
Abstract
Previous studies have revealed c-kit-positive (c-kit+) cardiac stem cells (CSCs) in the adult mammalian heart and these cells could be a suitable cell source for heart regeneration therapy. However, these cells have not been fully evaluated in terms of characterization and effect of long-term culture, which is necessary for their safe and optimal usage. Therefore, we isolated c-kit+ CSCs from adult rat hearts to characterize these cells and investigate stability over long-term culture. We performed isolations of c-kit+ CSCs 11 times and passaged them 40 times in a bulk culture system; we termed these cultures, bulk culture CSCs (CSC-BC). c-kit+ CSCs expressed stemness genes and exhibited stem cell properties of single cell-derived clone formation, cardiosphere generation, and potential to differentiate into the three main cardiac lineages: cardiomyocyte, smooth muscle, and endothelial cells in vitro. Over long-term culture, some CSC-BC up-regulated GATA-4 expression, which resulted in enhanced cardiomyocyte differentiation, suggesting that the GATA-4 high c-kit+ CSCs have potent cardiac regenerative potential. We also observed the spontaneous differentiation into cells other than cardiac lineages, such as adipocyte and skeletal myocyte. This effect of long-term culture on the c-kit+ CSCs has not been previously reported. Interestingly, when c-kit+ CSCs were co-cultured with adult rat cardiomyocytes, we found increased cardiomyocyte survival, and the growth factors, insulin-like growth factor 1 (IGF-1) and vascular endothelial growth factor (VEGF), appeared to be responsible factors. The present study suggests that c-kit+ CSCs have great therapeutic potential yet should be further investigated and optimized as a cell source for regenerative therapies prior to transplantation.
Introduction
H
ES cells can efficiently generate both functional beating mature cardiomyocytes and vasculature; however, their use is hampered by associated teratoma formation and the need for an allogeneic source [8 –10]. iPS cells, like ES cells, are also a highly attractive means [11,12] but, their safety is still questioned due to retroviral gene transfer. Somatic stem cells include several types identified in adult tissues and have been used to reconstitute dead myocardium [13]. Among them, endothelial progenitor cells and bone marrow-derived cells have been the most widely used and recently tested in clinical trials [14 –18]. However, controversy surrounds the findings of bone marrow-derived cell differentiation into cardiomyocytes [19 –21] and several investigators are unable to replicate the early optimistic results from small animal models [22]. Furthermore, it has been suggested that the newly formed myocytes remain immature, do not acquire the adult phenotype, and die with time by apoptosis [23].
Until recently [24], the heart was originally thought to be entirely composed of terminally differentiated cardiomyocytes that withdrew from the cell cycle shortly after birth, and that cardiac injury caused permanent myocardial loss coupled with cardiac dysfunction [25,26]. However, several research groups have reported the isolation of cardiac stem-like cells from mice, rat, dog, and human hearts based on the cell surface antigens, stem cell antigen 1 (Sca-1 [27 –29]) Abcg2 [30 –32], and c-kit [6,33 –36]. Islet-1-positive cells were reported to be a distinct population of cardiac progenitors identified in embryonic and neonatal mouse and human hearts, but their existence in the adult heart is still unclear [37]. Apart from the islet-1-positive cells, many of the cardiac stem and progenitor cells identified thus far exhibit differences and similarities in phenotype (See Torella et al. 2008 [38] for review). For example, the Abcg2 or side population cells [30 –32] also express Sca-1 and c-kit, yet the latter is described as being low [30,31]. Furthermore, some of the isolated Sca-1-positive cells are also positive for c-kit [28] and likewise for the c-kit-positive (c-kit+) cells [33]. However, some Sca-1-positive cells are negative for c-kit [27]. These differences and similarities in phenotype bring into question whether they are all exclusively different or actually of the same population of cell yet selected and identified at different physiological states [38]. It could be that there exists a single cardiac stem cell (CSC) that expresses all of the markers identified depending on its developmental transition pathway [39]. Sca-1-positive cardiac stem/progenitor cells isolated from adult mice hearts can give rise to functionally beating cardiomyocytes in vitro [28] and following intravenous injection after ischemia reperfusion, Sca-1-positive cardiac stem/progenitor cells home to the injured mouse myocardium and differentiate into ∼1.5% new cardiomyocytes [27]. c-kit+ CSCs have been extensively examined, in terms of stem cell characteristics, and consistently exhibit several in vitro characteristics that define a “stem cell” [40]; being clonogenic, self-renewing, and multipotent [6]. Furthermore, when transplanted into infarcted rat hearts, c-kit+ CSCs differentiated into cardiomyocytes (∼14 × 106 new cardiomyocytes) and vascular structures, generating a large volume of new regenerated myocardium (48 mm3), which led to restoration of cardiac function [6]. These data have since been confirmed for c-kit+ CSCs from the human [41,42] and dog [35] heart. Therefore, the use of c-kit+ CSCs as an appropriate candidate cell for future cardiac regenerative medicine strategies is promising. For this purpose, we need to better understand how to generate large numbers of c-kit+ CSCs and the long-term consequence of the culture conditions on their spontaneous differentiation into different cell lineages and how this could impact on their use as a cell for regenerative therapy following myocardial injury. Therefore, we isolated c-kit+ CSCs from adult rat hearts, cultured them for over 40 passages (∼80 population doublings), and characterized them for markers of clonogenicity, multipotency, and differentiation.
