
Abstract
Adult tissues are known to contain rare populations of stem cells with multilineage differentiation potential that are distinct from other resident tissue-specific stem cells. However, whether multilineage stem cells are involved in tissue development is uncertain, primarily because the identification and characterization of these cells in embryonic tissue primordia is not well established. We tested whether stem cells with multilineage potential are present within the early post-gastrula somite tissue. We show that clonally derived precursor cells generate colonies with self-renewal capacity and have both neurogenic and myogenic lineage potential. Somite colonies contain cells that express Sox2, nestin, and Sca1, but do not express genes indicative of somitic mesoderm specification. Furthermore, we demonstrate that this multilineage potential is not due to colony cells with a pluripotent epiblast identity or the selection of p75 receptor-positive neural crest stem cells. Despite utilizing a highly undifferentiated tissue source, colony formation was not enhanced relative to reported estimates of multilineage stem cells from adult muscle, a derivative of the embryonic somite. Thus, our findings suggest that a permissive in vitro environment is sufficient for the isolation of a discrete population of stem cells in the embryonic somite that may represent the earliest developmental precursor to adult muscle multilineage stem cells.
Introduction
I
Indeed, there are numerous studies demonstrating the presence of relatively rare populations of cells within postnatal and adult tissues that are hypothesized to function as stem cells with multilineage potential. For instance, several studies have reported the isolation of stem cells from skeletal muscle tissue with the capacity for self-renewal and differentiation into muscle cells (cardiac and skeletal) [9 –16], neural cells [12,13,16], hematopoietic cells [9], endothelial cells [10,11], as well as adipocytes and bone cells [11,15]. Interestingly, these rare cells with multilineage potential do not typically correlate with the muscle satellite stem cell population, in either location or molecular marker identification, suggesting that adult stem cell compartments are heterogeneous. Moreover, tissue-specific stem cells with multilineage potential share at least some features, such as clonal nonadherent proliferation in vitro and cell-surface marker expression (eg, Sca1), indicating that these cells have a distinct proliferative and differentiation capacity despite their widespread presence in many different mature tissues.
Despite these clear examples of stem cells with multilineage (eg, neural) potential in postnatal and adult muscle and other mesoderm-derived tissues, such as bone marrow [17], it is not clear whether these cells participate in the normal formation of muscle tissue. In other words, is there a developmental role for tissue-specific multilineage stem cells? This issue has been difficult to address, primarily because the identification of multilineage stem cells in embryonic tissues has been given comparatively little attention.
In order to determine whether multilineage stem cells exist in the embryonic precursor of adult muscle tissue (somitic mesoderm) at the earliest stages of development, we asked whether a well-characterized in vitro neural stem cell colony-forming assay could elicit the clonal proliferation of undifferentiated somite cells with myogenic and neurogenic differentiation capacity. We report that a subpopulation of early post-gastrula (9.5 dpc) somite cells have similar characteristics of neural stem cells when cultured in a permissive in vitro environment, generating neurons and glia as well as muscle cells. We demonstrate that cells within clonally derived somite colonies do not express epiblast-specific pluripotency markers, such as Oct4 and Nanog, and that the colony-forming somite cells do not express typical neural crest stem cell markers. The self-renewal and neurogenic potential, as well as the expression of Sox2, Sca1, and nestin, suggest that these mesoderm precursor cells share similar properties with the previously described adult multilineage stem cells [16,18]. Finally, the estimated frequency of the neurogenic mesoderm precursors is comparable to the reported frequency of adult muscle interstitial precursors [18]. Thus, our data suggest that even as early as 9.5 dpc, there is a discrete population of somite-derived precursor cells with neurogenic and myogenic potential that may represent the embryonic predecessors to the adult multilineage stem cell population within muscle tissue.
Materials and Methods
Mice
C57Bl/6-YFP, Wnt1-Cre, and Z/EG transgenic mice were a kind gift from Dr. Derek van der Kooy and were bred in-house in a SPF facility. The day of the vaginal plug was considered 0.5 days post-coitum (dpc) of gestation. CD1 WT (timed-pregnant from Charles River) and transgenic mice were studied at embryonic day 9.5 dpc. All mice were used in accordance with approved University of Toronto animal use protocols and Canadian Council on Animal Care guidelines.
