoxetine (FLX). Afterward, behavioral tests were performed to assess their anxio-depressive-like symptoms. In addition, the activation of OS markers and inflammatory factors was assessed in the hippocampus (HC) region of the animals. Molecular docking was also done for finding the affinity of LTN to inflammatory factors. By elucidating these mechanisms, our research paves the way for future investigations into LTN as a viable treatment option for managing psychological symptoms associated with PD, thus addressing a critical gap in current therapeutic strategies.
Materials and Methods
Study design
For this study, a total of 66 male Wistar rats weighing 250 ± 30 g were prepared from Beijing Vital River Laboratory Animal Technology Co. (Catalog number: A12200_1). The rats were housed under standard environment, with a controlled temperature of 23°C ± 1°C and a 12-hour light/dark cycle starting at 08:00 hours with unrestricted access to food and water. The study followed a set of experimental procedures based on the guidelines of the “NIH Guide for the Care and Use of Laboratory Animals.” Prior to the commencement of the study, the research protocol underwent thorough review and received approval from the Committee on the Ethics of Animal Experiments at Affiliated Taian City Central Hospital of
To prepare the LTN solution, it was dissolved in sterile saline solution, which served as the vehicle for LTN. The dosages of LTN were selected based on previous studies. 22 The 6-OHDA neurotoxin (6 μg/μL, Sigma-Aldrich) was prepared by dissolving it in a sterile saline solution with the addition of 0.2% ascorbic acid, which served as the vehicle for 6-OHDA. Following the stereotaxic surgery, a single dose of the neurotoxin (6 μg/μL) was infused bilaterally into the SNpc, following the methods described in the literature. 23
Neurosurgery
In this study, the researchers aimed to induce a lesion in SNpc. To achieve this, the rats were given xylazine and ketamine (5 mg/kg + 50 mg/kg, i.p.) for deep anesthesia. The skull of each animal was firmly fixed in a stereotaxic frame (Stoelting, Wood Dale, Illinois). 6-OHD ((6 μg/μL in 1 μL) was infused bilaterally by a Hamilton syringe (5 μL, Hamilton Co.) connected to 26-gauge steel needle into the SNpc. The injection coordinates for the neurotoxin administration were carefully determined: the anterior–posterior coordinate was set at −5.0 mm from the Bregma, the medial–lateral coordinate was set at ±2.3 mm from the midline, and the dorsal–ventral coordinate was set at −8.1 mm from the skull. 24 Moreover, sham operations were performed following the same procedure, but instead of the neurotoxin, a vehicle solution containing 0.9% sterile saline was injected.
Behavioral evaluation
To evaluate and analyze behavioral changes, we employed a variety of behavioral tests, including the tail suspension test (TST), sucrose preference test (SPT), forced swim test (FST), and open-field test (OFT). Before initiating any experimental procedures, the animals were provided with a minimum of 1 hour to adapt and acclimate to the testing room environment. This acclimation period allowed the animals to become familiar with their surroundings, reducing potential stress or anxiety that could influence the experimental results. The experimental timeline, outlining the duration of the experimental procedures, is visually represented in Figure 1.
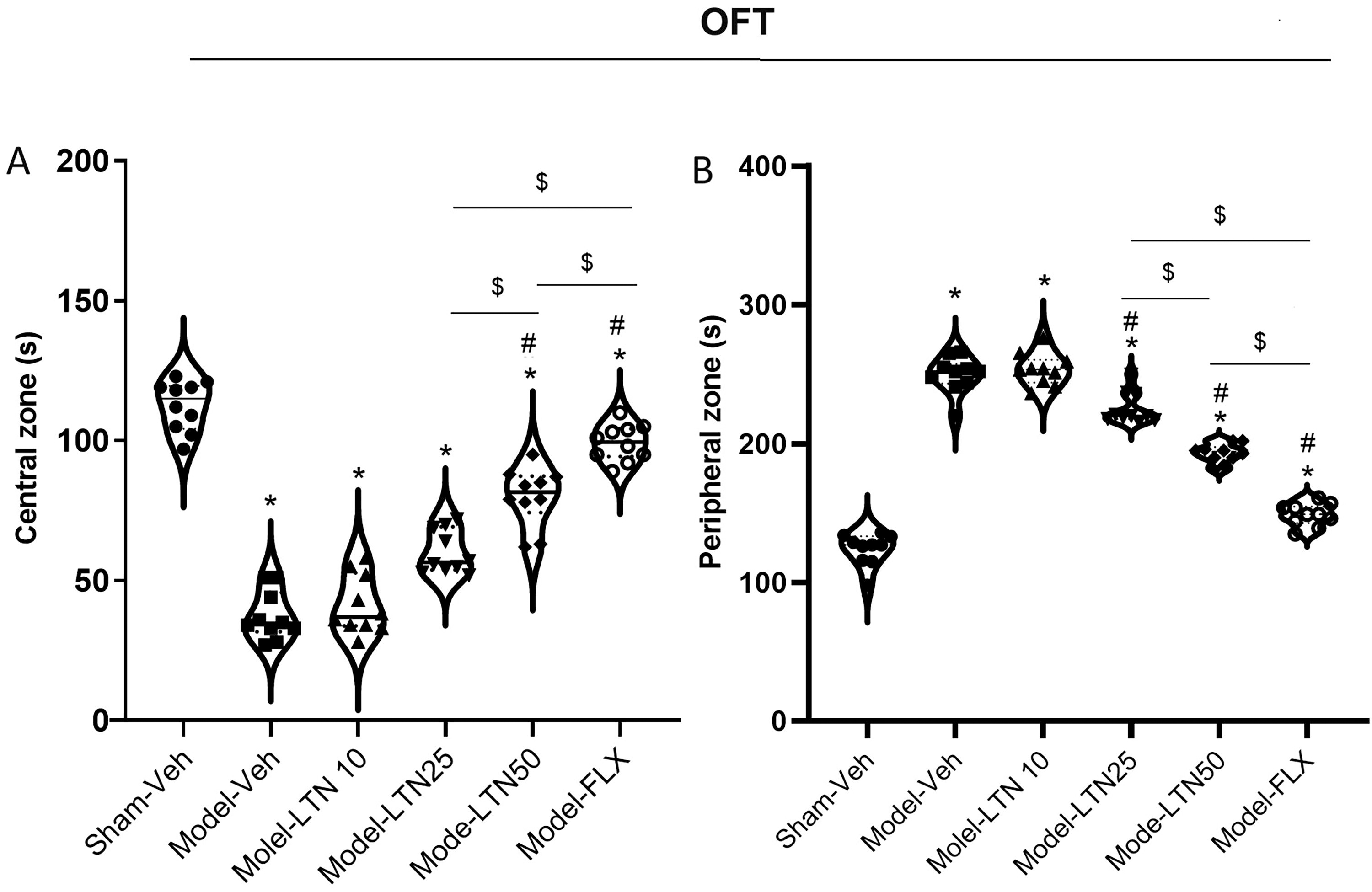
Impacts of LTN on anxiety-like behavior (OF) in the PD rat model. The OF test shows the activity time in:
OF test
The OF test is commonly utilized to evaluate anxiety levels in rodents. 25 The experimental setup consists of a rectangular arena (100 × 100 × 50 cm3). The floor of the enclosure is white and is divided into 16 individual squares. These squares include a central zone and peripheral zones. In this study, the activities of the animals were carefully observed and captured through video recordings. The main phase of test was conducted, and each animal was gently placed in the central zone of the testing arena and given a period of 5 minutes to freely explore and become acquainted with their surroundings. Throughout the test period, two specific parameters were measured: the amount of time spent by each animal in either the peripheral or central zones. By comparing these parameters among different groups, researchers were able to evaluate the anxiety levels exhibited by the animals.
