
Abstract
We have isolated a novel progenitor cell population from adult rat pancreatic ducts, termed pancreatic-derived progenitor cells (PDPCs). Here, we report the in vitro culture, selection, and characterization of Thy1.1-positive and Thy1.1-negative PDPC subpopulations. These cells exhibit bipotentiality for differentiation into both pancreatic and hepatic cell types. Significantly, they express Pdx-1. Using a serum-free FGF-4-containing differentiation protocol, we have observed a time course of both morphological and gene expression changes indicative of hepatic lineage differentiation for the Thy1.1-positive subpopulation. These cells express albumin and store glycogen, typical features of mature hepatocytes. The Thy1.1-positive subpopulation could also readily be induced to differentiate into a pancreatic lineage with characteristic morphological changes resulting in three-dimensional islet-like structures and the transcriptional expression of insulin and glucagon in addition to Pdx-1. No morphological evidence of islet-like clusters was observed using the Thy1.1-negative population. However, Thy1.1-negative cells grown in pancreatic differentiation medium did show insulin gene transcription. Glucagon was not expressed in the undifferentiated Thy1.1-negative cells, nor was it induced in vitro after differentiation. The detection of Pdx-1 transcriptional expression in both populations indicates their potential as a novel source of non-β-cell-derived insulin.
Introduction
T
Several putative pancreatic progenitor cells have been characterized from both pancreatic ductal and islet cells [8 –10]. One of these cell populations has been observed, after differentiation in vitro, to express alpha-fetoprotein (AFP) and c-Met, proteins typically expressed by liver cells [11]. More recently, a mesenchymal stem cell population isolated from human pancreatic ductal epithelium has been reported to have the potential for pancreatic, hepatic, and mesodermal differentiation [12]. Although these cells could be induced to express Gata4, albumin, and TAT (tyrosine aminotransferase), no functional assessment has been reported. A further cell type, pancreatic-derived progenitor cells (PDPCs), isolated from adult rat pancreatic ducts has been reported to show multipotency and functional efficacy in an STZ diabetes model [13]. PDPCs also express Thy1.1, a cell surface protein that is also expressed on mesenchymal stem cells and stem cells of the fetal liver, umbilical cord blood in humans, mouse, and rat. Furthermore, in adult rat liver, a population of putative Thy1.1-expressing stem cells can be induced by chemical injury, or partial hepatectomy. These oval cells are bipotential, retaining the ability to differentiate in vitro into both the bile duct epithelium and hepatocyte lineages, contribute to liver repair, and express CD34, Sca-1, AFP, and CK19 [14,15].
Given the observations of hepatocellular-like cells within the pancreas and expression of Thy1.1 on unsorted PDPCs, the aim of this study was to characterize and assess the pancreatic and hepatic differentiation potential of Thy1.1-positive and Thy1.1-negative PDPC populations.
Materials and Methods
Isolation and maintenance culture of rat PDPCs
Pancreatic ducts were isolated from 12-month-old Albino Swiss (Glasgow) rats by dissection and minced, prior to seeding in CMRL medium. The PDP cells emerged as a confluent monolayer after ∼5 weeks in culture. These were then harvested and washed in PBS. PDPCs were maintained in culture in 20 mL CMRL 1066 medium (Invitrogen, Paisley, UK) supplemented with 10% Fetal Bovine Serum (Sigma, Poole, UK), 2 mM Glutamax, 1.25 µg/mL Amphotericin B, and 100 U/mL Penicillin, 100 µg/mL Streptomycin, (all Invitrogen) in T75 culture flasks with 0.2-µm filter caps (Corning, UK) at 37°C in a 5% CO2 atmosphere. Subconfluent cultures were passaged by the total removal of culture medium by pipette and the washing of the adherent cells by the addition of 10 mL calcium- and magnesium-free Hanks Balanced Salt Solution (HBSS) (Cambrex Bio-Science, Wokingham, UK) to the flask for 5 min at room temperature. After the removal of the HBSS from the flask by pipette, 2 mL of Trypsin–Versene solution (200 mg/L Versene, 500 mg/L Trypsin) was added to the flask. The flask was periodically examined microscopically until dissociation of the cell monolayer was confirmed. Cells were then removed by pipette and recultured, by the addition of 20 mL of fresh culture medium.