Dissections
Female dams were sacrificed by cervical dislocation and embryos were removed and placed in a Petri dish containing 1× PBS (Dulbeccos; Invitrogen, Carlsbad, CA). The neural tube, somites, and heart tissue were dissected using a 27G-1/2″ syringe needle (B-D PrecisionGlide) or an electrolytically sharpened tungsten needle together with fine forceps (Dumont #5), placed in 1 mL serum-free media (SFM), triturated using a fire-polished Pasteur pipette, and viable cells were quantified using the Trypan blue exclusion method with a hemocytometer.
Tissue culture
Cells were cultured in vitro at a final density of 5–10 cells/µL in noncoated 24-well plates (Nunclon) with SFM (10× DMEM/F12 [1:1], 30% glucose, 7.5% NaHCO3, 1 M HEPES, 100×
Immunolabeling
Individual colonies were collected, gently triturated, and plated on Matrigel in 10% fetal bovine serum for 7 days to promote differentiation. Cells were fixed on ice with 4% paraformaldehyde for 15 min, washed in 1× PBS, permeabilized using 0.3% Triton X detergent for 20 min at room temperature, then blocked with 10% normal goat serum for 1 h at room temperature. The following primary antibodies were used: mouse anti-GFAP (1:500) (Sigma, St. Louis, MO), mouse anti-βIII-tubulin (1:500) (Sigma), mouse anti-MAP2 (1:500) (Chemicon, Temecula, CA), rabbit anti-Doublecortin (Dcx) (1:100) (Cell Signaling), human anti-HuC/D (1:100) (Molecular Probes, Eugene, OR), mouse anti-O4 (1:400) (MAB345, Chemicon), mouse anti-MF20 (1:10) (myosin heavy chain, Developmental Studies Hybridoma Bank), mouse anti-nebulin (Dr. Peter Zandstra), rabbit anti-skeletal muscle actin (1:200) (Abcam), mouse anti-MyoD1 (1:200) (Abcam), PE rat anti-mouse Ly-6A/E (Sca1, 1:100) (BD Biosciences, San Jose, CA), rabbit anti-Sox2 (1:100) (StemCell Technologies, Vancouver, BC), mouse anti-nestin (1:100) (Chemicon). Secondary antibodies were Alexafluor red 568 goat anti-mouse IgG or IgM (in the case of anti-O4), or Cy2 goat anti-mouse IgG (Molecular Probes; 1:200–400). For immunohistochemistry, 9.5 dpc embryos (CD1) were fixed overnight in 4% paraformaldehyde, rinsed in 1× PBS, and then cryoprotected with increasing concentrations of sucrose up to 30%. Embryos were then infiltrated with O.C.T. embedding compound (Miles, Inc.) at room temperature. Embedded embryos were frozen and cryosectioned at 20 µm and processed for immunolabeling as above. Cells and sections were counterstained with DAPI and examined using the Axiovert 2000 (Zeiss) or Leica DM4500B fluorescent compound microscope.
Flow cytometry
Primary somite tissue was dissected and dissociated into single cells in SFM as described earlier. Cells were washed 2 times in PBS containing 0.5% bovine serum albumin (BSA; Sigma), resuspended at 5 × 106 cells/mL, and kept on ice. Rabbit anti-mouse p75 NGF receptor polyclonal antibody (Chemicon) was added at a concentration of 0.1 µg/106 cells for 30 min on ice. For Sca1 sorting, cells were incubated with PE rat anti-mouse Ly-6A/E (Sca1) (BD Biosciences), washed 2 times in PBS/BSA, resuspended and incubated in secondary antibody (AlexaFluor 488 goat anti-rabbit IgG. 1:500; Invitrogen) for 30 min on ice. Following 2 times PBS/BSA washes, cells were suspended in PBS/BSA containing propidium iodide (1 µg/mL; Sigma), filtered through Nitex nylon membrane (Sefar America Inc., Kansas, MO) prior to sorting. Flow cytometric sorting was conducted using a Beckman Coulter Epics Elite Machine or FACSaria (Becton Dickinson, Franklin Lakes, NJ). Live cells were sorted for antibody-labeled cells used FACS gates set with unlabeled cells. Labeled and unlabeled fractions were collected separately in plating medium (described earlier) at equivalent densities (ranging from 1 to 5 cells per microliter depending on the numbers of cells isolated) in addition to unsorted live cells that were run through the FACS machine. The numbers of colonies were counted after 8–10 days in vitro.