FST
The FST is a commonly employed experimental method to assess depressive symptoms in animals. 26 The procedure entails placing the animal in a cylindrical container filled with water and measuring its behavioral. In this study, test was done in two separate sessions of training and test. During the training session, each animal was placed in a cylindrical tank with a diameter of 30 cm and a height of 80 cm. The tank was filled with water at a temperature of 22°C ± 1°C. The rats were forced to swim continuously for 15 minutes. This session served to familiarize the animals with the task and the water environment. Within 24 hours of the training session, a preliminary test was conducted. The test lasted for 6 minutes, during which the behaviors of the animals were recorded. During the final 5 minutes of the 6-minute test period, the researchers measured two parameters: (1) immobility time: refers to the period in which there is a lack of movement in the entire body, except for minimal motions essential to maintain the head of animal above the water surface, and (2) swimming time, refers to the period in which there are significant movements of the forepaws that generate water displacement and result in the body of animal moving within the cylinder, exceeding what is required for simply keeping the head of animal above the water surfaced.
TST
Following the FST, the TST was employed as a means of evaluating depressive symptoms. 27 In the TST, each animal was suspended by the tip of its tail, which was affixed to a flat surface using medical adhesive tape. The suspension height was set at 55 cm above the floor, and each rat remained suspended for a duration of 6 minutes. During the test, the duration of immobility was precisely measured for a specific time frame of 4 minutes, starting from the second minute and extending until the sixth minute. Immobility time refers to the duration during which the animal exhibited no movement or agitation.
SPT
The SPT is commonly employed as an indicator of anhedonia, a crucial symptom observed in rodents with depressive-like behavior. The SPT involves several steps to assess the animal’s preference for sucrose solution over water. 28 First, the animals were placed in individual housing and given free access to food during a 24-hour adaptation phase with a sucrose solution. Each rat had access to two similar preweighed bottles placed on opposite sides of their cages to ensure that they would drink from both bottles. After the adaptation phase, one of the bottles was randomly switched to contain a 1% sucrose solution, while the other bottle still contained water. This step was performed to introduce the choice between the two solutions. The rats were given 24 hours to drink from the bottles. To eliminate any potential side-preference effects, the positions of the bottles were reversed after the initial 24-hour period. After the completion of the experiment (24 hours later), the bottles were weighed to determine the amount consumed. The total intake was determined as the sum of sucrose consumption and water consumption. To calculate the sucrose intake as a percentage, the following equation was utilized: Sucrose preference (%) = (sucrose intake [in grams]/total intake [in g]) × 100. 29
Collection of brain samples
Once the behavioral assessments were completed, all rats were deeply anesthetized with a 1% pentobarbital sodium solution (60 mg/kg/i.p.). The isolated HC samples were stored at −80°C for further analysis. The collected samples served different purposes. The first set of tissues (n = 4 rats in each group) was used to assess OS biomarkers. The second set of tissues (n = 3 rats in each group) was utilized for the isolation of total RNA and subsequent quantitative PCR (qPCR) analysis. The third set of tissue samples (n = 4 rats in each group) were utilized to determine protein contents using the enzyme-linked immunosorbent assay (ELISA) technique.
Biomarkers of OS
To assess the OS status, the levels of antioxidant enzyme, catalase (CAT), superoxide dismutase (SOD), lipid peroxidation biomarker, and malondialdehyde (MDA) were assessed in the HC region of the animals. The HC samples were prepared for analysis by homogenizing them with the Polytron Homogenizer (IKA R T10). Following homogenization, the samples were subjected to centrifugation at 7200 ×g for 10 minutes at 4°C. This process resulted in the separation of a supernatant, which was then used for the assessment of OS biomarkers.
Measurement of SOD activity
The SOD activity was assessed by employing a SOD enzyme solution (Sigma-Aldrich), which measured the levels of protein capable of catalyzing the decomposition of oxygen radical (1 μM) into oxygen and hydrogen peroxide within a span of 1 minute. The quantification of SOD activity was achieved through a colorimetric measurement at a wavelength of 405 nm, utilizing a spectrophotometer. 30
Measurement of CAT activity
The evaluation of CAT activity involved the utilization of a CAT enzyme solution (Sigma-Aldrich). The assessment was conducted by quantifying the quantity of protein that facilitates the breakdown of 1 µM of hydrogen peroxide (H2O2) into water (H2O) and oxygen (O2) within a span of 1 minute. The measurement of this activity was carried out using spectrophotometry at a wavelength of 412 nm. 31
Measurement of MDA levels
The quantification of MDA content in the brain was performed using enzymatic assay kits obtained (ZellBio GmbH, Ulm, Germany), based on the manufacturer guidelines. The assessment of MDA levels was based on the reaction of the samples with thiobarbituric acid at an elevated temperature range of 90°C–100°C. The spectrophotometric measurement of this reaction was conducted at 535 nm, at room temperature, utilizing a spectrophotometer (Eon, Bio Tek). 32
qPCR
To quantify gene transcript levels of genes related to both pro-inflammatory and anti-inflammatory factors in the HC, qPCR was conducted. The samples were centrifuged at a (1000 ×g) for a duration of 15 minutes. Subsequently, the supernatants were then frozen at a temperature of −20°C to preserve the integrity of the RNA. The samples were subjected to the isolation of total RNA, from which 1 μg of RNA was employed for cDNA production by the reverse transcription kit (Takara Bio.). The resultant cDNA was analyzed using a LightCycler® 2.0 Instrument (Roche) with QuantiNova SYBR Green PCR Kit (QIAGEN). To calculate the relative expression levels of the target genes, the 2^(−ΔΔCt) method was employed. This calculation method involves comparing the threshold cycle (Ct) values of the target genes to reference genes and calculating the fold change in gene expression. In this study, the housekeeping gene was glyceraldehyde-3-phosphate dehydrogenase. The Ct values indicate the number of amplification cycles required for the fluorescent signal to surpass a predetermined threshold.
