
Abstract
Aging is associated with a gradual decline in cellular function, largely driven by oxidative stress, which leads to cellular senescence. These processes contribute to tissue degeneration and age-related dysfunction. Human dermal fibroblasts (HDFs), critical for maintaining skin structure, are highly vulnerable to oxidative damage, making them key contributors to skin aging. Umbilical cord blood plasma (UCBP), rich in growth factors and regenerative molecules, has shown potential in preventing cellular senescence and addressing key mechanisms of tissue aging. Based on findings from heterochronic parabiosis experiments that demonstrated the rejuvenating effect of young blood, we investigated the effects of UCBP on hydrogen peroxide (H2O2) induced oxidative stress in HDFs and compared its efficacy with adult blood plasma (ABP). Our results indicate that although both UCBP and ABP reduce reactive oxygen species (ROS), UCBP is more effective in suppressing cellular senescence and maintaining fibroblast proliferation. These findings suggest that UCBP’s protective effects extend beyond ROS reduction, potentially by modulating the senescence-associated secretory phenotype and the enhancement of tissue repair mechanisms.
Introduction
Aging is a complex biological process characterized by a gradual loss of cellular function and the accumulation of molecular damage. 1,2 This progression leads to tissue dysfunction and an increased risk of age-related diseases. 3 A key factor in this process is oxidative stress, which is driven by the excessive production of reactive oxygen species (ROS), such as hydrogen peroxide (H2O2). 4,5 ROS cause damage to essential cellular components, including DNA, lipids, and proteins, which accelerate the onset of cellular senescence. 6 ROS-induced damage impairs cellular homeostasis and promotes the secretion of proinflammatory molecules, which further contribute to the aging process. 7
Human dermal fibroblasts (HDFs) are vital for maintaining skin structure and function but are particularly susceptible to oxidative stress. 8 Environmental factors, such as UV radiation and pollution, increase ROS levels in the skin, further promoting cellular senescence. In vitro models using H2O2 demonstrate that oxidative damage leads to premature senescence in HDFs, impairing their proliferative capacity and disrupting tissue homeostasis. 9 Moreover, senescent cells secrete a range of proinflammatory cytokines, chemokines, and proteases known collectively as the senescence-associated secretory phenotype (SASP), which exacerbates tissue degeneration. 3,10
Emerging evidence suggests that components in young blood can counteract the detrimental effects of aging. 11 In heterochronic parabiosis experiments, where the circulatory systems of young and old animals are connected, young blood has been shown to rejuvenate tissues in older animals by reducing inflammation, promoting repair, and potentially clearing senescent cells. 12 –16 Growth factors such as GDF11 and oxytocin, along with other factors present in young blood, have been implicated in these rejuvenation processes. 17,18
In light of these findings, umbilical cord blood plasma (UCBP) has gained attention as a promising therapeutic agent. 19 –24 Similar to young blood, UCBP is rich in growth factors, cytokines, and molecules that promote tissue repair and cellular regeneration. 19,23 UCBP contains high concentrations of vascular endothelial growth factor (VEGF) and insulin-like growth factor (IGF), both of which are known to enhance the skin’s defenses against oxidative stress and support tissue recovery. 22 Despite this potential, the effects of UCBP on oxidative stress-induced senescence in HDFs remain largely unexplored.
Given its regenerative properties, we hypothesized that UCBP would outperform adult blood plasma (ABP) in mitigating oxidative stress, reducing ROS levels, delaying senescence, and preserving fibroblast proliferation. Furthermore, we aimed to investigate whether UCBP’s protective effects extend beyond ROS scavenging, specifically in modulating the SASP and enhancing cellular repair mechanisms. By comparing the efficacy of UCBP and ABP, this study seeks to clarify the therapeutic potential of UCBP in promoting skin health and counteracting age-related tissue degeneration.
Materials and Methods
Blood processing and plasma isolation
This study was approved by the Institutional Review Board of MEDIPOST Co., Ltd. (P01-202312-02-015). The human umbilical cord blood (hUCB) units were provided to MEDIPOST Co., Ltd. for research purposes. The umbilical cord blood units were received within 24 hours after collection. All samples were tested for infectious diseases, including HIV, syphilis, hepatitis B and C using Bioline™ test kits (Abbott, Korea) and were negative.