PDPCs were maintained long-term in CMRL 1066 medium (Invitrogen) supplemented with 5% Fetal Bovine Serum (Sigma), 2 mM Glutamax, 1.25 µg/mL Amphotericin B, and 100 U/mL Penicillin/Streptomycin (all Invitrogen) in T75 with 0.2-µm filter caps at 37°C in a 5% CO2 atmosphere. PDPCs were grown in a 37°C, humidified 5% CO2 atmosphere as a monolayer, and were passaged when 90% confluent with trypsin–EDTA (Invitrogen).
Magnetic activated cell sorting
Magnetic activated cell sorting (MACS) was performed for isolation and depletion using 1 µg of primary antibody, mouse anti-rat Thy1.1 (CD90) (Serotec, Oxford, UK) per 106 target cells for 20 min at 4°C as per the manufacturer’s protocol (Dynabeads Goat anti-mouse IgG (Dynal Biotech)). Sorted cell populations were resuspended in maintenance culture media and replated in tissue culture flasks. MACS was performed on each positive and negative sorted population twice before use in experiments. All sorted populations were checked with fluorescence-activated flow cytometry before use in subsequent differentiation experiments.
Flow cytometry assessment of cell surface antigens
Cells were resuspended in 0.5% BSA in HBSS. They were then centrifuged at 1,000× rpm for 10 min and the resulting cell pellet resuspended in HBBS. After a viability count with Trypan blue (Invitrogen, Paisley, UK), 1 × 106 cells/mL were labeled with 100 µL primary antibody. Primary antibodies used were against CD90 (Thy1.1) (Serotec MCA47R, 1:75), CD44 (Serotec MCA643, 1:10), CD49f (Serotec MCA2034, 1:50), CD147 (Serotec MCA729, 1:10), c-KIT (Santa Cruz Biotechnology, CA, USA; SC-19983, 1:20), CD71 (Serotec MCA 155FT, 1:10), CD24 (BD Biosciences 551133, 1:50), CD45 (BD Biosciences 554875, 1:50), CD31 (Serotec MCA1334GA, 1:50), and CD34 (Santa Cruz sc-7324, 1:50). Secondary antibodies were added in 0.2% BSA/PBS for 45 min at 4°C in the dark. The cells were then washed and centrifuged three times at 1,000× rpm in 0.2% BSA/PBS before labeling by the addition of 100 µL FITC-conjugated Fab2 fragment of Rabbit anti-Mouse Immunoglobulins (Dako Cytomation, Ely, UK 1:20) in 0.2% BSA for 45 min at 4°C in the dark. An isotype FITC control was also performed. After washing three times and centrifuging as before, the resulting cell pellet was resuspended in 1 mL of HBSS and the cells were analyzed using a Beckman Coulter XL Flow Cytometer (Beckman Coulter, High Wycombe, UK).
Differentiation experiments
For pancreatic differentiation Thy1.1-positive and Thy1.1-negative cell populations were plated at 6,600 cells/cm2 cell density. After 24 h, maintenance media was removed and monolayers were washed thrice with HBSS. Cells were subsequently cultured in DMEM:F12 (Lonza, Basel, Switzerland) supplemented with 1× ITS, 1.25 µg/mL Amphotericin B, 100 U/mL Penicillin/Streptomycin (all Invitrogen), Nicotinamide 10 mM (Sigma), KGF 10 ng/mL (Sigma), and 0.2% BSA (Sigma).