ELISA
ELISA was conducted to assess the protein concentrations of anti-inflammatory marker IL-10 and pro-inflammatory markers IL-6, IL-1β, and TNF-α in the HC. The ELISA kits, including rat IL-1β, IL-10, IL-6, and TNF-α kits (MyBioSource Inc.), were employed. The measurements were performed in line with the protocols provided by the manufacturer. The measurements were performed using the Varioskan Flash microplate reader (Thermo Scientific). This instrument operated at a wavelength of 450 nm, which is the commonly used detection wavelength for ELISA.
Molecular docking verification
To show the affinity of LTN to the components of NF-κB/NLRP3 axis, including NF-κB, NLRP3, ASC, and Casp1, we utilized docking analysis. The structure files of target proteins were obtained from the Protein Data Bank website (https://www.rcsb.org/). Then the chemical structure of LTN was obtained with the suffix name “sdf” from PubChem database (https://pubchem.ncbi.nlm.nih.gov/). 33 CB-Dock2 website (https://cadd.labshare.cn/cb-dock2/php/index.php/) was used to visualize the interaction between the LTN and target proteins utilized. The style of the protein was set as “cartoon,” and ligand was set as “spacefill.” The receptor color was based on “hydrophpbosity,” and the ligand color was based on the “element.” 34
Statistical analysis
The findings of study were demonstrated as the mean ± SEM. GraphPad Prism software (Ver. 6) was utilized for statistical analyses of both the behavioral test and molecular data. One-way ANOVA with Tukey’s post hoc test were utilized to compare data between groups. A significance level of p < 0.05 was applied for all tests.
Results
Behavioral changes
The OFT revealed significant differences in the delay experienced in both the central (F (5, 54) = 119.8; p < 0.0001) and peripheral (F (5, 54) = 252.9; p < 0.0001) zones among various groups. The results indicated that the Model-Veh (both: p < 0.0001), Model-LTN10 (both: p < 0.0001), Model-LTN25 (both zones: p < 0.0001), Model-LTN50 (both zones: p < 0.0001), and Model-FLX (p < 0.01 and p < 0.0001 for each zone) experimental groups exhibited a substantial reduction in time of activity in the central zone, accompanied by a simultaneous elevation in the time of activity in the peripheral zone, as compared with the Sham-Veh animals (Fig. 1A). Furthermore, the administration of both LTN and FLX significantly improved the movement duration in both the central and peripheral zones in the Model-LTN25 (p < 0.001 and p < 0.0001 for each zone), Model-LTN50 (both zones: p < 0.0001), and Model-FLX (both zones: p < 0.0001) compared with the Model-Veh animals. Notably, the results obtained from animals receiving a dosage of 50 mg/kg were comparable with those observed in the Model-FLX group, indicating a similar improvement in the time of activity within both the central and peripheral zones. Based on the OFT findings, the exposure to PD induced anxiety-like behaviors in the animals; however, the administration of LTN (25 and 50 mg/kg) effectively alleviated anxiety in these animals.
The results from the FST demonstrated significant differences in swimming time (F (5, 54) = 52.66; p < 0.0001) and immobility time (F (5, 54) = 62.64; p < 0.05) among the different experimental groups. Figure 2A illustrated a statistically significant reduction in swimming duration and a concomitant increase in immobility duration for the Model-Veh (both: p < 0.0001), Model-LTN10 (both: p < 0.0001), Model-LTN25 (both: p < 0.0001), Model-LTN50 (both: p < 0.0001), and Model-FLX (p < 0.05 and p < 0.001 for each parameter) animals in comparison with those of the Sham-Veh group. Furthermore, the administration of LTN and FLX effectively improved the behavioral alterations in swimming time and immobility duration in the Model-LTN25 (p < 0.05 and p < 0.0001 for each parameter), Model-LTN50 (both: p < 0.0001), and Model-FLX (both: p < 0.0001) animals when compared to those observed in the Model-Veh group. Notably, the outcomes observed in animals receiving a dosage of 50 mg/kg LTN were compared with those witnessed in the Model-FLX group. Based on the findings from the FST, the exposure to PD induced depressive symptoms in the animals; however, the administration of LTN (25 and 50 mg/kg) effectively alleviated these changes in animals with PD.

Impacts of LTN on depressive-like behavior (FST) in the PD rat model. The FST shows
The findings of the TST demonstrated a variation in the duration of immobility among the experimental groups (F (5, 60) = 104.3; p < 0.0001). Analysis of Figure 3A illustrated an elevation in immobility period of time for the Model-Veh, Model-LTN10, Model-LTN25, Model-LTN50 (all: p < 0.0001), and Model-FLX (p < 0.05) animals compared to the Sham-Veh animals. In comparison to the Model-Veh group, the administration of LTN and FLX effectively decreased immobility duration in the TST results of the Model-LTN25 (p < 0.001), Model-LTN50 (p < 0.0001), and Model-FLX (p < 0.0001) groups. LTN (25 and 50 mg/kg) effectively mitigated these behavioral alterations. Moreover, treatment with 50 mg/kg LTN resulted in a significant improvement in immobility during the TST, comparable with that observed with FLX.

Impacts of LTN on depressive-like behavior (TST and SPT) in the PD rat model. The immobility time in
SPT showed revealed a significant impact of PD on anhedonia in various experimental groups (F (5, 54) = 79.65; p < 0.0001). Figure 3B revealed that animals in the Model-Veh, Model-LTN10, Model-LTN25, Model-LTN50, and Model-FLX groups exhibited a decrease in preference for sucrose solution drinks compared with the Sham-Veh animals (p < 0.0001). Importantly, administration of LTN and FLX treatment led to a substantial improvement in the sucrose intake of the Model-LTN25 (p < 0.05), Model-LTN50 (p < 0.0001), and Model-FLX (p < 0.0001) animals compared to the Model-Veh animals. LTN (25 and 50 mg/kg) effectively mitigated these depressive symptoms in PD animals. Notably, the results obtained with a dosage of 50 mg/kg LTN exhibited a comparable improvement to that seen with FLX in terms of the SPT.