RT-PCR
RNA was isolated from E9.5 somites and neural crest tissue. Tissue samples and colonies were homogenized and the RNA was prepared using RNeasy kit (Qiagen, Mississauga, ON). RT-PCR was performed using the OneStep RT-PCR kit (Qiagen, Mississauga, ON) using the following primers (see below). PCR was performed with Taq polymerase (Invitrogen) according to the manufacturer’s instructions for 32 cycles at 58°C annealing temperature and PCR products were separated out on a 2% agarose gel and stained for imaging with SYBR Safe DNA gel stain (Sigma-Aldrich, Oakville, ON). Gene-specific primer sequences used in these experiments are listed in Supplementary Table 1 (Supplementary materials are available online at http://www.liebertpub.com).
Results
Clonal proliferation of isolated precursor cells from somite tissue in serum-free neural colony growth-promoting cultures
Mouse epiblast cells derived from the early post-implantation period at 5.5 dpc retain their pluripotency when cultured as cell lines in vitro [19,20] and in general, epiblast cells maintain pluripotent potential until late gastrula to early neurula stages (7.0–8.0 dpc) when grafted to ectopic locations [21,22] or lineage traced [23] in vivo. Thus, we isolated cells from somite tissue at 9.5 dpc. At this stage, the somites contain undifferentiated stem and progenitor cells that give rise only to somite derivatives, such as dermis, bone, cartilage, and muscle, but not neural cells [24]. We isolated embryonic trunk tissue between the anterior and posterior limb buds (Fig. 1A-i) and removed the overlying epidermal ectoderm and underlying tissues (eg, dorsal aorta, notochord, gut) resulting in a trunk fragment typically containing 8–10 somite pairs surrounding the thoracolumbar segment of the spinal neural tube (Fig. 1A-ii). We then used a sterile electrolytically sharpened tungsten needle or a 27G-1/2″ sterile syringe needle to remove the somite tissue from either side of the neural tube such that ∼8–10 individual somites were dissected and pooled from a single embryo (Fig. 1A-iii). We purposely did not remove the somite tissue that is most proximal to the lateral wall of the neural tube (sclerotome and myotome precursors) to ensure no contamination of neural tube cells. This proximal somitic tissue was also removed for analyses of neural tube-derived cells (isolated from separate embryos).
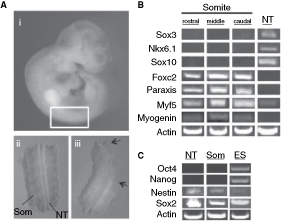
Reliable somite dissection in the absence of neural tube tissue. (
To confirm that our neural tube and somite dissections were not cross-contaminated, we performed RT-PCR on samples of dissected tissue and analyzed for somite- and neural tube-specific gene expression (Supplementary Table 1). We isolated rostral, middle, and caudal somites within the dissected region and assayed for expression of the spinal neural tube markers sox3, nkx6.1, and sox10. These genes were not expressed in the dissected somite tissue at any rostral–caudal level, but were expressed in neural tube tissue derived from the same region of the embryo (Fig. 1B). In contrast, expression of the somite genes foxc2, paraxis, myf5, and myogenin was present in somite tissues at all rostral–caudal locations (Fig. 1B). With the exception of myf5, none of these somite genes were expressed in dissected neural tube tissue. We detected low levels of myf5 expression in dissected neural tube tissue suggesting the possibility of selective contamination of the dorsal medial lip of the somite (boundary of myotome and dermomyotome), where myf5 is expressed in the absence of some other somite genes or the low level expression of this gene within the neural tube. Thus, these data demonstrate that somite tissue can be reliably removed from 9.5 dpc embryos in the absence of neural tube tissue.