For hepatogenic differentiation cells were plated at 6,600 cells/cm2 in T75 and six-well plates, and at 2,500 cells/cm2 in chamber slides (Nunc, NY, USA) after 24 h; cells were washed thrice with HBSS, and maintenance media replaced with DMEM:F12 (Lonza) supplemented with Fibroblast Growth Factor-4 10 ng/mL (Sigma), 1 × ITS, 100 U/mL Penicillin/Streptomycin (Invitrogen), and 0.2% Bovine serum albumin (Sigma). Medium changes were performed thrice weekly and cells were harvested for RNA extraction from undifferentiated Thy1.1-positive and Thy1.1-negative cells at day 0 and day 28 for pancreatic differentiation and at days 0, 7, 14, 21, and 28 for hepatic differentiation. Cells undergoing hepatic differentiation in chamber slides were washed twice with PBS and fixed with 4% paraformaldehyde for 15 min at room temperature between days 10 and 14. Undifferentiated Thy1.1-positive and Thy1.1-negative populations were also grown in chamber slides concurrently and were fixed as above at 90% confluency.
Immunofluorescence
For staining of intracellular proteins, cells were fixed as above. Cells were thrice washed in PBS and permeabilized with 0.1% Triton X-100 (Sigma-Aldrich) for 10 min. Slides were incubated with donkey serum for 20 min and then incubated with previously optimized primary antibodies diluted in 0.5% BSA/PBS against rat albumin (Abcam, Cambridge, UK; ab14255, 1:100), CK19 (Biodesign Int., M08029M, 1:100), CK7 (Chemicon, Hertfordshire UK; MAB3226, 1:100), CK18 (Sigma F-4772), and Vimentin (Abcam ab8979, 1:50) for 1 h. Slides were washed thrice in PBS followed by the appropriate FITC-labeled secondary antibody (Abcam ab6749 or Dako Cytomation F0313, Ely, UK). Omission of the primary antibody was performed as negative control. Frozen sections of rat liver were used as positive controls. Slides were washed three times before mounting in Vectashield (Vector Laboratories, CA, USA) and were visualized and photographed by fluorescence microscopy.
RT-PCR
Total RNA was extracted using Trizol® according to the manufacturer’s instructions and quantified using the GeneQuant analyzer. Samples were DNAse treated (Ambion, TX, USA) and reverse transcription to cDNA was performed using SuperScript II reverse transcriptase (Invitrogen) according to manufacturer’s instructions. RT-negative controls were performed for all samples. RT-PCR was performed using Taq Polymerase (Invitrogen). The housekeeping gene β-actin was used to assess template quality. All PCR were performed using a Peltier Thermal Cycler-200. Nested PCR was performed for pancreatic differentiation experiments.
The following specific olignucleotide primers were used for PDX-1, Insulin II, and Glucagon (PDX-1: forward 5-cggccacacagctctacaagg-3, reverse 5-ctccggttctgctgcgtatgc-3, nested reverse 5-ttccaggcccccagtctcgg-3 (305 bp); Insulin: forward 5-atggccctgtggatccgctt-3, reverse 5-tgccaaggtctgaaggtcac-3, nested forward 5-cctgctcatcctctgggagcc-3 (209 bp); Glucagon: forward 5-gaccgtttacgtggctgg-3, reverse 5-cggttcctcttggtgttcatcaag-3, nested forward 5-acaaggcagctggcagcatgc-3 (210 bp)). Rat pancreatic total RNA was reverse-transcribed and used as a positive control. The following specific oligonucleotide primers were used for hepatic differentiation experiments: albumin (141 bp) forward 5-ctgggagtgtgcagatatcagagt-3, reverse 5-gagaaggtcaccaagtgctgtagt-3; HNF3β (63 bp) forward 5-cctactcgtacatctcgctcatca-3, reverse 5-cgctcagcgtcagcatctt-3; HNF1α (138 bp) forward 5-agctgctcctccatcatcaga-3, reverse 5-tgttccaagcattaagttttctattctaa-3; Gata4 (173 bp) forward 5-catgcttgcagttgtgctag-3, reverse 5-attctctgctacggccagta-3; Alpha-Fetoprotein (124 bp) forward 5-gtcctttcttcctcctggagat-3, reverse 5-ctgtcactgctgatttctctgg-3; CYP2B1(549 bp) forward 5-gagttcttctctgggttcctg-3, reverse 5-actgtgggtcatggagagct-3; CK19 (193 bp) forward 5-agtaacgtgcgtgctgacac-3, reverse 5-agtcgcactggtagcaaggt-3; CK18 (70 bp) forward 5-ggacctcagcaagatcatggc-3, reverse 5-ccacgatcttacgggtagttg-3. The PCR products then underwent agarose gel electrophoresis and were visualized by ethidium bromide staining. Rat liver tissue was used as a positive control.