Levels of OS biomarkers
The levels of hippocampal CAT (F (5, 18) = 90.52), SOD (F (5, 18) = 94.37), and (F (5, 18) = 95.56) were affected by PD in different groups (Fig. 4A–C, all: p < 0.0001). Figure 4A showed that exposure to PD resulted in a reduction in the concentration of CAT and SOD and an elevation in the activity of MDA in the Model-Veh (all: p < 0.0001), Model-LTN10 (all: p < 0.0001), Model-LTN25 (all: p < 0.0001), Model-LTN50 (all: p < 0.0001), and Model-FLX (p < 0.01, p < 0.001, and p < 0.05 for each both marker) animals compared to the Sham-Veh animals. When comparing the Model-Veh animals to those treated with LTN and FLX, it was found that CAT and SOD activities were significantly increased in the Model-LTN25 (p < 0.01 and p < 0.001 for each parameter), Model-LTN50 (both: p < 0.0001), and Model-FLX (both: p < 0.0001) animals. However, treatment with LTN and FLX resulted in lower levels of MDA in the Model-LTN25 (p < 0.001), Model-LTN50 (p < 0.0001), and Model-FLX (p < 0.0001) animals compared to the Model-Veh animals. LTN (25 and 50 mg/kg) effectively reduced OS in PD animals, as confirmed by the improvement in concentrations of CAT, SOD, and MDA. Notably, the results of 50 mg/kg LTN showed comparable improvement to FLX in terms of levels of OS.

Impacts of LTN on the hippocampal concentrations of oxidative stress biomarkers in the PD rat model.
Neuroinflammatory biomarkers
The gene expression of hippocampal NF-κB (F (5, 12) = 92.82), NLRP3 (F (5, 12) = 246.7), the adapter apoptosis-associated speck-like (ASC) (F (5, 12) = 113.9), and Casp1 (F (5, 12) = 183.3) was altered in the PD-induced animals. A significant upregulation of NF-κB (Fig. 5A), NLRP3 (Fig. 5B), ASC (Fig. 5C), and Casp1 (Fig. 5D), in the Model-Veh, Model-LTN10, Model-LTN25, Model-LTN50 (all: p < 0.0001), and Model-FLX (p < 0.001, p < 0.001, p < 0.05, and p < 0.01, respectively) was seen compared with the Sham-Veh group. LTN (25 and 50 mg/kg) or FLX effectively reversed the alterations in gene expression of hippocampal NF-κB, NLRP3, ASC, and Casp1 in these groups compared with the Model-Veh group.
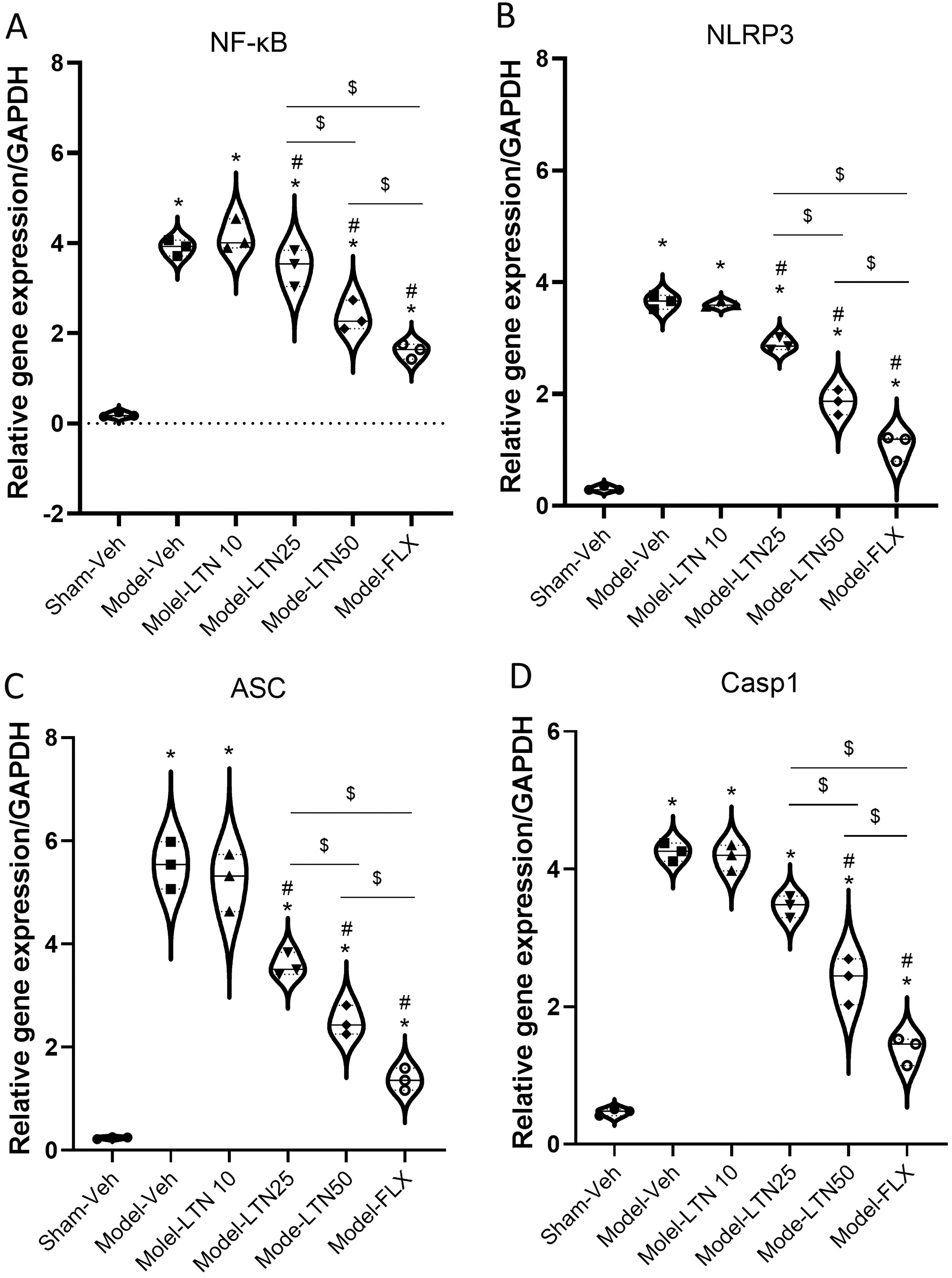
Impacts of LTN on the activity of NF-κB/NLRP3 inflammasome in the HC of the PD rat model. The gene expression of
Furthermore, the gene expression of pro-inflammatory mediators TNF-α (F (5, 12) = 145.1), IL-1β (F (5, 12) = 82.93), and IL-6 (F (5, 12) = 68.92) exhibited significant variations among the experimental groups (all: p < 0.0001; Fig. 6A–D). Furthermore, an upregulation of TNF-α was observed in the Model-Veh, Model-LTN10, Model-LTN25, Model-LTN50 (all: p < 0.0001), and Model-FLX (p < 0.05) animals in comparison with those of the Sham-Veh animals. Compared with the Sham-Veh group, the exposure to PD led to elevated levels of IL-1β in the Model-Veh, Model-LTN10, Model-LTN25 (all: p < 0.0001), and Model-LTN50 (p < 0.01) animals. Similarly, a significant upregulation of IL-6 in the Model-Veh (p < 0.0001), Model-LTN10 (p < 0.0001), Model-LTN25 (p < 0.001), and Model-LTN50 (p < 0.001) animals compared to the Sham-Veh animals. Administration of LTN (25 and 50 mg/kg) or FLX effectively reversed the alterations in gene expression of TNF-α, IL-1β, and IL-6 in these animals compared to the Model-Veh group. Moreover, the results obtained from the 50 mg/kg dosage of LTN exhibited a similar level of improvement in neuroinflammation to that observed with FLX.