To prepare UCBP, the collected cord blood was centrifuged at 6,000 g for 10 minutes to remove residual red blood cells, and the supernatant was frozen at −80°C. ABP was obtained as fresh-frozen plasma (FFP) from the Korean Red Cross for research purposes. Both UCBP and ABP samples were thawed and filtered using a 40 μm strainer (Corning®, USA, Cat #: CLS352340) to remove cryoprecipitates. The 40 μm pore size of the filter does not retain small proteins such as catalase, SOD, or VEGF. Filtered plasma samples were aliquoted and stored at −80°C until use. The human plasma samples were fully anonymized and could not be traced back to any personal information or linked to individual identities.
Cell culture
Normal HDFs (donor; adult, cat #: C-12302), were purchased from PromoCell GmbH, Germany. The cells were grown in high glucose (4.5 mg/mL) Dulbecco’s modified Eagle medium (DMEM) containing pyruvate (Gibco™, Thermo Fisher Scientific, USA, Cat #: 11995065) supplemented with 10% fetal bovine serum (FBS) (HyClone™ Characterized Fetal Bovine Serum, AU Origin, Cytiva™, Cat #: SV30207.02) and 1.0% penicillin streptomycin (Gibco™, Thermo Fisher Scientific, USA, Cat #: 15140122). Cells were seeded at an initial density of 4000 cells/cm2 and incubated at 37°C in a humidified atmosphere with 5% carbon dioxide (CO2). Cells were passed every 3–4 days, ensuring confluence did not exceed 80%. For maintaining young characteristics, the number of passages from 5 to 9 were used in experiments.
Cell treatment
Catalase inhibitor treatment
As a catalase inhibitor, we purchased 3-amino-1,2,4-triazol (3-AT) (Sigma Aldrich, USA, Cat #: A8056). According to the product Information, 3-AT was dissolved in distilled water and stored at −20°C until use. 25,26 Considering the concentration of 3-AT dose used in previous research, fibroblasts were treated with 0.25–25 mM 3-AT. Except for the 25 mM concentration, which exhibited cytotoxicity in HDFs, 3-AT was added to plasma-containing medium before and after H2O2 treatment.
Plasma treatment and induction of cellular senescence
HDFs were seeded at a density of 4000 cells/cm2 with DMEM supplemented with 10% FBS, after 24 hours cells were washed with serum-free DMEM once and added media containing 5% ABP or UCBP with 0.5 unit/mL heparin (Sigma-Aldrich, USA, Cat #: H3149). The heparin dose was selected based on its effects on HDF morphology, as reported in previous study. 27 To induce the cellular senescence state, the concentration of hydrogen peroxide (H2O2, Sigma-Aldrich, USA, Cat #: H3410) was determined to be 600 μM by trials with dose and time dependent treatment on HDFs following previous studies. 9,28 –33 To examine the potential effects of plasma, HDFs were pretreated with 5% plasma for 4 hours before exposure to H2O2, based on previous studies. 34 –37 Cells were induced to the state of senescence-phenotype by treatment with H2O2 for 20 hours, then it was washed with serum-free DMEM twice and supplemented with 5% plasma containing DMEM (0.5 U/mL of heparin). HDFs were incubated at 37°C, 5% CO2 for 3 or 7 days. Before the confluency of cells was over 80%, subculture was performed then media was exchanged to plasma containing DMEM.
To induce replicative senescence, HDFs were cultured in DMEM supplemented with 10% FBS under standard growth conditions until passage 20, at which point the population doubling time reached approximately 64 hours. Cells were seeded at a consistent density and on the following day, the culture medium was replaced with DMEM containing 5% plasma (0.5 U/mL of heparin) and maintained for three days.
Intracellular ROS measurement
To measure the quantitative level of reactive oxidative species (ROS), DCFDA/H2DCFDA—Cellular ROS Assay Kit was used (Abcam, UK, Cat #: ab113851). Following the protocol, cells were washed with 1× buffer without serum then incubated 20 μM of 2',7'-Dichlorofluorescin Diacetate (DCFDA) for 30 minutes at 37°C. After the reaction with DCFDA, cells were added with plasma containing medium before and after treatment of H2O2. Fluorescence intensity was measured using Microplate Reader (Perkin Elmer VICTOR Nivo, USA) with Ex/Em = 485/535 nm. After compensating the value by offsetting the observed background value, ROS level was presented as the fold change to normal cells untreated with H2O2.