Periodic acid Schiff staining
Periodic acid Schiff (PAS) staining for glycogen storage was performed on undifferentiated Thy1.1-positive and Thy1.1-negative cells and on Thy1.1-positive and Thy1.1-negative populations at day 21 of hepatic differentiation. Human liver sections were used as positive controls. Cells were fixed in 4% paraformaldehyde at room temperature for 10 min. Cells were thrice washed in PBS and permeabilized with 0.1% Triton X-100 (Sigma-Aldrich) for 10 min and washed twice with PBS and once with ddH2O. Cells were immersed in periodic acid solution (1 g/dL) for 5 min at room temperature. Wells rinsed in distilled water three times. Cells were then immersed in Schiff’s reagent for 15 min at room temperature and then washed in running tap water for 5 min. Cells were counterstained in hematoxylin solutions for 90 s and then rinsed in running tap water for 15–30 s.
Results
Characterization of Thy1.1-positive and Thy1.1-negative populations
MACS of PDPCs was used to isolate Thy1.1-positive cell populations expressing Thy1.1 at >98.5% and Thy1.1-negative cell populations at 98.7% purity, respectively. These populations were then cultured and reassessed by flow cytometry regularly every 10–12 days prior to any differentiation or characterization experiment.
Phenotypically, Thy1.1-positive and Thy1.1-negative populations demonstrated distinct differences in morphology: the Thy1.1-positive population exhibited a fibroblast-like morphology (Fig. 1A), whilst Thy1.1-negative population exhibited a more epithelial-like morphology (Fig. 1D).

Hepatic and pancreatic differentiation in vitro for Thy1.1-positive and Thy1.1-negative pancreatic-derived progenitor cell (PDPC) populations. (
The expression of cell surface markers between the Thy1.1-positive and Thy1.1-negative cell populations was similar but not identical. Both cell lines expressed CD147, CD44, and CD49f. Both showed low levels of CD71 expression and did not express the hematopoietic markers CD31, CD34, CD45, and c-kit. However, in contrast to the Thy1.1-positive-sorted cell population, the Thy1.1-negative population was positive for CD24 (Fig. 2A and B).
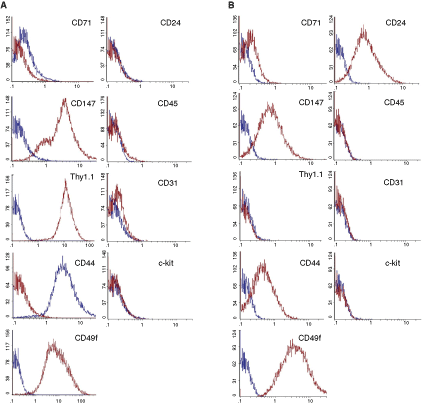
Cell surface characterization of magnetic activated cell sorting (MACS) of pancreatic-derived progenitor cells (PDPCs) by fl ow cytometry. (A) Thy1.1 (CD90)-positive populations are negative for the expression of CD24, CD31, CD45, and c-kit and low for CD71, but positive for the expression of CD147, CD44, and CD49f. (B) Thy1.1 (CD90)-negative populations are negative for the expression of CD31, CD45, and c-kit and low for CD71, but positive for the expression of CD24, CD147, CD44, and CD49f.
The subpopulations were also assessed by immunocytochemistry with hepatic, biliary, and mesenchymal markers, namely albumin, vimentin, CK7, and CK19 (Fig. 3).
Both Thy1.1-sorted populations were negative for albumin, CK7, and vimentin (Fig. 3B, C, F, G, J, and K). The Thy1.1-positive population was also negative for CK19 (Fig. 3O), whereas the Thy1.1-negative cells were weakly positive (Fig. 3N). Both populations were positive for c-Met and nestin by RT-PCR (data not shown).