Impacts of LTN on the gene expression of inflammatory mediators in the HC of the PD rat model.
The gene expression levels of IL-10 (F (5, 12) = 38.41) in the HC and TNF-α/IL-10 (F (5, 12) = 44.21), IL-1β/IL-10 (F (5, 12) = 23.41), and IL-6/IL-10 (F (5, 12) = 64.32) to the gene expression levels IL-10 differed among groups of study (all: p < 0.0001; Fig. 6E–G). According to Figure 6E, a substantial downregulation in the IL-10 in the Model-Veh (p < 0.01) and Model-LTN10 (p < 0.05) animals and a significant upregulation of this gene in the Model-LTN50 (p < 0.05) and Model-FLX (p < 0.001) animals were recorded compared with the Sham-Veh animals. In addition, significant upregulation of IL-10 was seen in the HC of Model-LTN25 (p < 0.01), Model-LTN50 (p < 0.001), and Model-FLX (p < 0.05) animals compared with those Model-Veh group. Furthermore, significant increases in the TNF-α/IL-10, IL-1β/IL-10, and IL-6/IL-10 were seen in the Model-Veh (all: p < 0.0001) and Model-LTN10 (p < 0.001, p < 0.001, and p < 0.0001 for each parameter) animals compared with the Sham-Veh animals. Reduced ratio of TNF-α/IL-10 was seen in Model-LTN10 (p < 0.01 and p < 0.01 for each parameter), Model-LTN25 (p < 0.0001, p < 0.001, and p < 0.0001 for each parameter), Model-LTN50 (p < 0.0001, p < 0.001, and p < 0.0001 for each parameter), and Model-FLX (all: p < 0.0001) animals compared with the Model-Veh animals. According to the results, the administration of 25 and 50 mg/kg of LTN and FLX showed promising effects on IL-10 levels and the ratios of pro-inflammatory cytokines to IL-10 in animals with PD-induced symptoms. Specifically, both LTN and FLX were found to increase the levels of IL-10 in the HC while reducing the ratios of IL-1β/IL-10, IL-6/IL-10, and TNF-α/IL-10. Notably, the dose of 50 mg/kg of LTN demonstrated comparable effects to FLX.
The protein concentrations of hippocampal inflammatory cytokines, including TNF-α (F (5, 18) = 279.0; p < 0.0001), IL-1β (F (5, 18) = 190.2; p < 0.0001), and IL-6 (F (5, 18) = 164.8; p < 0.0001), and IL-10 (F (5, 18) = 227.6; p < 0.0001), exhibited significant variations among the experimental groups (Fig. 7A–D). The results indicated a significant upregulation of TNF-α, IL-1β, and IL-6 in the Model-Veh (all: p < 0.0001), Model-LTN10 (all: p < 0.0001), Model-LTN25 (all: p < 0.0001), Model-LTN50 (all: p < 0.0001), and Model-FLX (all: p < 0.05) animals compared with the Sham-Veh animals (Fig. 7A–C). Figure 7D demonstrates a statistically significant reduction in the protein levels of IL-10 in the Model-Veh (p < 0.0001) and Model-LTN10 (p < 0.0001) animals, while the Model-LTN50 (p < 0.001) and Model-FLX (p < 0.0001) animals exhibited a raise in the IL-10 protein concentrations in comparison with those of the Sham-Veh animals. LTN (25 and 50 mg/kg) or FLX effectively reversed the alterations in protein concentrations of IL-1β, IL-6, TNF-α, and IL-10 in these animal groups when compared with the Model-Veh group. In addition, the results obtained from the 50 mg/kg dosage of LTN demonstrated a comparable level of improvement in neuroinflammation to that observed with FLX.