Materials and Methods
Animal experiments
All animal experiments were performed according to Guidelines of Tokyo Women’s Medical University on Animal Use, The Principles of Laboratory Animal Care, formulated by the National Society for Medical Research, and Guide for the Care and Use of Laboratory Animals, prepared by the Institute of Laboratory Animal Resources and published by the National Institutes of Health (NIH Publication No. 86-23, revised 1985).
Isolation of c-kit+ cardiac cells and adult cardiomyocytes
c-kit+ cells were prepared by two protocols. Cardiac cells were isolated from adult Wistar, GFP-transgenic Wistar (GFP+, SLC; Shizuoka, Japan), or Lewis rats. Hearts were digested by perfusion with calcium-free Krebs-Henseleit buffer with collagenase, hyaluronidase, trypsin, and DNAse. The whole cell population was then divided into myocytes and small cells by centrifugation. Myocytes were discarded or used for the co-culture assays. When used for the co-culture assays, the myocytes were passed through a bovine serum albumin (BSA) size separation gradient that further separated and purified the myocyte population [7]. The small cells were treated with rabbit anti-c-kit primary antibody (Santa Cruz, Santa Cruz, CA). After rinsing, the cells were treated with goat anti-rabbit secondary antibody conjugated with magnetic beads (Miltenyi, Bergisch Gladbach, Germany). c-kit+ cells were isolated by magnetic cell sorting (MACS) (Miltenyi), and the purity of the isolation was assessed by FACS analysis.
For the second protocol and in order to better replicate what would happen in a clinical application, we used explant culture technique to obtain the small cells from pieces of myocardial tissues. One heart was separated into six parts, which consisted of left atrium, right atrium, left ventricle, right ventricle, septum, and apex. Then myocardial tissue from each part were cut into 1 to 2 mm3 pieces, washed with Ca2+–Mg2+-free phosphate-buffered solution (PBS) (Invitrogen, Carlsbad, CA), and digested three times for 5 min at 37°C with 0.2% trypsin (Invitrogen, Carlsbad, CA) and 0.1% collagenase (Sigma, St. Louis, MO). The suspended cells were discarded, and the remaining tissue fragments were washed with complete medium (Comp M) [Dulbecco’s modified Eagle’s medium [DMEM] supplemented with 10% ES cell grade fetal bovine serum, 5% horse serum, 10 ng/mL leukemia inhibitory factor (LIF; Chemicon, Millipore, Billerica, MA), 5 U/L erythropoietin (EPO; Sigma, St. Louis, MO), penicillin–streptomycin (Wako, Osaka, Japan), fungizone (Wako, Osaka, Japan), and gentamicin (Invitrogen)]. The cells were cultured in Comp M at 37°C in 5% CO2 incubator. After a period ranging from 1 to 2 weeks, attached cells, which were surrounding the explants having migrated out, were harvested and expanded. The tissue fragments were replated. These steps of cell harvest and replating the tissue were repeated for 3 months. The c-kit+ cells were isolated from these expanded cells using MACS as described earlier. With this method, the atria myocardial pieces yielded the highest amount of c-kit+ cells (9% ± 3% of total cells), and a left atria cell preparation represents bulk culture 5 (CSC-BC5) and was used for continued growth and analysis.
Primary culture of neonatal rat cardiomyocytes
Ventricles from 1-day-old Wistar rats (Nisseizai, Tokyo, Japan) were digested at 37°C in Hank’s balanced salt solution (Sigma, St. Louis, MO) containing collagenase (class II, Worthington Biochemical, Lakewood, NJ). Isolated cells were suspended in the culture medium comprising 6% FBS (Moregate Biotech, Bulimba, Australia), 40% Medium 199 (Invitrogen), 0.8% penicillin–streptomycin solution (Wako, Osaka, Japan), 2.7 mM glucose, and 54% balanced salt solution containing 116 mM NaCl, 1.0 mM NaH2PO4, 0.8 mM MgSO4, 1.18 mM KCl, 0.87 mM CaCl2, and 26.2 mM NaHCO3. Cell suspensions were plated in 35-mm dishes coated with laminin (1 mg/mL; Sigma, St. Louis, MO) and incubated at 37°C in a humidified condition.