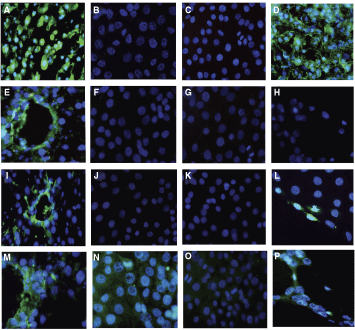
Immunocytochemistry for Thy1.1-sorted pancreatic-derived progenitor cell (PDPC) populations. Positive controls are shown for albumin (
Differentiation capacity of Thy1.1-positive and Thy1.1-negative populations
Pancreatic differentiation. The Thy1.1-positive and Thy1.1-negative populations exhibited markedly different morphological changes in pancreatic differentiation media. The Thy1.1-positive population, initially in fibroblast-like morphology, formed matted cell clusters by days 14–21. These formed into islet-like spherical clusters by day 28, which eventually detached from the parent cell layer (Fig. 1D,
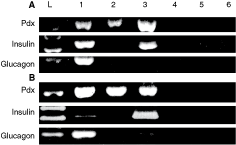
RT-PCR analyses of Thy1.1-negative pancreaticderived progenitor cells (PDPCs) (
We further examined the differentiation of the two populations by RT-PCR over a 28-day period. RT-PCR was performed for endodermal-specific genes HNF3β and GATA4 CK19, early liver makers, AFP and CK18, mature liver markers HNF1α, albumin, and the cytochrome P450 enzyme CYP2B1.
Undifferentiated Thy1.1-negative cells expressed HNF3β and CK19 by RT-PCR, but did not express albumin, CK18, HNF1α, CY2B1, GATA4, or AFP (Fig. 5). Undifferentiated Thy1.1-negative cells were negative for CK7, vimentin, and albumin expression and weakly positive for CK19 by immunocytochemistry. None of the other early or mature liver markers, including GATA4, AFP, and CY2B1, were induced in the Thy1.1-negative population in serum-free, FGF-4-containing media.

RT-PCR analyses of Thy1.1-positive pancreatic-derived progenitor cells (PDPCs) and Thy1.1-negative PDPCs in hepatic differentiation media. Thy1.1-positive and Thy1.1-negative PDPCs were cultured in hepatic differentiation medium for up to 28 days. Expression profiles were checked every 7 days. Column: (1) positive control, (2) undifferentiated PDPCs, (3) day 7, (4) day 14, (5) day 21, (6) day 28, (7–13) no RT controls for samples 1–6.
Undifferentiated Thy1.1-positive PDPCs expressed the early endodermal markers HNF3β, GATA4, and AFP, but did not express later markers of hepatocyte differentiation, HNF1α and albumin, until day 7 of hepatic differentiation (Fig. 5). The induction of albumin expression was confirmed by immunocytochemistry. Undifferentiated cells stained negatively for albumin content (Fig. 3A), while day 14 differentiated cells were strongly positive for albumin staining (Fig. 3D). CK18 was also expressed in undifferentiated Thy1.1-positive cells and throughout the 28-day differentiation period. CYP2B1 was present in undifferentiated Thy1.1-positive cells and throughout the differentiation period.
The undifferentiated Thy1.1-positive population was negative for CK19 expression by immunocytochemistry, but CK19 mRNA expression was induced during days 7–28, as detected by RT-PCR. CK19 is normally expressed by biliary cells, and this is consistent with the appearance of the luminal structures in culture. Interestingly, differentiated cells at days 10 and 14 time points did not stain for CK7 by immunocytochemistry but did so for the biliary marker CK19 (Fig. 3N and G) and for vimentin in the lumen-like structures only (Fig. 3L).
Periodic acid Schiff staining for glycogen storage
The presence of stored glycogen, as determined by PAS staining, was not observed in Thy1.1-positive or Thy1.1-negative PDPCs, nor in day 21 differentiated Thy1.1-negative cells. However, positive staining with PAS, indicative of glycogen storage, was observed in the Thy1.1-positive differentiated cells by day 21 (Fig. 6).