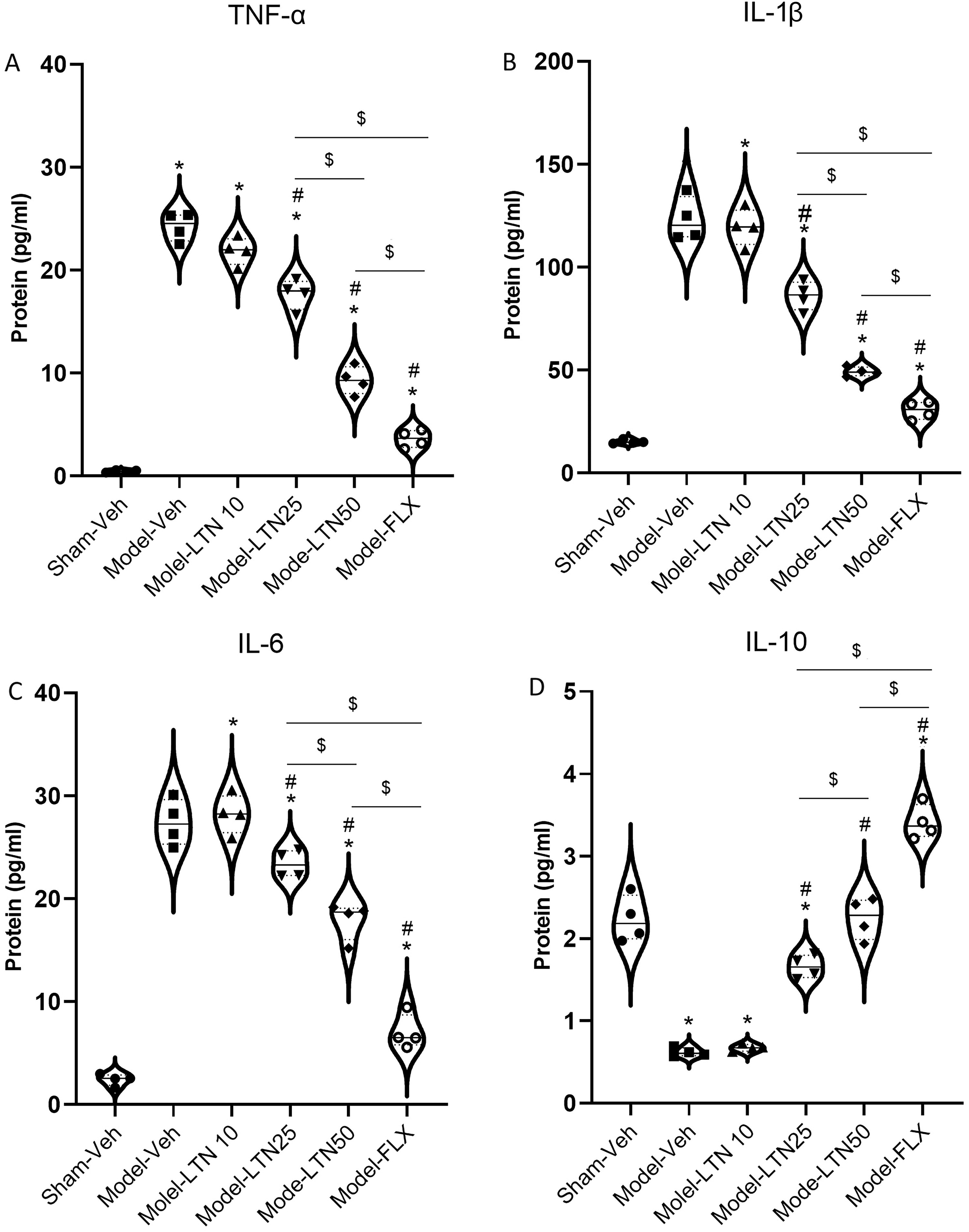
Impacts of LTN on the protein concentrations of inflammatory mediators in the HC of the PD rat model.
Molecular docking
The analysis with CB-Dock2 involved examining the affinity between LTN and four proteins, including NF-κB, NLRP3, ASC, and Casp1 via molecular docking. These interactions were identified through topological analysis. In this part of the study, we identified the most stable binding configuration between the ligand and proteins which can be shown by more negative vina scores. 35 The most stable bindings were between the LTN and core protein targets of SC in the context of NP/DD treatment, we specifically selected vina scores that were negative and less than −5.9. 36 Based on the docking scores presented in Figure 8, the combinations of LTN and NF-κB (vina score = −8.9), NLRP3 (vina score = −8.2), ASC (vina score = −6.7), and Casp1 (vina score = −6.8) exhibited the strongest affinity.

Molecular docking between LTN and targets. LTN, luteolin.
Discussion
In this research, we aimed to find the potential anxiolytic and antidepressant effects of LTN in PD animals. The findings of various behavioral tests, including OF, FST, TST, and SPT, confirmed that the PD-induced symptoms in our rat models successfully mimicked anxiety- and depressive-like behaviors. The I.C. administration of 6-OHDA resulted in elevated time of activity in the peripheral zone and reduced in time of activity in the central region of the OF apparatus, which further validated the existence of anxiety-like symptoms in the animals. In addition, the PD animals exhibited raised immobility durations in the TST and FST tests, along with a decrease in sucrose preference, indicating the existence of depressive behaviors. Moreover, the induction of PD also led to increased hippocampal OS, as evidenced by elevated concentrations of MDA and reduced activity of CAT and SOD. This suggests that PD triggers oxidative damage in the HC. Besides, we found raised neuroinflammatory responses in the HC of PD animals, as indicated by the upregulation of NF-κB, IL-1β, TNF-α, and IL-6, as well as elevated protein concentrations of IL-1β, TNF-α, and IL-6. Interestingly, we also noted a downregulation of IL-10, indicating a decreased anti-inflammatory response in PD animals. Our study revealed an increase in the ratios of gene expression TNF-α/IL-10, IL-1β/IL-10, and IL-6/IL-10 in the HC, further indicating the presence of neuroinflammation in PD animals. These ratios serve as markers of the pro-inflammatory state, as they reflect the imbalance between pro-inflammatory factors (IL-1β, IL-6, and TNF-α) and the anti-inflammatory factor (IL-10) in the HC.
PD is a progressive neurological disorder accompanied by nonmotor impairments of varying severity. 37 Nonmotor manifestations, including depression and anxiety, are commonly observed in individuals with PD and significantly impact their quality of life. 38 The etiology of anxio-depressive disorders in PD is complex and influenced by multiple factors. Anxio-depressive symptoms that occur in PD may result from a reactive response to the psychosocial stress caused by the chronic nature of the disease and the resulting disability. Alternatively, it is proposed that depression in PD could be attributed to structural alterations in the brain due to the process of neurodegeneration. 39 Brain imaging studies can be used for the detection of relationship between changes in neuropsychiatric function and structural alterations within the brain. Magnetic resonance imaging studies of PD individuals with depressive disorders showed a decrease in the cortical thinning of anterior cingulate, left temporal, right hippocampal, and posterior cingulate cortices. 40,41 Furthermore, individuals diagnosed with PD who also experience depression exhibit reduced densities of 5-HT1A receptors in limbic areas such as the insula, HC, and frontal cortex. 42 The interplay between a reduction in dopamine concentrations and alterations in the posttranslational modification of α-Syn protein has a detrimental synergistic effect, leading to the impairment of neurogenesis in the adult HC. 43 The impairment of adult neurogenesis and the occurrence of hippocampal atrophy have been observed in both postmortem investigations on the brain samples from PD individuals and genetic models. 44
While changes in the serotonin (5-HT) system and central dopaminergic deprivation related to motor manifestations in PD individuals may play an essential role in the progression of depression, there is a scarcity of research specifically focusing on the models of PD and depression. 45 Recent research has demonstrated that PD models induced using 6-OHDA can replicate anxio-depressive-like behaviors as comorbidities of PD. 38,46,47 Multiple mechanisms have been recognized as contributors to the comorbidity of anxiety and depression in animal models of 6-OHDA-induced PD. Notably, a study by Vieira et al. found that the 6-OHDA-induced parkinsonism model in rats elicited anxiety-like behaviors, which was correlated with an imbalance in neurotransmitter systems in the regions associated with anxiety, including the amygdala, striatum, and prefrontal cortex. 38
Our study findings indicate that OS and neuroinflammation are significant contributors to the pathophysiology of anxio-depressive-like behaviors that emerge following PD. The existing literature has extensively explored the role of OS in both depression and PD. In the context of depression, it is suggested that heightened OS, coupled with immune responses mediated by the hypothalamic–pituitary–adrenal axis, leads to increased levels of inflammatory mediators that activate apoptotic pathways, ultimately resulting in neuronal death. 48,49 In PD, OS is associated with the degeneration of dopaminergic neurons in the SN. 50 The contribution of neuroinflammatory responses to the development of depression related to PD remains a subject of ongoing investigation. It has been hypothesized that OS, stemming from mitochondrial dysfunction, triggers an inflammatory cascade that disrupts the functionality of glial cells, specifically astrocytes and microglia. This disruption is believed to play a crucial role in neurodegeneration, which is a key factor in the onset of depressive symptoms. 