We further determined whether dissected neural tube and somite tissues demonstrated the potential for pluripotency by examining the expression of oct4 (pou5f) and nanog, both of which have been shown to be required for pluripotency [25,26]. We compared the expression of oct4, nanog, sox2, and nestin by RT-PCR in isolates of 9.5 dpc neural tube and somite tissue, as well as from ES cells grown in standard LIF- and serum-supplemented culture media. Expression of nestin was detected in somite tissue as well as neural tube tissue, albeit at a relatively lower level (Fig. 1C). Very low levels of nestin were also detected in growing ES cell cultures, and this is consistent with previous observations of a low level of spontaneous neural differentiation in ES cell cultures [27]. Furthermore, sox2, which is present in the neural tube and in ES cells, was robustly expressed in all samples (Fig. 1C). In contrast, neither oct4 nor nanog was expressed in neural tube or somite tissues (Fig. 1C). Thus, consistent with published work, our data indicate that both of these embryonic tissues likely do not contain resident pluripotent cells during this early stage of development although it remains a formal possibility that a rare population of cells may exist that are undetectable using the methods employed.
In order to test whether single precursor cells isolated from somite dissections had colony-forming properties, we cultured cells at clonal densities in defined serum-free neural stem cell culture conditions, supplemented with EGF, FGF2, and heparin, as well as other factors known to promote neural stem cell survival and proliferation in SFM, such as B27, N2 supplement, N-acetyl-

Clonal colony formation in somite and neural tube cultures. (

Clonal derivation of somite and neural tube colonies as observed through cell mixing. Table represents the combined data from 3 separate experiments showing that low-density cultures of equally mixed cell genotypes (CD1 vs. YFP-transgenic) results in predominantly YFP-positive or YFP-negative colonies. Images on the right show a manually arranged group of colonies from a representative neural tube culture (above, phase-contrast image) containing both YFP-positive and YFP-negative colonies (below, fluorescence image). A typical mixed colony (above, arrow) shows a pattern of YFP-expressing and nonexpressing cells arranged as fused clumps, rather than a more uniform distribution of cell types. Abbreviations: NT, neural tube; YFP, yellow fluorescent protein.
The somite-derived colony-forming cells are self-renewing, regionally unspecified, and correspond to the Sca1+ cells present in somite tissue in vivo
Nonadherent colony formation in vitro is a property of multilineage stem cells isolated from postnatal and adult muscle tissues. To test whether somite-derived colony-forming cells have similar characteristics, we assayed for in vitro self-renewal as well as neuronal, glial, and muscle differentiation potential. Single somite or neural tube colonies of similar size were dissociated and replated in similar culture conditions and the numbers of newly generated colonies was counted after 7 days (Fig. 4A). Individual somite-derived colonies gave rise to approximately 1 new colony (primary colonies, n = 17; secondary colonies, n = 8) upon subcloning. In contrast, neural tube-derived colonies generated approximately 6 new colonies on average from primary (n = 252) or secondary (n = 131) subcloned colonies. These data indicate that the somite colonies, similar to neural stem cell-derived colonies from the neural tube, have the ability to undergo self-renewal and hence support the hypothesis that the somite colonies are derived from cells with self-renewal ability.