Glycogen storage in differentiated pancreatic-derived progenitor cells (PDPCs). Positive control (
Discussion
In this study, we report the in vitro culture, selection, and characterization of Thy1.1-positive and Thy1.1-negative PDPC subpopulations derived from adult tissue. Furthermore, we describe their potential with respect to differentiation to pancreatic and hepatic lineages and describe a cell population sorted using the marker Thy1.1, which displays lineage bipotentiality in vitro.
Thy1.1 is a cell surface protein whose function is not clearly understood. However, it has been suggested to be involved in cellular recognition [16,17], cellular adhesion [17 –19], and signal transduction [20]. Thy1.1 expression has been observed in various stem cell populations; notably, the oval cell population in adult rat liver has led to the supposition that Thy1.1 may have such a role in adult stem cells, allowing them to recognize and adhere to stromal tissue potentially as repair cells after injury [14,17,21,22]. Thy1.1 is also expressed on stem cells of the fetal liver, umbilical cord blood, and mesenchymal stem cells in humans, mouse, and rat. Our findings of greater in vitro differentiation potential within the Thy1.1-positive population would be consistent with these observations. They also demonstrate a method of isolation and purification with which to enable further characterization of such cells.
The exact nature and markers of any facultative pancreatic progenitors are still controversial. This may be confounded by an inherent quiescent nature in such cells, making them difficult to detect in vivo until activated or isolated from pancreatic tissue grown in culture [23].
However, candidate islet progenitor cells have been described previously based upon expression of the neural stem cell marker nestin and lack of known islet and duct cell markers [11]. In addition, Seaberg et al. have described the culture of cells from adult mouse pancreas, which expressed markers of both neurons and pancreatic islet cells. However, these cells, unlike PDPCs, appear to have a limited capacity for self-renewal and do not express the stem cell marker Oct-4. In addition, after differentiation, pancreatic proteins were expressed only in 4%–6% of cells [21,24]. Several putative pancreatic progenitor cells have been characterized from both pancreatic ductal and islet cells. Most have undertaken phenotypic description and examined pancreatic differentiation and assessment in transplant experiments. However, none have subselected populations in vitro for assessment of differentiation potential [8 –10,25]. Zulewski et al. observed expression of AFP and c-Met, also observed in Thy1.1-positive PDPCs. Such proteins are typically expressed by liver cells [11]. More recently, a mesenchymal stem cell population isolated from human pancreatic ductal epithelium has been reported to have the potential for pancreatic, hepatic, and mesodermal differentiation. Although these cells could be induced to express GATA4, albumin, and TAT (tyrosine aminotransferase), no functional assessment was reported
Previous studies have demonstrated hepatic differentiation of a number of different cell types, including bone marrow-derived MSCs, MAPCs, endometrial- and pancreatic-derived MSCs [12,26 –28]. The Thy1.1-positive subpopulation of PDPCs shares morphological phenotype and cell surface marker profile with these populations, including CD44+, CD24−, CD45−, CD31−. In contrast to this, however, Thy1.1-positive PDPCs appear to be a distinct cell type, expressing GATA4, HNF3β, and AFP, which have not been described as being expressed for any of these other cell types.
Furthermore, in adult rat liver, after induction by chemical injury, or partial hepatectomy, oval cells can be stimulated. These retain the ability, in vitro, to differentiate into both the bile duct epithelium and hepatocyte lineages and contribute to liver repair. While they express CD90 and AFP, in common with CD90+ PDPCs, they differ in that they are also CD34, Sca-1, and CK19-positive. It has also been hypothesized that liver resident precursors may be capable of pancreatic differentiation. Although fewer observation support this, Suzuki et al. identified a population of fetal liver cells, which were c-kit−, CD45−, Ter119−, and CD49f+/low, which could be differentiated into hepatic or biliary lineages, but also expressed mature pancreatic markers after transplant via common bile duct to pancreas in vivo. However, neither in vitro culture nor analysis of pancreatic differentiation was performed [29].