51,52 Furthermore, dysregulation of pro-inflammatory cytokines such as TNF-α, interferon-gamma (IFN-γ), IL-1β, IL-2, and IL-6 in the striatum has been linked to anxio-depressive-like behaviors observed in the 6-OHDA model of PD. 53 Additionally, stress, in combination with 6-OHDA administration, has been shown to induce OS through an increase in nitrite/nitrate concentrations and dysregulation of SOD and MDA in various brain regions. 54 In fact, OS, marked by an elevation in reactive oxygen species and a reduction in the antioxidant capacity, is intricately intertwined with the amplification of inflammatory responses. Substantial evidence revealed that alterations in inflammatory mediators such as IL-6 and TNF-α in individuals afflicted with depressive disorder, play a contributory role in the deterioration of symptoms seen in PD. 55,56 Moreover, substantial increases in microglial activation and the expression of the NLRP3 inflammasome have been identified in the SN tissues of patients with PD. 57 Research indicates that various neurotoxins, which induce PD-like symptoms in both cellular and animal models—such as 6-OHDA, 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine, 1-methyl-4-phenylpyridinium, and paraquat—can activate the NLRP3 inflammasome, as demonstrated in numerous recent studies. 58 The complex interplay between OS, inflammation, and neurodegeneration highlights the intricate pathophysiological mechanisms underlying both PD and depressive disorders. This complexity underscores the need for further research to elucidate their interactions and identify potential therapeutic targets. Consequently, strategies focused on mitigating OS and neuroinflammation may offer effective approaches for managing anxiety and depression in individuals with PD.
Over the past few years, there has been increasing academic interest in the potential antidepressant effects of natural active components found in herbs. 59,60 These compounds have gained attention due to their multitarget potential, meaning they can act on multiple pathways in the brain associated with anxiety and depression. 60,61 In our study, we investigated the potential of LTN (at doses of 10, 25, and 50 mg/kg) in managing anxio-depressive symptoms in a rat model of PD. The results of the behavioral tests, including the OF, FST, TST, and SPT, support the potential of LTN (at doses of 25 and 50 mg/kg) as a therapeutic approach for alleviating anxiety and depressive symptoms in PD. The results of the OF test showed a decrease in the time duration spent in the peripheral zone and an increase in the time duration spent in the central zone, suggesting that LTN (at doses of 25 and 50 mg/kg) may possess anxiolytic properties in the PD-induced rat models. The FST and TST revealed a reduction in immobility duration following LTN administration at both doses, indicating an improvement in depressive-like symptoms. The SPT also demonstrated an increase in sucrose preference in the LTN-treated animals, suggesting an improvement in anhedonia. Notably, the efficacy of LTN (at a dose of 50 mg/kg) was compared with that of FLX, a known strong antidepressant, highlighting the potential of LTN as an effective alternative or adjunctive treatment for depression.
The literature indicates that LTN exhibits notable anxiolytic and antidepressant effects through the regulation of multiple biological pathways. Gadotti and Zamponi confirmed LTN’s anxiolytic properties in a mouse model of acute dextran sodium sulfate-induced colitis, as demonstrated by behavioral tests including the novelty suppressed feeding test, OF test, and elevated plus maze test. 62 Zhou et al. conducted a review on LTN’s antidepressant effects, linking these to various mechanisms such as reducing OS, preventing apoptosis, alleviating endoplasmic reticulum (ER) stress, and mitigating neuroinflammation. They identified critical targets that LTN interacts with, including AKT serine/threonine kinase 1, insulin-like growth factor 1 receptor, estrogen receptor alpha, epidermal growth factor receptor, and prostaglandin-endoperoxide synthase 2. This suggests that LTN may positively influence depression by acting on several targets and pathways. 63 Moreover, Zhu et al. reported that the antidepressant-like effects of LTN were associated with both direct and indirect regulation of 5-HT reuptake, achieved through the inhibition of the plasma membrane monoamine transporter. 64 In addition, LTN has been shown to exhibit antidepressant properties by inhibiting ER stress. This is evidenced by its ability to reduce Casp3 cleavage, a marker of apoptosis, while enhancing the expression levels of glucose-regulated protein 78 (GRP78) and GRP94, which are molecular chaperones predominantly found in the ER. 65 Furthermore, LTN may alleviate depressive-like symptoms by modulating glycerophospholipid metabolism in the prefrontal cortex and HC of rat models with late-onset depression. 66 This multifaceted approach highlights LTN’s potential as a therapeutic agent in managing anxiety and depression, emphasizing its role in various neurobiological processes.
In our study, LTN at doses of 25 and 50 mg/kg showed anxiolytic, antiparkinsonian, and antidepressant effects through the regulation of hippocampal OS and neuroinflammation. Intriguingly, LTN administration appeared to reduce OS in the HC by modulating lipid peroxidation (MDA) and the activity of CAT and SOD. This suggests that LTN may have antioxidant properties and can help alleviate OS in the HC. For the regulation of neuroinflammation, LTN successfully downregulated the gene expression of NF-κB/NLRP3 pathway components, including NF-κB, NLRP3, ACS, and Casp1 in the HC of PD animals. High affinity (more negative vina score) was recorded between LTN and NF-κB, NLRP3, ACS, and Casp1. In addition, LTN treatment led to the downregulation of gene expression and protein levels of pro-inflammatory mediators, including TNF-α, IL-1β, and IL-6. Interestingly, we also observed an elevation in levels of the anti-inflammatory cytokine IL-10, indicating an enhancement of the anti-inflammatory response in these animals. Moreover, the observed decrease in TNF-α/IL-10, IL-1β/IL-10, and IL-6/IL-10 ratios reflects an improvement in the imbalance between pro-inflammatory cytokines and the anti-inflammatory cytokine within the HC. Furthermore, LTN (50 mg/kg) exhibited comparable effects to FLX in the regulation of OS and inflammation, further supporting the potential of LTN as an effective alternative or adjunctive treatment for anxiety and depression.