Cell culture and cloning
c-kit+ cells were cultured with Comp M and the medium was replaced at 3- to 4-day intervals. Cultures of c-kit+ cells were passaged >40 times. At different passages, c-kit+ cells were seeded by serial dilution into single wells of 96-well plates at a density of 0.5 cell per well for the generation of single-cell clones. The colonies were expanded in sphere medium, modified from the previously reported NSCM [43] [Dulbecco’s MEM and Ham’s F12 (ratio 1:1; Sigma, St. Louis, MO), bFGF (10 ng/mL; Wako, Osaka, Japan), EGF (20 ng/mL; Wako, Osaka, Japan), LIF (10 ng/mL; Chemicon, Millipore, Billerica, MA), insulin–transferrin–selenite (Gibco or Sigma), B27 (Gibco, Invitrogen), N2 (Gibco, Invitrogen), penicillin–streptomycin (Wako, Osaka, Japan), fungizone (Wako, Osaka, Japan), and gentamicin (Gibco, Invitrogen)]. The clonogenicity of the c-kit+ cells was determined by counting the number of clones generated in each 96-well plate and expressed as a percentage. A total of 10 plates were analyzed. The medium for cardiomyocyte differentiation (CMDM) was composed of MEM Alpha (GIBCO, Invitrogen), 10% FBS, and supplemented with 1 µM dexamethasone (Sigma, St. Louis, MO), 50 µg/mL ascorbic acid (Sigma, St. Louis, MO), and 1 mM β-glycerophosphate (Sigma, St. Louis, MO) [44]. For inducing differentiation into adipocytes, osteoblasts, and skeletal myocytes, the cells were incubated in the following media, respectively. MEM/FBS containing 1 µM dexamethasone, 0.5 mM 3-isobutyl-1-methylxanthine, and 5 µg/mL insulin; MEM/FBS containing 1 µM dexamethasone, 50 µg/mL ascorbic acid, 10 mM β-glycerophosphate, and 10 nM 1-α,25-dihydroxyvitamin D3; and MEM/FBS containing 1.5 µg/mL amphotericin B (Wako, Osaka, Japan).
Immunofluorescence staining
Cultured cells were fixed with 4% formaldehyde for 20 min, permeabilized with 0.2% Triton-X 100 for 20 min, washed with PBS, blocked with 2% BSA (Sigma, St. Louis, MO) for 15 min, and then treated with antibodies against cardiac troponin I (cTnI; Santa Cruz, Santa Cruz, CA), smooth muscle actin (SMA; Sigma, St. Louis, MO), von Willebrand factor (vWF; Chemicon, Millipore, Billerica, MA), GATA-4 (Santa Cruz, Santa Cruz, CA), skeletal-type troponin I (Sigma, St. Louis, MO), and green fluorescent protein (GFP) (Molecular Probes, Invitrogen). After rinsing with PBS, the cells were treated with secondary antibodies, Alexa 488 or Alexa 594 (Molecular Probes, Invitrogen). Cells were all counterstained for nuclei detection with 4′,6-diamidino-2-phenylindole (DAPI; Invitrogen). Secondary antibody incubation alone was used as a negative control. Immunostaining was visualized and analyzed using epifluorescence (Zeiss, Oberkochen, Germany Axioplan2) or laser scanning confocal microscopy (Zeiss LSM510 META).
RT-PCR and quantitative real-time RT-PCR analysis
RNA was extracted from c-kit+ cells using ISOGEN (Wako, Osaka, Japan) or Nucleospin (Macherey-Nagel, Düren, Germany). RT-PCR was performed by two steps (Invitrogen), and cDNA was generated using a first strand cDNA Synthesis kit (Amersham, GE Healthcare, England). In brief, 5 µg total RNA was used as a template and random hexamer was used as a primer. cDNA was synthesized according to the manufacturer’s instructions. PCR was performed using the primers shown in Supplementary Table 1 (Supplementary materials are available online at http://www.liebertpub.com/). Cycle conditions were first, 94°C, 30 s; and then, 94°C, 15 s for denature, 60°C, 30 s for annealing, and 72°C, 30 s for extension. Cycle number was 25 at 72°C for 7 min until the end. RT-PCR or PCR products were run on 4% agarose/1× TBE gel (Reliant gel system, CAMBREX, Lonza, Valais, Switzerland). The expression levels were evaluated by the strength of the signal (the intensity of the band stained with ethidium bromide).