Enzyme linked immunosorbent assay
The concentrations of human Catalase (Cusabio® Techonology LIc, Houston, TX, USA) in plasma were measured in accordance with the manufacturer’s protocols. All samples were assayed with technical triplicate. The optical density (OD) of sample wells was measured with a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Senescence-associated β-galactosidase staining
To observe the extent of senescence, as the representative marker of senescence, β-Galactosidase Staining Kit was used (Cell Signaling Technology, USA, Cat #: 9860). Cells were maintained for 7 days to confirm the noticeable senescence-induced β-galactosidase activity in a condition supplemented with plasma after treatment of H2O2. An equal number of cells were plated and incubated with staining solutions for 48 hours. The positive SA-β-gal cells (blue staining) were counted under a microscope more than three places and the experiments were conducted in triplicate.
RNA extraction and quantitative real-time PCR
Induced senescent and replicatively senescent HDFs were maintained in conditions supplemented with FBS, ABP or UCBP for 3 days. Total RNA was extracted using TRIzol (Invitrogen™, USA, Cat #: 15596018). Quantified RNA sample was reverse transcribed with SuperiorScript III RT Master Mix (RT300M, enzynomics) according to the manufacturer’s protocol. RT-PCR was performed with LightCycler® 480 SYBR Green I Master (04887352001, Roche) and primers described in Table 1. Samples were obtained with relative copy numbers and normalized to the value of housekeeping gene, beta-actin. The fold changes were calculated using the 2−ΔΔCT method.
The Primers Used in Our Experiment
Western blot analysis
The normal HDFs were washed with sterile phosphate buffered saline (PBS) and lysed using RIPA buffer (Pierce™ RIPA buffer, Thermo scientific, USA, Cat #: 8990) containing 1x protease inhibitor cocktails (Halt™ Protease and Phosphatase Inhibitor Cocktail, Thermo Scientific, USA, Cat #: 1861281). Protein concentrations were determined using a Bicinchoninic Acid (BCA) Protein Assay kit (Pierce™ BCA Protein Assay Kit, Thermo Fisher Scientific). Equal amount (10–15 μg) of cell lysate samples were loaded equivalently and separated on 4%–15% gel (Mini-PROTEAN® TGX™ Precast Gels, BIO-RAD). After transferring to nitrocellulose membranes (Amersham™ Protran® Western bloting membranes, Cytiva™), membranes were blocked for 30 minutes at RT with 1%–3% bovine serum albumin (BSA) in 1× Tris-buffered saline with Tween 20 (TBST) (Dyne Bio, Korea, Cat #: CBT3060). The membrane was then incubated with the primary antibody overnight at 4°C. The following primary antibodies were used: Cell signaling; p21 Waf1/Cip1, Cat #: 2947, MMP-3 Cat #: 14351, Phospho-Histone H2A.X (Ser139) Cat #: 9718, Lamin B1 Cat #: 13435 and Abclonal; β-Actin Cat #: AC026. After washing three times for 5 minutes in 1× TBST, the membranes were incubated for 1 hour at room temperature with a peroxidase-conjugated secondary antibody (Antirabbit IgG, Cell signaling, Cat #: 7074). After an additional three washes in 1x TBST, the membranes were developed using an enhanced ECL solution (WesternBright™ ECL, advansta, USA, Cat #: K-12045-D50). The intensity and band area of the proteins were analyzed using ImageJ software. β-actin was used as a loading control.
Cell proliferation assay
Seeded cells were treated with medium containing plasma and H2O2 in sequence according to the experimental scheme. The proliferation of HDFs was determined by using the Cell Counting Kit-8 (Dojindo, Japan, Cat #: CK04). According to the manufacturer’s instructions, the Water-Soluble Tetrazolium salt-8 (WST-8) was added to cells as 1:10 ratio to the cell culture medium and incubation was carried out for 2 hours in 37°C. The absorbance was measured with a microplate reader at 450 nm. Cell proliferation was calculated by comparing the OD values to those of the control HDFs.
Statistical analysis
The data were subjected to unpaired t-test or ordinary one-way analysis of variance (ANOVA) with multiple comparisons test Tukey. All results are represented as means ± standard error means (n ≤ 3). Statistical significance is implied as *p < 0.05, **p < 0.01, and ***p < 0.001.
Results
UCBP protects HDFs from H2O2-induced oxidative stress
To induce senescence, HDFs were treated with 600 μM H2O2 after a 4-hour pretreatment with 5% UCBP or ABP. The optimal H2O2 concentration was selected based on previous dose- and time-dependent studies. 9,28 –33 After senescence induction, SA-β-gal activity was markedly increased, confirming cellular senescent 38 (data not shown). Cells were first treated with 5% plasma for 4 hours, then incubated with 600 μM H2O2 in the plasma-containing medium for 20 hours. The following day, cells were washed twice with serum-free medium to remove residual H2O2 and incubated in plasma-containing medium for an additional 3–7 days (Fig. 1).