LTN, a type of flavonoid, is present in numerous plants such as fruits, vegetables, and medicinal herbs. 15 Research has indicated that LTN possesses antioxidant, anti-inflammatory, and antitumor properties, positioning it as a promising therapeutic candidate for enhancing human health. 67 To comprehend the mechanisms of LTN’s action within the body, it is essential to explore its metabolic characteristics, bioavailability, and absorption, in living organisms. While the research is still limited, it is believed that LTN undergoes metabolic processes related to intestinal absorption after being ingested. 68 Although specific details on lead optimization for LTN in PD are not directly available from the provided sources, LTN has shown optimal microsomal stability with a half-life greater than 60 minutes per oral, indicating that it is metabolically stable and may have a prolonged duration of action in the body. 69 LTN is stable in blood plasma, which is crucial for maintaining its therapeutic levels in the bloodstream. 70 LTN has a kinetic solubility of 21 ± 4 µM in Phosphate Buffered saline (PBS) neutral buffer (pH 7.4), which is a measure of its solubility under physiological conditions. 71 These pharmacokinetic properties suggest that LTN could be a promising candidate for further development in PD treatment. The neuroprotective effect of LTN plays a crucial role in the treatment of PD. 72 Several studies have assessed the antioxidant and anti-inflammatory impacts of LTN, revealing that its antioxidant properties are primarily linked to the regulation of nuclear factor erythroid-2-related factor 2 (Nrf2). 15 Elmazoglu et al. found that LTN could protect BV2 microglial cells from toxicity induced by rotenone in a hormetic manner. This protection was achieved by targeting OS responses and genes associated with PD, as well as inflammatory pathways, which included reductions in TNF-α and IL-1β levels, LDH release (measured via ELISA), and the expression of genes such as PTEN-induced putative kinase 1 (Pink1), Parkin (Park2), Leucine-rich repeat kinase 2 (Lrrk2), Thioredoxin 1 (Trx1), and Nrf2. 73 Research involving primary microglial cells from rats and BV2 microglial cells demonstrated that LTN effectively reduced the production of nitric oxide and pro-inflammatory cytokines induced by lipopolysaccharides and IFN-γ. This action contributed to a decrease in OS levels. Furthermore, LTN plays a key role in inhibiting the signaling pathways of NF-κB, IFN regulatory factor 1, and signal transducer and activator of transcription 1, resulting in a diminished inflammatory response. 74
These findings underscore the potential therapeutic value of LTN in alleviating PD-related symptoms and in the regulation of OS and inflammatory pathways.
LTN has emerged as a promising compound with antidepressant effects, primarily by targeting autophagy biomarkers such as AMPK, the LC3II/LC3I ratio, and Beclin-1. In addition, LTN plays a crucial role in regulating neuroinflammatory pathways by modulating the NLRP3 inflammasome, which leads to the inhibition of inflammatory markers such as IL-1β and IL-18. 12 For instance, Mokhtari et al. demonstrated that LTN alleviates pain-induced anxiety and depressive-like symptoms through the regulation of molecular pathways in the HC and prefrontal cortex. LTN was also reported to combat OS by influencing markers such as MDA, SOD, CAT, and Nrf2. It also exhibited antiapoptotic effects by modulating Bcl-2 and Bax expressions. Moreover, LTN showed potential anti-inflammatory effects, as evidenced by its ability to regulate NF-κB, NLRP3, TNF-α, IL-18, IL-1β, and IL-6. The neuroprotective effects of LTN were associated with its regulation of GDNF and BDNF in the HC and prefrontal cortex. 19 In another study, Xiong et al. reported that LTN improves anxiety and depression in an animal model of sleep deprivation by modulating components of the NF-κB and NLRP3 inflammasome, including ASC, NLRP3, and Casp1 in the HC. 21 Collectively, these findings highlight LTN’s multifaceted role in promoting neuroprotection, reducing inflammation, and alleviating symptoms of anxiety and depression.
Conclusion
In conclusion, our study successfully induced PD in rat models, which manifested as anxiety- and depressive-like behaviors, evidenced by increased anxiety responses and depressive symptoms. These behavioral changes were correlated with heightened OS and neuroinflammation in the HC, underscoring the roles of these processes in PD-related psychiatric manifestations. Notably, the administration of LTN at doses of 25 and 50 mg/kg demonstrated significant promise in alleviating these anxiety and depressive disorders in PD-affected rats. LTN treatment resulted in a marked reduction of anxiety-like behaviors and less immobility during the TST and FST, indicating its potential as both an anxiolytic and antidepressant agent. In addition, LTN improved reward responsiveness, as evidenced by increased sucrose preference, suggesting its potential utility in counteracting anhedonia, a common symptom in PD. Moreover, LTN exhibited beneficial effects on hippocampal alterations associated with PD, showcasing antioxidant properties that reduced OS markers while enhancing the activity of antioxidant enzymes. LTN treatment also led to a decrease in neuroinflammation, as indicated by the suppression of NF-κB/NLRP3 inflammasome activity, downregulation of pro-inflammatory biomarkers, and an increase in the anti-inflammatory cytokine IL-10 in the HC. These findings strongly indicate that LTN could serve as a valuable therapeutic intervention for mitigating anxiety and depressive symptoms related to PD, primarily through its antioxidant and anti-inflammatory effects in the HC. Clinical studies are warranted to explore the application of LTN in human patients with PD, focusing on its efficacy, safety, and potential mechanisms of action. By investigating these aspects, we can better understand how LTN might improve the quality of life for patients suffering from the psychiatric symptoms associated with PD, ultimately guiding its integration into clinical practice.
Footnotes
Authors’ Contributions
R.S. and X.L.: Study conception and design. R.S., Z.Z., M.H., and D.Z.: Methodology, investigation, and data collection. J.Z., Y.C., and X.Z.: Analysis and interpretation of results. R.S., D.Z., and X.Z.: Draft article preparation. Y.C. and X.L.: Supervision, writing-review and editing, and funding acquisition. All authors reviewed the results and approved the final version of the article.
Availability of Data and Material
Data are however available from the authors upon reasonable request.
Ethics Approval
Animals had ad libitum access to standard food and water. The present research was approved by the Affiliated Taian City Central Hospital of
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
This study was funded by Affiliated Taian City Central Hospital of Qingdao University, Taian, China.