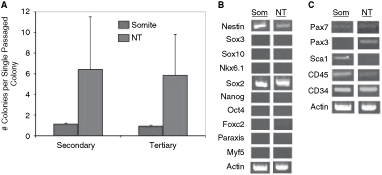
Distinct patterns of self-renewal and gene expression between somite and neural tube colonies. (
Previous studies have demonstrated that neural stem cell colonies derived from the 14.5 dpc brain transiently express selected genes that are indicative of regional specification [33]. On the other hand, neural colonies derived from the early post-gastrula neuroepithelium or from ES cells express neural-specific genes in a nonregionally specified manner [32]. We hypothesized that if somite-derived colonies exhibited properties of neural stem cell colonies then their gene expression profiles may be similar to those derived from the 9.5 dpc neural tube. In contrast, if these colonies are unlike neural stem cell colonies, then they may represent more primitive colonies (eg, ES cell colonies) or ones that are somite-specific despite their derivation in neural stem cell culture conditions. Using single-colony RT-PCR, we assayed for several neural tube, somite, ES cell, and muscle stem cell markers. Of the relatively specific markers used to identify neural tube identity (eg, sox3, sox10, nkx6.1), somite identity (eg, foxc2, paraxis, myf5), or ES cell identity (eg, oct4, nanog), none were expressed in either neural tube-derived colonies or somite-derived colonies (n ≥ 10 colonies from each of somite and neural tube) (Fig. 4B). However, several patterning genes that mark precursor populations in both neural plate/tube as well as pre-somitic and somitic mesoderm (eg, nestin, sox2, pax3, pax7) showed variable patterns of expression in colonies. For example, sox2 and nestin were quite abundantly expressed in somite colonies and neural tube colonies, whereas pax7 was only weakly expressed in both colonies. However, despite the fact that pax3 is expressed in dissected neural tube and somite tissue (data not shown) this gene was specific for neural tube colonies and not expressed in somite colonies (Fig. 4B). Recently, studies of embryonic and adult muscle precursors have identified a population of muscle stem cells that express Pax3 and Pax7, as well as other early stem/progenitor markers such as CD45, CD34, and Sca1 [17,34] that mark various stem and progenitor subpopulations, in particular from mesoderm-derived tissues. Both CD45 and CD34 are expressed in colonies derived from neural tube and somite tissues. In contrast, sca1 is not expressed in neural tube-derived colonies, but is specifically expressed in somite-derived colonies (Fig. 4C). Thus, of all of the markers analyzed, Sca1 and Pax3 are differentially expressed in somite versus neural tube colonies, respectively.
Sca1 was previously shown to mark multilineage stem cells in muscle tissue [eg, 16], therefore we examined whether Sca1+ cells were in fact present in 9.5 dpc somite tissue in vivo, or whether this marker was up-regulated in somite colony cells in vitro. We analyzed coronal and horizontal cryosections from the trunk region (n = 6 embryos; 2–3 sections each; Fig. 5) using a Sca1-specific antibody and detected a relatively small population Sca1+ cells scattered throughout the somitic compartment (Fig. 5A). These cells could be observed in small clusters or scattered in a proximal position along the neural axis (Fig. 5B, E), but often were scattered both within and surrounding the somites distal to the neural tube (Fig. 5H). Sca1+ cells could also be detected in the limb bud mesoderm (Fig. 5D), another location that we were able to derive clonal colonies with the properties of somite colonies (data not shown). Moreover, in vivo the Sca1+ cells in the somitic compartment coexpress Sox2 (Fig. 5C) and nestin (Fig. 5G, J), consistent with our colony RT-PCR data. Sca1+ cells were not observed in the trunk neural tube.
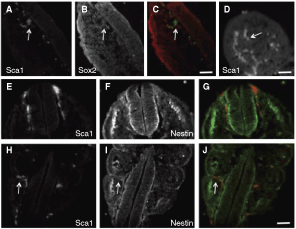
Sca1+ cells in 9.5 dpc embryos. Cryosection depicting the spinal neural tube surrounded by somitic mesoderm tissue immunolabeled for Sca1 (
The in vivo expression pattern of Sca1 corresponds with our observation that somite colony cells express Sca1 transcript, and suggests that the Sca1+ cells in vivo are the source of the clonally derived somite colonies. Indeed, Sca1+/Sox2+/nestin+ cells are present in a typical primary dissection of somite tissue (supplementary Fig. 1). In contrast, Sca1-expressing cells are not observed in neural tube dissections, despite the presence of numerous Sox2+ and nestin+ cells in these dissections (supplementary Fig. 2). Finally, consistent with the hypothesis that the Sca1+ cell is the somite colony founder cell, analyses of Sca1-expressing cells from somite dissections using flow cytometry reveal that only the Sca1+ cell fraction gave rise to somite colonies in vitro.