Real-time RT-PCR was performed either using SYBR (Qiagen, Hilden, Germany) or FAM-labeled probes (Applied Biosystems, Foster City, CA). PCR included 1 µL of template cDNA, and 250 nM forward and reverse primers. Cycling conditions were those as recommended by the manufacturer. SYBR or FAM signal was detected at the end of each cycle. The cycle program using SYBR was 50°C for 30 min, and then 95°C for 10 min, followed by 45 cycles of 94°C for 15 s, 60°C for 30 s, and 72°C for 40 s. The cycle program using FAM probe was 50°C for 30 min, and then 95°C for 10 min, followed by 45 cycles of 94°C for 15 s, 60°C for 1 min. Data were analyzed according to the instruction manual. PCR efficiency was evaluated by using a standard curve of five serial dilution points. Quantified values were normalized against the housekeeping gene, GAPDH.
Cardiomyocyte survival and ELISA analysis
Cloned cells from CSC-BC4 (CSCc4) and adult cardiomyocytes were co-cultured in a 24-well plate in CMDM. At the same time, CSCc4 were co-cultured with adult cardiomyocytes in CMDM using inserts (Corning Inc., Corning, NY) to separate contact between the CSCs and the myocytes. For the co-culture using inserts, CSCc4 (1 × 106 cells/insert) were cultured on the inserts and adult cardiomyocytes were cultured on the substrate. Adult cardiomyocytes were obtained from two rat hearts (2 × 104 cells/well). As control, CSCc4 (1 × 106 cells/well) alone or adult cardiomyocytes (2 × 104 cells/well) alone were cultured on a substrate. After culturing (both with inserts and without inserts) overnight, the medium was changed for fresh CMDM without FBS. Forty-eight hours later, 75% of the medium was harvested and fresh CMDM without FBS was applied. Ninety-six hours later, the medium was harvested and the cells on the substrate were fixed for immunostaining using cTnI antibody. To determine if specific growth factors contributed toward myocyte survival in vitro, ELISA assay was performed on the conditioned medium at 96 h from the co-culture assay. ELISA assay was performed using kits against specific growth factors, insulin-like growth factor 1 (IGF-1), TGF-β1, BMP-2 (R&D Systems, Minneapolis, MN), TNF-α (Pierce Biotech. Inc., Thermo Fisher Scientific, Rockford, IL), vascular endothelial growth factor (VEGF; RayBiotech, Inc., Norcross, GA), according to the manufacturer’s instruction.
Statistical analysis
Data are reported as mean ± SEM. Significance between multiple comparisons was evaluated by the analysis of variance (ANOVA). Bonferroni post hoc method was used to locate the differences. Significance was set at P < 0.05.
Results
c-kit+ cardiac cells can be extensively grown in vitro, are self-renewing, clonogenic, form cardiospheres, and differentiate into cardiomyocytes
c-kit+ cells were sorted by automacs and the purity of the cell sorting was assessed by FACS analysis. The c-kit+ cells isolated by automacs showed a purity of 95%, and almost 0% in a negative fraction (Fig. 1A), agreeing with previous reports [6,7]. When placed as a single cell into a 96-well plate, c-kit+ cardiac cells generated clones (Fig. 1B). The clonal efficiency of c-kit+ cardiac cells was 21% ± 7% (Fig. 1C). The remaining single c-kit+ cardiac cells, which did not form clones or colonies, most likely represent progenitor cells that have retained their expression of c-kit yet are more advanced in the differentiation transition pathway [39]. This is in line with a heterogeneous cell preparation containing cells that express both stemness and cardiac differentiation genes (see below). In addition, the non-clonal c-kit+ cardiac cells could be phenotypically similar cells but without stem cell potential (ie, cardiac mast cells [45]). When c-kit+ cells were placed in specific sphere-generation medium, spheres of cells formed (cardiospheres) (Fig. 1D). When the cardiospheres were plated on plastic and cells were allowed to migrate from the cardiosphere, they exhibited multipotency by differentiating into the three main cardiac lineages: cardiomyocyte, smooth muscle, and endothelial cells (Fig. 1E). The primary preparation of c-kit+ cardiac cells were analyzed