HNF3β is a marker of definitive endoderm believed to play an important role in endoderm competency [30], whilst GATA4 is a transcription factor required for ventral foregut endoderm development and for early liver gene expression [30,31]. HNF3β has been demonstrated to direct nucleosome positioning within the context of the albumin enhancer [32,33] with the subsequent facilitation of binding of GATA4 to the albumin enhancer. Both GATA4−/− and HNF3β−/− embryos show defects in foregut morphogenesis [34]. Therefore the expression of HNF3β and GATA4 in undifferentiated PDPCs and the subsequent FGF-4-stimulated induction of expression of liver-specific genes, such as albumin and HNF1α, are consistent with the proposal that HNF3β and GATA4 cooperate to control the potential of these cells to commit to a hepatic fate. Moreover, the presence of PAS staining in the Thy1.1-positive population after 21 days of differentiation demonstrated a functional characteristic of more mature hepatocytes, which is consistent with the expression of HNF1α. HNF1α is known to bind to genes whose products are related to mature hepatic functions, including carbohydrate storage and synthesis and lipid metabolism [35].
Undifferentiated Thy1.1-positive PDPCs express AFP. This observation is consistent with reports describing AFP expression in Nestin-positive islet-derived progenitor cells and low-level AFP and TTR expression, prior to hepatogenesis in the early ventral foregut endoderm. This expression is subsequently lost in endoderm isolated from cardiac mesodermal signaling [11,30,36]. It has also been suggested that this is a feature of the default pancreatic fate of ventral foregut endoderm [1]. Expression of AFP in the Thy1.1-positive PDPC population, which demonstrates capacity to differentiate into both pancreatic and hepatic lineages, would not be inconsistent with this finding [1].
Significantly, the undifferentiated Thy1.1-negative population, whilst expressing HNF3β, did not express GATA4 or AFP, nor were they induced during the differentiation experiment. No evidence of hepatic competency was observed in the Thy1.1-negative population. This is congruent with Pdx-1 expression and the absence of GATA4 expression within this undifferentiated population.
Vimentin was not expressed in either undifferentiated Thy1.1-positive or Thy1.1-negative populations, but was expressed in the cells forming the ductal-like structures during hepatic differentiation of the Thy1.1-positive population. Vimentin is considered to represent a mesenchymal marker. However, Masson et al. have observed coexpression of Thy1.1 and vimentin in portal structures, as well as demonstrating vimentin expression in epithelial cells within tissue sections and in culture of fetal liver epithelial cells [21].
The data pertaining to pancreatic differentiation are intriguing. No morphological evidence of islet-like clusters was observed using the Thy1.1-negative population. In contrast, Thy1.1-positive PDPCs could readily be induced to differentiate into a pancreatic lineage with characteristic morphological changes resulting in three-dimensional islet-like structures, the induction of transcriptional expression of insulin and glucagon, and the continued transcriptional expression of Pdx-1.
The detection of Pdx-1 transcriptional expression in both Thy1.1-positive and Thy1.1-negative undifferentiated populations indicates their potential to become insulin-producing cells. Notably, Thy1.1-negative cells when grown in differentiation medium, despite showing no morphological changes, expressed insulin. Glucagon was not expressed in the undifferentiated Thy1.1-negative cells, nor was it induced in vitro after differentiation (Fig. 4).
Various different candidate populations of pancreatic progenitor/stem cell have been described previously, including islet progenitor cells expressing nestin or other neuronal stem cell markers [8 –10,25], all of which appear capable, to a variable degree, of differentiating into insulin-producing cells in vitro. Another population have been shown to express Pdx-1, a known marker for insulin-producing cells, and these cells can stimulate both ductal and endocrine differentiation in vitro under appropriate conditions [37,38]. However, no assessment of the hepatic potential of these cell populations was reported. Moreover, there is evidence that pancreatic ductal epithelial cells have the potential to dedifferentiate to a progenitor cell capable of proliferation and formation of new islets and acini [39]. Recently, CK19+ nonendocrine pancreatic epithelial cells (NEPCs) were reported to be partially induced to differentiate into insulin-producing cells in vivo, when in the presence of fetal pancreatic tissue. However, they could not induce in vitro differentiation [40].