Neurogenic and myogenic potential of somite-derived colony-forming cells
Clonally derived progeny from neural stem cell colonies can be differentiated into neuronal and glial fates by replacing the mitogens with serum and culturing for 7 days. Under such differentiation conditions, neural stem cell colonies derived from neural tube cells along the entire neuraxis generate neurons (immunopositive for βIII-tubulin, MAP2, Dcx, HuC/D), astrocytes (immunopositive for GFAP), and oligodendrocytes (immunopositive for O4), which we confirmed in our differentiated neural tube colonies (data not shown). The expression of specific muscle lineage markers, such as nebulin, MF20, skeletal muscle actin, and MyoD1, are never observed in differentiated cultures of neural tube-derived colonies. Interestingly, differentiation of somite colonies in these standard neural stem cell differentiation conditions was sufficient to reveal neuronal and glial cells. In 1 representative experiment (n = 4 separate colonies analyzed), we observed cells that were βIII-tubulin/MAP2+ (7.1%, 43 of 605 cells quantified) with typical morphological characteristics of neurons, such as small rounded soma and long, thin and branched processes (Fig. 6A). These cultures also contained Dcx+ and HuC/D+ neurons (Fig. 6B, C). Other cells that were relatively large and flat were GFAP+ (31.9%, 190 of 595 cells quantified) (Fig. 6A) or small with short, highly branched processes expressing the oligodendrocyte marker O4 (2.6%, 4 of 151 cells quantified) (Fig. 6D). In addition, somite colony cells were found to express the muscle markers nebulin and MF20 (Fig. 6G) as well as skeletal muscle actin (Fig. 6E) and MyoD1 (Fig. 6F) (<1% of all cells quantified). In rare instances, the nebulin/MF20+ cells fused to form structures that appeared similar to primitive myotubes (Fig. 6H).
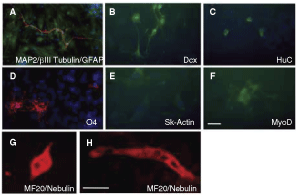
Somite-derived colonies contain neural and muscle cells. (
Colony-forming cells can be isolated from 9.5 dpc cardiac mesoderm
To address whether neurogenic potential was a general feature of early mesoderm or specific to somitic tissue, we asked whether colony-forming cells with neurogenic capacity were also present in cardiac mesoderm. Cardiac tissue was dissected from 9.5 dpc embryos with care to ensure that surrounding tissues (eg, aorta) were removed. Cells were plated in the same neural conditions and clonally derived colonies were generated after ∼7 days in vitro (Inset, supplementary Fig. 3A). Co-culture experiments with cardiac mesoderm from CD1 and YFP-transgenic cells demonstrated that the vast majority of colonies were clonally derived (Supplementary Fig. 3A). The 9.5 dpc-derived cardiac colonies had similar self-renewal capacity as the somite-derived colonies, generating approximately 1 new colony upon subcloning of primary or secondary colonies (0.9 ± 0.1 secondary colonies, n = 21; 0.8 ± 0.02 tertiary colonies, n = 7). Upon mitogen removal and differentiation in serum, cardiac mesoderm-derived colony cells differentiated into neurons and glia (>5 colonies examined for differentiation). In 1 representative experiment, βIII-tubulin/MAP2+ cells were detected (1.2%, 1 of 83 cells quantified) (Supplementary Fig. 3B), along with GFAP+ (31.3%, 26 of 83 cells quantified) (Supplementary Fig. 3C).Cells that morphologically resembled oligodendrocytes and expressed O4 were extremely rare (<1% from several separate colonies analyzed) (Supplementary Fig. 3D). Strikingly, cardiac mesoderm colonies were observed to beat synchronously in culture (Supplementary Video 1), revealing the presence of contractile muscle cells within the colony and these same beating colonies generated the neuronal and glial cells upon differentiation. Taken together these findings indicate that colony-forming precursors with neurogenic potential may be present in many post-gastrula mesoderm-derived tissues.