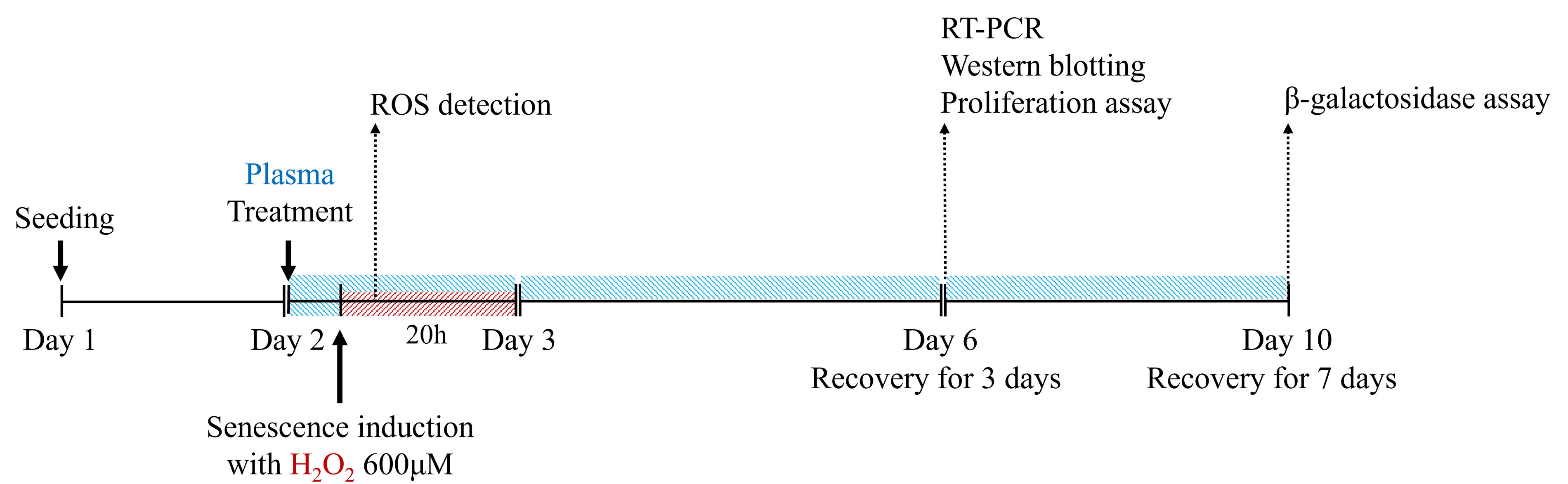
Experimental design for plasma treatment and oxidative stress induction in HDFs. HDFs were pretreated with 5% UCBP or ABP for 4 hours, followed by 600 μM H2O2 treatment for 20 hours, with plasma remaining present during oxidative stress induction. For ROS measurement, intracellular ROS levels were assessed 2 hours after H2O2, exposure using the DCFDA fluorescence assay. In all other conditions, after oxidative stress induction, cells were washed and maintained in 5% plasma-supplemented DMEM for 3–7 days to assess recovery. ABP, adult blood plasma; DMEM, Dulbecco’s modified Eagle medium; HDF, human dermal fibroblast; ROS, reactive oxygen species; UCBP, umbilical cord blood plasma.
Oxidative stress, primarily caused by ROS accumulation, is a key inducer of cellular senescence, as it triggers damage and SASP-related pathways. 7 To assess the protective effects of plasma, we compared pretreatment and cotreatment conditions using ABP or UCBP with H2O2. In the pretreatment group, HDFs were exposed to plasma for 4 hours, washed, and then treated with H2O2 in plasma-free medium. In the cotreatment group, plasma and H2O2 were added simultaneously. As shown in Figure 2, both pretreatment and cotreatment with plasma reduced ROS levels, with cotreatment demonstrating a stronger effect. These findings suggest that plasma, especially UCBP, mitigates oxidative stress in HDFs, which plays a central role in senescence induction.