The precise physiological role played by these cells has been questioned. Dor et al. have challenged the view that neogenesis from ductal or progenitor cells occurs, instead arguing that β-cell replication, rather than new islet generation, is the predominant mechanism by which pancreatic endocrine tissue regenerates after near-total pancreatectomy [41], although this interpretation still remains controversial [42]. Our findings are consistent with a role for a non-β-cell that can produce insulin as potentially facilitating such a process. The significance of this observation remains to be determined, but these cells may offer an alternative insulin-producing cell source for transplantation therapies.
Recent work by Xu et al. has demonstrated that ngn3-positive facultative pancreatic progenitors are activated following pancreatic duct ligation and engender ngn3-mediated pancreatic regeneration, not observed in the injury models reported by Dor et al. and Teta et al. [23,41,43]. The latter do not induce ngn3-expressing cells [44]. These ngn3-positive cells were isolated and differentiated in explant cultures, but could not be expanded or maintained in culture. This implies that the adult pancreas, in certain situations, retains the capacity to regenerate through a progenitor population. This is significant, particularly given the observation that in normal pancreas ngn3-positive cells are not identified [23]. We have similarly sought to definitively identify PDPCs in the adult pancreas in vivo, without success (data not shown). The absence of identifiable PDPCs in normal rat pancreas is therefore not unsurprising in light of the findings of Xu et al. and in keeping with the concept of a facultative progenitor population.
This negative result is also consistent with reports of islet regeneration in transgenic models. A transgenic mouse model, in which the cytokine interferon-gamma (IFNγ) is expressed under control of the human insulin promoter, thereby directing its expression to pancreatic β-cells, demonstrated that ectopic expression of IFNγ results in new islet formation as well as expansion of pancreatic ducts [45,46]. Indeed, the site of new islet growth was found to be budding into the expanding pancreatic ducts. Interestingly, Kritzik et al. found regions of ducts in the transgenic mice that expressed HNF3β and PDX-1, but not Isl-1, insulin, or glucagon. Moreover, only a weak level of staining was observed in the ducts of nontransgenic and therefore nonregenerating mice [47]. These expression patterns observed are in keeping with the population of PDPCs we have isolated in vitro. Interestingly, both populations of PDP cells expressed HNF3β and Pdx-1, but only the Thy1.1-positive cells expressed GATA4.
Our observations on the Thy1.1-positive population are also in keeping with previous reports, in that they do not express insulin and glucagon when undifferentiated and are also negative for CK19, but demonstrate greater endodermal potency than previously reported [11].
We observed a time course of both morphologic and gene expression changes indicative of hepatic lineage differentiation by use of a serum-free FGF-4 containing differentiation protocol. The possible bipotentiality of embryonic ventral endoderm for pancreas and liver differentiation has been investigated in explant experiments where ventral endoderm differentiated to hepatic lineage by proximity to the cardiac mesoderm. The absence of inductive factors, such as FGF-1, FGF-2, and FGF-4, secreted by cardiac mesoderm allow the default pancreatic pathway of ventral endoderm to continue [1]. This is supported by the use of FGF signaling antagonists to inhibit hepatogenesis in vitro [1]. Our data are entirely congruent with this concept.
We have demonstrated isolation and characterization of PDPCs, which demonstrate potency and transcriptional responses to signaling consistent with a population of bipotential endodermal progenitors in vitro. Previously, administration of unsorted PDPC populations in a murine streptozocin-induced diabetes model demonstrated differentiation and production of rat insulin with concurrent stimulation of mouse pancreatic regeneration [13]. Further studies to examine in vivo effects of Thy1.1-sorted rat PDPC populations and human PDPCs are ongoing in diabetes and liver injury models and should provide insight into potential cellular therapies and regenerative medicine.
Footnotes
Acknowledgments
This work was supported by funding from Darlinda’s Charitable Trust to P.S. The authors would like to thank J. Ross for advice and criticisms.
Author Disclosure Statement
None of the authors have any disclosure to declare. No competing financial interests exist.