by RT-PCR for expression of specific transcripts relevant to a stem cell phenotype and cardiac differentiation potential. As shown in Figure 1F, c-kit+ CSCs analyzed at passage 0 expressed the four genes known to be required for the induction of multipotency (Oct-4, Nanog, Klf-4, and Sox-2) and cardiac-specific differentiation genes; GATA-4, cTnI, SMA, and vWF. At passage 0, c-kit+ cardiac cells did not express the transcripts for the adipocyte (PPAR-α) or skeletal muscle lineage (myogenin) (Fig. 1F). These data show that c-kit+ cells isolated from the adult rat heart are a heterogeneous population, consisting of primitive and more committed cardiac progenitor and precursor cells. We thereafter termed these cells “cardiac stem cells” (CSCs). When cardiosphere-forming CSCs were treated with 10−8 M oxytocin for 1 month, all cells went onto express cTnI (Fig. 2A). However, these troponin I-positive cells did not reveal sarcomeric structure within the cytoplasm (Fig. 2A), and therefore cannot be considered as fully differentiated functional, mature cardiomyocytes, consistent with the previous article [6]. Next, we isolated c-kit+ CSCs from GFP-positive (GFP+) rats for a co-culture differentiation experiment with adult cardiomyocytes. As shown in Figure 2B, GFP+ c-kit+ CSCs differentiated into cTnI-positive cardiomyocytes (yellow arrowheads) when placed in co-culture with GFP-negative adult cardiomyocytes (white arrowheads) for 3 weeks. Furthermore, these cells had a defined sarcomeric structure, consistent with a more differentiated phenotype (Fig. 2B). The characteristics of c-kit+ CSCs isolated from the adult rat myocardium are similar to previous reports [6] and confirm the presence of an endogenous stem/progenitor cell in the adult mammalian heart.
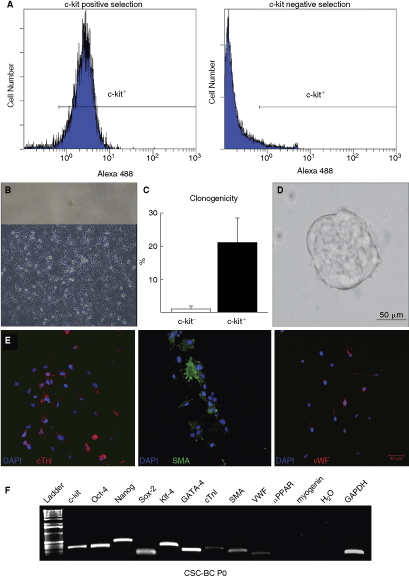
c-kit+ cardiac stem cells (CSCs) from the adult rat heart have stem cell characteristics. (
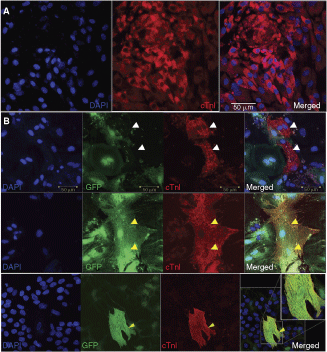
c-kit+ cardiac stem cells (CSCs) differentiate into the cardiomyogenic lineage in vitro. (
c-kit+ CSC bulk cultures (CSC-BC) exhibit variable expression of GATA-4, which determines cardiosphere formation and cardiac differentiation potential
For further characterization, especially in terms of stability over long-term culture, we grew 11 bulk cultures of c-kit+ CSCs (CSC-BC) and determined GATA-4 expression level by quantitative real-time RT-PCR (qRT-PCR) at different passages, up to passage 40. GATA-4 is a member of the GATA family of zinc finger transcription factors and is an early cardiomyocyte marker, playing an important role in transducing nuclear events that modulate cell lineage differentiation during development [46,47] and hypertrophy of adult cardiomyocytes [48]. The expression levels of GATA-4 in CSC-BC were variable (Fig. 3A). Most cultures showed a low and stable expression level of GATA-4 transcript over time, indicating that c-kit+ CSCs remained either primitive and undifferentiated or as committed progenitor cells throughout the culture period (Fig. 3A). In CSC-BC2 and 5, GATA-4 expression decreased between passages 20 and 24 (Fig. 3A). However, one particular bulk culture of c-kit+ CSCs (CSC-BC4) showed increased GATA-4 expression from passages 15 to 40 (Fig. 3A). The qRT-PCR results were confirmed by identification of the PCR products on an agarose gel where the observed band was the expected size as well as confirmed by the DNA sequence extracted from the band (data not shown). Immunofluorescence identified CSC-BC4 cells as expressing high levels of GATA-4 and CSC-BC1 cells as expressing low levels of GATA-4, at passage 40 (Fig. 3B). We then determined whether these GATA-4 high and low CSC-BC differed in terms of their ability to form cardiospheres and differentiate into the cardiomyocyte lineage. As shown in Figure 3C at passage 40, the GATA-4 high cardiac-committed CSC-BC4 showed decreased cardiosphere generation capability, compared to the low GATA-4 CSC-BC1. Moreover, CSC-BC4 exhibited a greater capacity to give rise to cells positive for cardiomyocyte-specific proteins, following migration from the cardiosphere (Fig. 3D and 3E).