Effect of plasma treatment on intracellular ROS levels in HDFs under oxidative stress. HDFs were treated with 5% UCBP or ABP under three conditions: (1) Pre-plasma (baseline ROS), where cells were pretreated with plasma for 4 hours, and ROS levels were measured before H2O2, exposure; (2) Pre-plasma (H2O2,-treated), where cells were pretreated with plasma for 4 hours, washed to remove plasma, and then exposed to 600 μM H2O2 for 2 hours without plasma; and (3) Co-plasma, which followed the same pre-plasma treatment but retained plasma during the subsequent 2-hour H2O2 exposure instead of being washed out. Control groups without plasma treatment, with or without H2O2 exposure, were also included. The underline in the figure indicates the time point of oxidative stress induction (H2O2 treatment). Intracellular ROS levels were measured using the DCFDA fluorescence assay, and fluorescence intensity was quantified using a microplate reader (Ex/Em = 485/535 nm). Data are presented as fold change relative to untreated controls and analyzed using one-way ANOVA followed by Tukey’s post hoc test ***p < 0.001. ABP, adult blood plasma; ANOVA, analysis of variance; DMEM, Dulbecco’s modified Eagle medium; HDF, human dermal fibroblast; ROS, reactive oxygen species; UCBP, umbilical cord blood plasma.
UCBP exhibits higher catalase levels than ABP
Catalase is a key peroxisomal enzyme that decomposes H2O2 into water and oxygen, contributing to antioxidant defense and redox homeostasis. 39 Reduced catalase activity has been implicated to age-related pathologies marked by oxidative damage and cellular senescence phenotypes. 40 Given this, catalase is considered a critical component in cellular antiaging mechanisms.
Although both UCBP and ABP reduced ROS levels, we focused on catalase as a representative ROS-scavenging enzyme, based on previous studies showing significantly higher catalase activity in UCBP compared to ABP. 41 To validate this, we quantified catalase levels in plasma samples using enzyme-linked immunosorbent assay (ELISA). Eight samples from each group were randomly selected and analyzed in triplicate. As shown in Figure 3, UCBP exhibited approximately threefold higher catalase concentration than ABP. This elevated catalase content may contribute to UCBP’s greater antioxidant capacity and stress mitigation, despite similar level of ROS reduction observed in Figure 2.

UCBP exhibits higher catalase levels than ABP. Catalase concentrations in ABP and UCBP were measured by ELISA. Data are shown as mean ± SEM (n = 8 per group), and significance was determined using unpaired t-test **p < 0.01. ABP, adult blood plasma; UCBP, umbilical cord blood plasma; ELISA, enzyme linked immunosorbent assay; SEM, standard error of the mean.
UCBP attenuates senescence markers induced by oxidative stress
To visualize H2O2-induced senescence, SA-β-gal staining was performed after culturing senescent HDFs in plasma-supplemented medium for 7 days. Equal numbers of cells were seeded into six-well plates. As shown in Figure 4A and 4B, over 80% of H2O2-treated cells exhibited SA-β-gal activity. ABP treatment modestly reduced this activity, while UCBP treatment resulted in a significantly greater reduction.

UCBP reduces senescence markers and DNA damage in HDFs.
To support these morphological observations at the molecular level, we measured mRNA expression levels of senescence markers including p16, p21, and Lamin B1 (primer sequences provided in Table 1). H2O2 exposure strongly upregulated p16 and p21, which were significantly attenuated by UCBP treatment. In addition, Lamin B1 expression, which decreased following H2O2 exposure, was restored in the UCBP-treated group, as confirmed by Western blot (Fig. 4C and 4D). Protein level of p21 and γH2AX, a DNA damage marker, was also reduced in response to UCBP treatment. These results suggest that UCBP alleviates oxidative stress-induced senescence by not only reducing ROS but also preserving nuclear and genomic stability.
UCBP suppresses SASP-associated inflammatory responses
Matrix metalloproteinases (MMPs) and interleukins (ILs) are key SASP and are closely associated with inflammation and tissue dysfunction. 10,42 To assess whether UCBP modulates SASP, we examined the mRNA expression of SASP markers including MMP1, MMP3, IL-1α, and IL-1β. H2O2 exposure significantly upregulated these markers, which were subsequently suppressed by plasma treatment. UCBP had more pronounced suppressive effect than ABP (Fig. 5A). At the protein level, MMP3 expression was also reduced in the UCBP group (Fig. 5B and 5C). To determine whether the antisenescent effects of UCBP extend beyond oxidative stress, we also examined SASP marker expression in replicatively senescent HDFs. As shown in Supplementary Figure S1, UCBP significantly reduced the expression of p16, p21, IL-1α, IL-1β, and PAI-1 compared with ABP and untreated senescent controls. These findings indicate that UCBP more effectively regulates SASP-related proinflammatory responses than ABP.