Somite-derived precursor colonies are not neural crest stem cell colonies
Neural crest precursor cells emerge from the mouse dorsal neural tube in the trunk region starting at ∼9.0 dpc [35]. Although these cells have not migrated extensively at this early time point, we considered the possibility that emigrated neural crest precursors in the surrounding somites were the colony founder cells in our assay. Since premigratory and migratory neural crest stem cells express Sox10 [36], we initially performed single-colony RT-PCR experiments and found that neither somite-derived colonies nor neural tube-derived colonies expressed sox10 (Fig. 4B), suggesting that these colonies are not derived from neural crest stem cells. To further support this conclusion, we performed 3 separate experiments to test whether somite-derived colonies are, in fact, neural crest stem cell colonies. First, we analyzed expression of the neurotrophin co-receptor p75, which is expressed in embryonic and postnatal neural crest stem cells [37], by single-colony RT-PCR. Neither neural tube-derived colonies nor somite-derived colonies expressed p75 (Fig. 7A), indicating that our culture conditions likely do not support the formation of clonal neural crest colonies. It is possible that colony-forming cells initially express p75 upon dissection, but down-regulate its expression in vitro over the course of colony formation. Since selection of p75-expressing cells by flow cytometry can significantly enrich for neural crest stem cells [37], presumably independently of their subsequent gene expression profile, we next sorted for cells enriched for p75 expression from our tissue isolates prior to culturing in the colony-forming assay. A significant number of cells from 9.5 dpc somite or neural tube dissections were found in the p75-expressing fraction (Fig. 7B). We analyzed the number of colonies derived from unsorted somite cells after recovery from the flow cytometer and cultured in the same media and cell density conditions as our primary cultures. From 4 independent samples, a total of 15 somite colonies were derived from unsorted cells (Fig. 7C). In contrast, we never observed colony-forming cells in the p75-positive fraction. All somite colony-forming cells were within the p75-negative fraction (Fig. 7C). Similar experiments with neural tube-derived cells also revealed that the colony-forming cells were present only in the p75-negative fraction (data not shown).
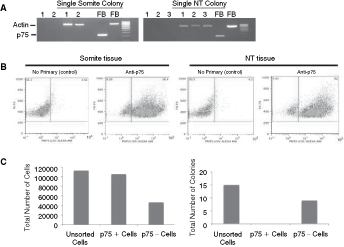
Somite colonies are not neural crest stem cell-derived. (
Finally, we crossed Wnt1-Cre transgenic mice to Z/EG transgenic reporter mice in order to assess whether any of the somite-derived colonies from 9.5 dpc dissections expressed Wnt1, which is expressed by multipotent neural crest cells [38,39]. If the colony-forming cells in our assay were derived from Wnt1-expressing neural crest cells, then recombination at the Z/EG locus (which activates GFP expression) would provide an indelible genetic lineage marker for all progeny of the colony-forming cell, regardless of whether endogenous Wnt1 expression was down-regulated in vitro. Although fewer somite colonies were generated from this mixed genetic background compared to the CD1 strains used in most of our experiments, none of the colonies generated after 7 days of in vitro culture were GFP+ despite the presence of viable GFP+ single cells that remained in the culture dish during this period (0/46 colonies, n = 3 separate experiments). Thus, our findings suggest that the somite-derived colonies with neurogenic potential are not derived from neural crest stem cells.
Discussion
We demonstrate that early post-gastrula somite tissue contains a precursor cell subpopulation with both neurogenic and myogenic potential. The overall fraction of somite cells with clonal colony-forming capacity in serum-free, nonadherent conditions was ∼0.1%, which is consistent with reports of colony-forming neurogenic interstitial precursors (or stem cells) found in adult muscle [18]. Thus, despite the undifferentiated nature of the entire cell population analyzed during a very early developmental stage of tissue development, the fraction of colony-forming neurogenic precursors is not enhanced relative to their putative adult-derived counterparts. One prediction from the outset of this study was that the proportion of stem cells with clonal colony-forming neurogenic potential would be greater in a population of embryonic somite cells compared to adult muscle, given the relative enrichment of undifferentiated stem/progenitor cells from the embryonic tissue. However, our findings do not support the general notion that the most undifferentiated cells in a population are necessarily more responsive to permissive environmental cues that reveal multilineage potential. This does not exclude the possibility that some environmental cues may induce a broader differentiation potential of tissue-specific stem cell lineages, since others have demonstrated that definitive neural stem cells and/or progenitor cells can give rise to non-neural cell types, such as endothelial cells or smooth muscle cells under specific conditions in the absence of cell fusion [40,41]. Nonetheless, our data suggest that the culture conditions likely do not induce a neurogenic potential in otherwise mesoderm-specified cells, but rather that these permissive conditions select for rare stem cells with multilineage potential.