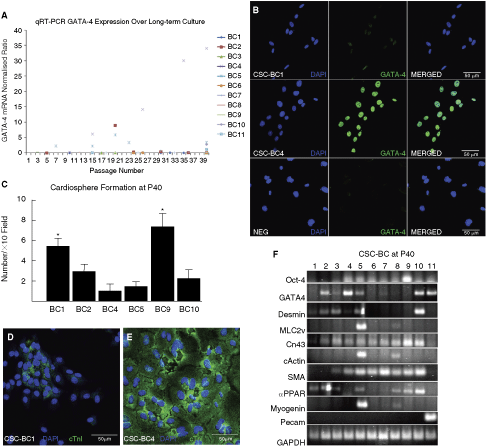
Long-term culture leads to changes in levels of GATA-4 in different bulk cultures of c-kit+ cardiac stem cells (CSCs; CSC-BC), which determines cardiosphere formation and cardiomyocyte differentiation potential. (
CSC-BCs consist of primitive stem cells and committed progenitor/precursor cells for the cardiac, adipocyte, and skeletal lineages
To determine whether CSC-BC showed commitment to lineages other than cardiac over culture time and because CSCs were reported to overcome cell senescence [6], we characterized the 11 CSC-BC after 40 passages for gene expression patterns of stemness (Oct-4), adipocyte (PPAR-α), skeletal muscle cells (myogenin, desmin), cardiomyocyte (GATA-4, MLC2v, cardiac actin (cActin), desmin, connexin43 [Cn43]), smooth muscle cell (SMA; smooth muscle actin), and endothelial cells (Pecam) by RT-PCR. The results of the RT-PCR are shown in Figure 3F and are summarized in Table 1. Three out of 11 CSC-BC revealed double positive expression of GATA-4/desmin. Seven out of 11 CSC-BC revealed adipocyte-specific gene expression, such as PPAR-α, and 2 out of 11 CSC-BC revealed skeletal myocyte gene expression, such as myogenin. Overall, we categorized three cell types within the CSC-BC according to their expression pattern: adipocyte (CSC-BC1, 2, 3, 9), mesenchymal/muscle-like (CSC-BC5, 8), and cardiac (CSC-BC4, 6, 7, 10, 11). All 11 CSC-BC retained cardiosphere formation ability at passage 40; however, some CSC-BC (1, 9) formed more cardiospheres than other CSC-BC (2, 4, 5, 10) (Fig. 3C). Moreover, these latter CSC-BC showed a high expression level of cardiac-specific differentiation genes (ie, GATA-4; MLC2v; cActin; Desmin; Fig. 3F and Table 1) and therefore had increased efficiency in giving rise to the cardiomyocyte lineage, compared to CSC-BC that did not express these transcripts. Similar to the findings earlier, concerning GATA-4 high (CSC-BC4) and low (CSC-BC1) CSC-BC, we also found that CSC-BC5, which was composed of mesenchymal/muscle and cardiac-committed CSCs (Fig. 3D and Table 1), exhibited decreased cardiosphere formation (Fig. 3C) and increased cardiomyocyte differentiation at passage 40 (data not shown).
CSC-BC indicates cardiac stem cell bulk-cultured cells 1–11.
Abbreviations: Adi, pre-adipocyte; Car, cardiac; Mes, mesenchymal. –, indicates no band observed; ±, very faint; +, faint; ++, visible; +++; clearly visible. MLC2v, myosin light chain 2v; Cn43, connexin43; cActin, cardiac actin; SMA, smooth muscle actin.
c-kit+ CSCs improve survival of adult cardiomyocytes in vitro
We showed that a co-culture system of c-kit+ CSCs with adult cardiomyocytes could induce CSC differentiation into cardiomyocytes (Fig. 2B). Interestingly, we also found that the presence of CSCs in the culture led to enhancement of adult rat cardiomyocyte survival, evidenced by an increased number of cardiomyocytes remaining attached and beating over time when cultured with CSCs (Fig. 4A). When the co-cultured cells were fixed and stained after 96 h, the number of survived troponin I-positive cardiac adult myocytes was increased (Fig. 4B). We then investigated whether different growth factors present in the conditioned culture medium could be associated with increased myocyte survival and number of beating myocytes. The protein level of IGF-1, TGF-β1, VEGF, TNF-α, and BMP-2 in the culture medium at 96 h after co-culture was assessed by ELISA. From all the factors analyzed, the concentration of IGF-1 and VEGF were significantly increased in the co-culture, using both inserts and without inserts (Fig. 4C and 4D). The enhancement for IGF-1 concentration following co-culture of myocytes and CSCs separated by inserts and without inserts was more than 6- and 2-fold higher, compared with myocytes alone, respectively (Fig. 4C). VEGF levels were also significantly increased following co-culture of myocytes with CSCs using inserts and without inserts (Fig. 4D), but the amount was not as high as IGF-1 concentration. The factors, TGF-β1, BMP-2, and TNF-α, were found at a very low level (data not shown). These findings are the first to show that c-kit+ CSCs have paracrine effects on cardiomyocytes as they increase myocyte survival in a co-culture in vitro experimental setting.