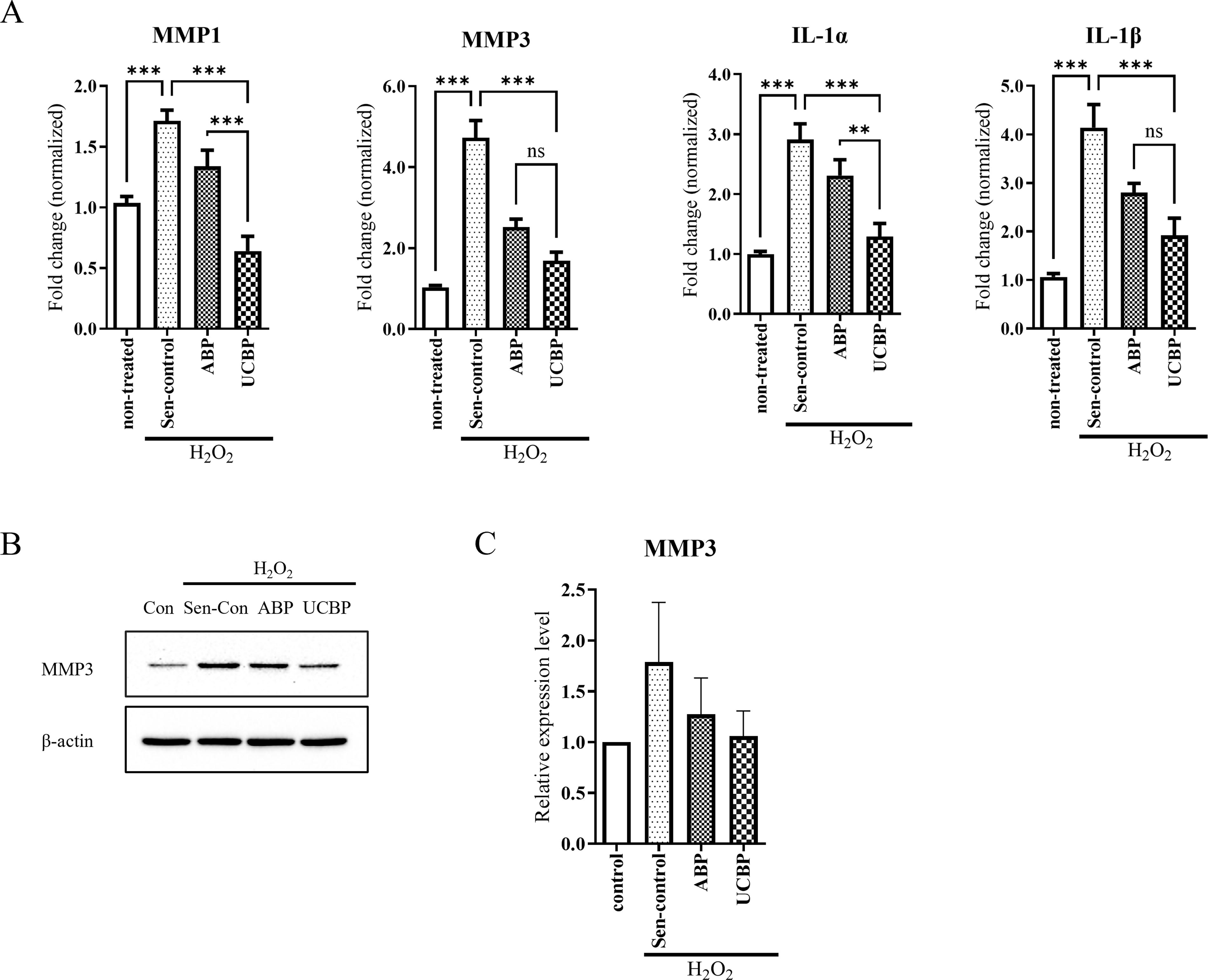
UCBP modulates SASP marker expression in senescent HDFs.