Consistent with previous findings of extremely limited or no pluripotency of mouse epiblast-derived precursor cells beyond ∼8.0–8.5 dpc, clonally derived 9.5 dpc somite colonies do not express the pluripotency regulators Oct4 and Nanog, indicating that the neurogenic and myogenic capacity of these cells is not due to remnant pre-gastrula epiblast cells. Finally, our colony-forming assay does not appear to select for typical neural crest stem cells, since genetic lineage tracing of dorsal neural tube derivatives (Wnt1-Cre;Z/EG transgenics) as well as expression analyses of neural crest stem cell markers (Sox10 and p75) could not identify somite-derived colony-forming cells. However, we cannot rule out the possibility that the Sca1+/Sox2+/nestin+ colony-forming cells present in the somite compartment are a distinct subset of neural crest precursor, rather than an intrinsically mesodermal cell. Nonetheless, our data reveal a novel distinct population of multilineage stem cells resident within somite tissue that may represent the developmental predecessors of adult skeletal muscle multilineage stem cells.
Adult skeletal muscle interstitial precursor cells are a rare and relatively quiescent population that is multipotential (including neurogenic) in vitro and in vivo [42]. Furthermore, these adult nonsatellite skeletal muscle stem cells do not appear to express Pax3 and Pax7, but are instead CD45+/CD34+/Sca1+ [10,18,43], which distinguishes them from the Pax3+/Pax7+ skeletal muscle satellite stem cell lineage [reviewed in refs. 44 and 45]. Moreover, these cells are likely distinct from the MPC-like CD45+ hematopoietic cells that have been shown to have neurogenic and myogenic potential since they do not express the pluripotency markers Oct4 and Nanog [46]. Interestingly, our mRNA expression analyses indicate that somite-derived colonies are CD45+/CD34+/Sca1+ with relatively low Pax7 and Pax3 expression, suggesting the intriguing possibility that the developmental origin of the adult muscle interstitial precursor cell lies within a subpopulation of somite cells established as early as 9.5 dpc. This proposed embryonic origin of interstitial lineage specification is reminiscent of the recently characterized ontogeny of adult skeletal muscle satellite stem cells. Precursor cells from the dermomyotome compartment of the embryonic somite that express Pax3 and Pax7 were shown to give rise to the satellite stem cell population [34], which later retains Pax7 expression under quiescent conditions presumably throughout life. In contrast, Sca1 and CD45 markers are not expressed in adult satellite stem cells [47]. We speculate that the origin of the adult muscle interstitial stem cell-like lineage is established very early in development within the somites. Furthermore, it will be interesting to determine whether the 9.5 dpc neurogenic cardiac tissue precursors that we have also identified here share similar properties to the previously derived adult cardiac stem cells [48]. Regardless, further experiments are required to determine the precise localization of the Sca1+ cells within the various early post-gastrula mesodermal compartments and the molecular mechanisms guiding the ontogeny of these lineages as they establish resident adult stem cell subpopulations within mesodermal tissue derivatives.
Footnotes
Acknowledgments
We thank Freda Miller, Peter Zandstra, Ben Blencowe, and Derek van der Kooy for transgenic mice and reagents, and Stuart Rae for technical assistance. This research was supported by grants from CIHR and the Canadian Stem Cell Network to C.M.M., and the Canada Foundation for Innovation, Ontario Innovations Trust, and NSERC to V.T.