Co-culture of c-kit+ cardiac stem cells (CSCs) with adult rat cardiomyocytes leads to enhanced myocyte survival and increased IGF-1 and VEGF concentration in the co-culture medium. (
CSCs differentiate into adipocytes and skeletal muscle-like cells
We next determined if the CSC-BC that were composed of increased adipocyte (PPAR-α-positive) cells (CSC-BC1, 3, 9) and mesenchymal/muscle-like cells (myogenin-positive) (CSC-BC5, 8) (see Table 1 and Fig. 3F) could differentiate in vitro into those respective lineages. All of these CSC-BC differentiated into mature adipocytes, as shown by Oil Red staining (Fig. 5A), after incubation in adipocyte differentiation medium for 1 week. Some cells within CSC-BC5 showed a change in morphology toward an elongated shape at passage 39 (Fig. 5B and 5C). When further grown in skeletal myocyte differentiation medium and analyzed by real-time RT-PCR, these cells were GATA-4- (Fig. 5D) and MEF2C-positive (Fig. 5E), and went onto express skeletal-type troponin I (Fig. 5F), suggesting that they differentiated into skeletal muscle cells.
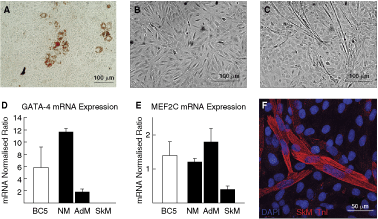
c-kit+ cardiac stem cells (CSCs) differentiate into adipocytes and skeletal muscle cells. (
Discussion
The results of the present study confirm that c-kit+ CSCs exist in the adult rat heart, which is in line with previous data [6,7]. We show here that c-kit+ CSCs can be expanded through growth in culture beyond the population doubling limit of somatic cells (>40). These cells are clonogenic, express multipotency and stemness genes, and form cardiospheres (markers of clonogenicity and multipotency, and can be used for isolation of stem cells from tissue culture [49]). The cells that migrate from the sphere spontaneously express protein markers for the cardiomyocyte, smooth muscle, and endothelial lineage. When placed in a specific differentiation medium together with adult rat cardiomyocytes, c-kit+ CSCs differentiated into cardiomyocytes, expressing cTnI with cytoplasmic sarcomeric structure (Fig. 2B). Further analysis is needed on the c-kit+ cardiac cell population to determine the presence of CD markers such as CD45 and CD34, and to distinguish whether the presence of these markers on c-kit+ cardiac cells enables them to also behave like CSCs [45]. The present study also confirms that a high purity of c-kit+ CSCs can be isolated from the adult rat heart by AUTOMACS technology. The present findings show that bulk-cultured uncloned cardiac-derived c-kit+ CSCs can be grown for over 40 passages and maintain a cardiac phenotype. Most importantly, we show the expression of GATA-4 can change over culture time and that c-kit+ CSCs that are GATA-4 high, and therefore have committed to the cardiomyocyte lineage, are less efficient at generating cardiospheres (which is probably representative of decreased clonogencity and multipotency) but show increased capacity to differentiate into cardiomyocytes, compared to GATA-4 low CSCs (Fig. 3C–3E). These findings have relevance for future regenerative cardiovascular protocols, in that c-kit+ CSCs could be driven or manipulated to increase GATA-4 expression before transplantation in order to increase cardiomyocyte differentiation. Therefore, the GATA-4 high population of CSCs could represent the most potent population to regenerate the cardiomyocyte population in the injured heart. Furthermore, analysis is warranted to determine the in vivo regenerative potential of these cells.