UCBP restores proliferation and cell cycle regulation in oxidatively stressed HDFs
Cellular senescence is characterized by irreversible growth arrest and morphological changes. 43 To assess UCBP’s regenerative potential, HDFs were subjected to H2O2-induced senescence following plasma pretreatment and subsequently incubated in plasma-containing medium for 3 days before proliferation analysis. As shown in Figure 6A, UCBP treatment significantly restored the reduced proliferation caused by H2O2 exposure, whereas ABP had minimal effect. Furthermore, the addition of 3-AT, a catalase inhibitor, attenuated this proliferative recovery in a dose-dependent manner, indicating that catalase activity contributed to the observed effects. Supplementary Figure S2 confirmed that the applied 3-AT concentrations were not cytotoxic to HDFs under either control or ABP-treated conditions.
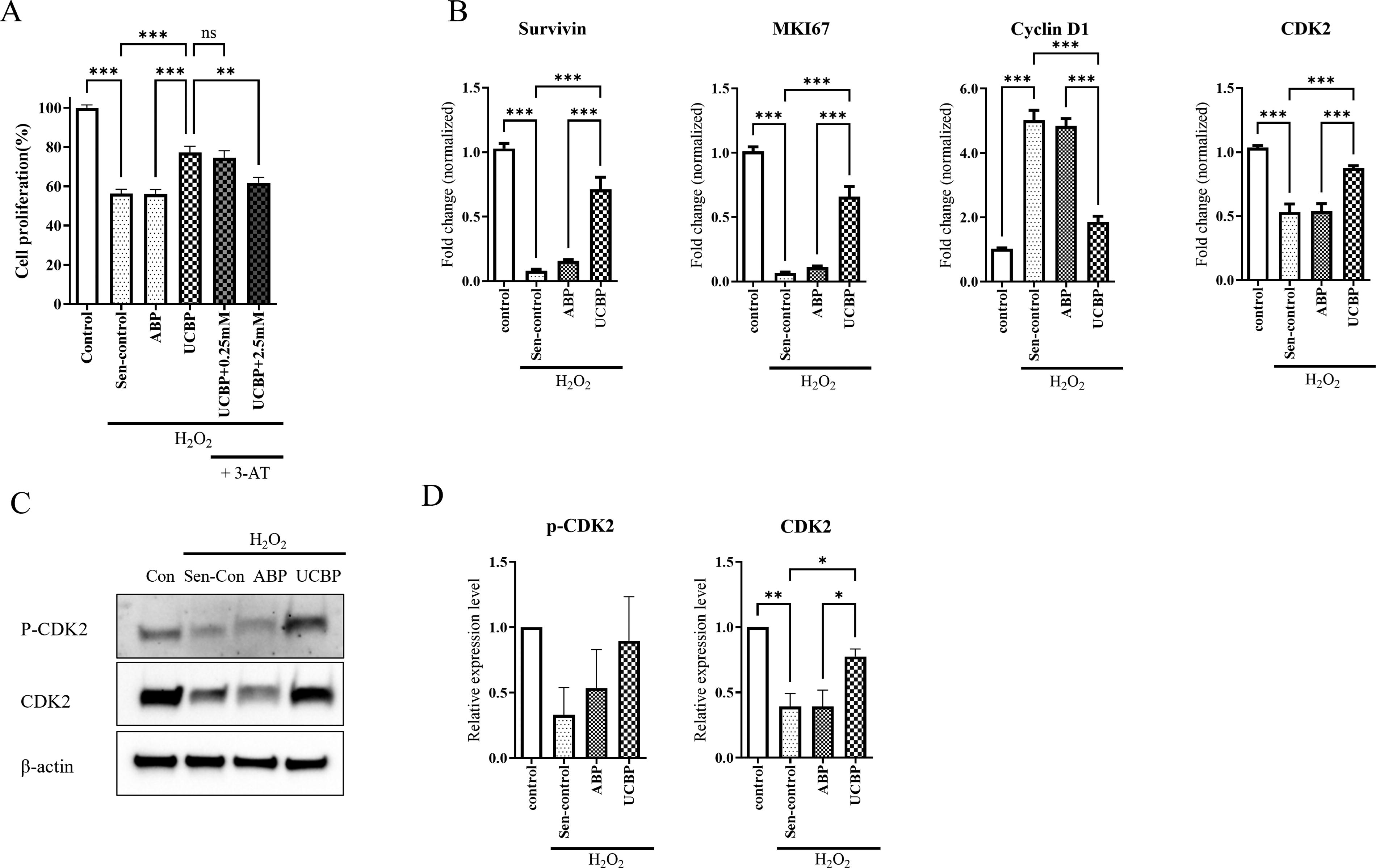
UCBP promotes proliferation and cell cycle recovery in oxidative stress-induced senescent HDFs.
To further investigate the molecular basis of this effect, we analyzed the expression of proliferation-related genes to assess transcriptional regulation. Survivin and MKI67, both downregulated by H2O2, were significantly restored by UCBP but not by ABP. Conversely, Cyclin D1, a marker of cell cycle arrest, was upregulated by H2O2 and remained elevated in the ABP group, while UCBP suppressed its expression. 44 Similarly, CDK2, a marker of cell cycle progression, was downregulated by H2O2 and remained low in ABP, but was recovered by UCBP treatment. Protein levels of p-CDK2 and CDK2 were also restored by UCBP (Fig. 6B and 6C). These findings suggest that UCBP not only enhances proliferation but also restores proper cell cycle regulation in senescent HDFs, highlighting its broader regenerative and antisenescent potential beyond ROS scavenging.
Discussion
Our findings demonstrate that both UCBP and ABP reduce ROS levels in H2O2-treated HDFs to a similar degree. However, UCBP’s protective effects cannot be solely attributed to ROS scavenging. Its higher catalase concentration suggests a selective detoxification of H2O2, which may explain its ability to mitigate oxidative stress-induced damage despite similar overall ROS reduction. Although UCBP contains higher catalase concentrations, ABP may possess other antioxidant components that account for its strong ROS scavenging activity. Future studies will expand the profiling of plasma antioxidant factors.
In addition, UCBP effectively suppressed senescence markers (p16, p21) and preserved fibroblast proliferation under oxidative stress, indicating that UCBP influences pathways beyond ROS reduction, possibly by modulating the SASP and enhancing tissue repair mechanisms.
UCBP’s ability to reduce proinflammatory SASP factors such as IL-1α, IL-1β, MMP1, and MMP3 suggests a mechanism through which it mitigates senescent cell-associated tissue dysfunction, a key driver of aging at the cellular level. 3 In addition to H2O2-induced senescence, UCBP also reduced SASP gene expression in replicatively senescent HDFs, as shown in Supplementary Figure S1. These results suggest that UCBP’s antisenescent effects extend beyond oxidative stress models and may be broadly effective across different types of cellular aging.
While cellular senescence involves the growth arrest of individual cells in response to damage, systemic senescence refers to the accumulation of senescent cells across tissues and organs, contributing to chronic inflammation and tissue degeneration. 45 Our in vitro results demonstrate UCBP’s ability to counteract cellular senescence in fibroblasts, preserving their functionality and proliferation under oxidative stress. However, further in vivo studies are necessary to explore the systemic consequences of UCBP’s protective effects and their potential impact on tissue dysfunction and inflammation.
Recent studies highlight the therapeutic potential of UCBP exosomes in promoting wound healing by enhancing fibroblast function and angiogenesis. 46 In addition, Mesenchymal Stem Cell (MSC)-derived exosomes have shown anti-inflammatory effects in psoriasis 47 and reduced UVB-induced skin aging. 48 While exosomes offer a targeted approach with fewer cellular risks, plasma-based therapies, such as those explored in our study, provide a broader range of bioactive molecules that may have synergistic effects in reducing oxidative stress and senescence. Future research should compare the efficacy and mechanisms of plasma versus exosome therapies in skin regeneration and antiaging.
Long-term studies will be crucial to evaluate UCBP’s safety, efficacy, and applicability across various tissues, particularly in the context of age-related diseases and oxidative stress-related conditions. 49 These studies will help assess UCBP’s potential to modulate systemic senescence and mitigate age-related tissue inflammation and degeneration, offering a more comprehensive antiaging strategy.
In summary, UCBP’s effects extend beyond ROS reduction to influence both cellular repair and senescence pathways. This multifaceted action suggests its potential as a comprehensive antiaging therapy, targeting not only oxidative stress but also key processes involved in cellular dysfunction and aging. While our in vitro findings highlight UCBP’s ability to mitigate cellular senescence, future in vivo studies will be essential to explore its role in preventing systemic senescence and further establish its therapeutic potential in antiaging interventions.
Footnotes
Authors’ Contributions
M.J.: Conceptualization, methodology, investigation, data curation, validation, writing—original draft. H.L.: Conceptualization, data curation, validation, visualization. T.-H.K.: Investigation, validation, visualization. S.J.C.: Conceptualization, supervision. W.O.: Project administration. S.K.: Conceptualization, project administration, validation, writing—reviewing and editing.
Author Disclosure Statement
The authors declare no conflicting interests.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.