We showed that when c-kit+ CSCs are grown in bulk culture they spontaneously differentiated into not only cardiac-like cells, but also muscle- and adipocyte-like cells, expressing markers and transcription factors for these lineages. These results suggest that c-kit+ CSCs are a heterogeneous population within the heart consisting of primitive stem cells and more committed cardiac progenitor and precursor cells, and that c-kit+ CSCs change their phenotype with long time culture [50] with commitment to lineages other than cardiac. This is consolidated through the absence of transcripts for adipocyte (PPAR-α) and skeletal muscle (myogenin) differentiation in c-kit+ CSCs at passage 0 (Fig. 1F). Interestingly, CSCs enhanced the survival of adult rat cardiomyocytes when placed in a co-culture system. This was evidenced by sustained contraction and increased number of attached adult cardiomyocytes. This effect was not evident when adult rat cardiomyocytes were cultured without CSCs or when cultured with fibroblasts (data not shown). We showed that the levels of IGF-1 and VEGF were the specific growth factors that were elevated in the co-culture media and could therefore be associated with increased cardiomyocyte survival (Fig. 4C and 4D). These data suggest that increased numbers of CSCs could have a beneficial survival effect, due to IGF-1 and VEGF secretion, which could explain the recovery of cardiomyocyte performance in vivo [51 –53]. Indeed, bone marrow-derived cells exert a beneficial effect on cardiac function upon administration to the human ischemic myocardium [14]. The mode of action does not seem to involve differentiation into cardiomyocytes and vascular cells. A robust effect on revascularization and remodeling is observed; however, this is most likely mediated by paracrine factors [54,55]. The level of IGF-1 concentration in the culture medium when CSCs were cultured alone was extremely low (Fig. 4C), suggesting that CSCs are not responsible for IGF-1 release per se yet it is another effect of the combination of myocytes and CSCs in co-culture, which leads to an increase in IGF-1 expression. It could be that IGF-1 is secreted in high concentration by cardiomyocytes. However, as the level of IGF-1 expression was also low when myocytes were cultured alone (Fig. 4C), it could be that the CSCs mediate the release of other paracrine factors and molecules, which in turn drive the release of IGF-1 expression from the CSCs and myocytes, leading to improved cardiomyocyte survival through an auto/paracrine mechanism. VEGF has been reported to have a role for vascularization in in vivo regeneration [56,57] and myocyte contraction in vitro [57]. In the present study VEGF was up-regulated in the medium following co-culture of CSCs with myocytes; however, its level of expression was not as high as IGF-1. Nevertheless, these results suggest that VEGF may have a role in CSC paracrine-mediated cardiomyocyte survival.
Recently identified drawbacks associated with cardiac mesenchymal stem cell therapy include differentiation into unwanted mesenchymal cell types such as osteocytes and adipocytes, the occurrence of cytogenetic instability upon prolonged expansion, and immunization when used in an allogeneic setting. The present findings show that c-kit+ CSCs spontaneously differentiate into adipocyte-, muscle-, and cardiac-like cells over long-term culture. Furthermore, when these pre-adipocyte and pre-muscle-like cells were placed in their respective specific differentiation medium they went onto mature into that phenotype. The underpinnings of differentiation into cell types other than cardiac during culture conditions need to be investigated further in order to avoid unwanted differentiation before cell transplantation for myocardial regenerative therapies in vivo. This apparent increase in the multipotency of c-kit+ CSCs after long-term culture could result from either a change in phenotype in response to culture conditions [58] or, more likely, the preferential growth and survival in culture of a reduced number of cells with a broader developmental potential present in the adult heart and, therefore, in the primary cell population. In conclusion, our results indicate that c-kit+ CSCs have stem cell characteristics and the capacity for differentiation into cardiomyocytes, and can release specific factors to maintain cardiomyocyte survival in vitro. We also show that committed c-kit+ cardiac progenitor cells, which express high levels of GATA-4, have increased capacity to give rise to cardiomyocyte-specific progeny. Over long-term culture, we found the unwanted differentiation of c-kit+ CSCs into adipocytes and skeletal muscle cells, which could potentially confound the effectiveness of future regenerative protocols using c-kit+ CSCs. Further characterization of these cells is needed before the efficient usage of CSCs for regenerative medicine therapies.
Footnotes
Acknowledgments
The authors thank Dr. Kamrul M. Hasan for technical help, and Drs. Bernardo Nadal-Ginard and Goki Matsumura for advice. This project was supported by the encouraging Development of Strategic Research Centers, Special Coordination Funds for Promoting Science and Technology, Ministry of Education, Culture, Sports, Science and Technology, Japan, and a Grant-in Aid for Scientific Research (A) (Japan). G.M.E. is the recipient of a Marie Curie International Reintegration Grant within the 7th European Community Framework Programme (PIRG02-GA-2007-224853).
Author Disclosure Statement
Authors have nothing to disclose and declare no financial conflict of interest.