
Abstract
Members of helix-loop-helix (HLH) protein family of Id (inhibitor of differentiation) dimerize with bHLH transcription factors and function as negative regulators of differentiation during development. Most of inhibitory roles of Id proteins have been demonstrated in non-neural tissues, and their roles in the developing nervous system are not clearly demonstrated. In this study, we show that Id1, Id2, and Id3 increase self-renewing and proliferation potential of cortical neural stem cells (NSCs) while inhibiting neuronal differentiation. In electrophoretic mobility gel shift and luciferase assays, Id proteins interfered with binding of NeuroD/E47 complexes to the E-box sequences and inhibited E-box-mediated gene expression. Overexpression of Id proteins in NSCs increased both the number and the size of neurospheres in colony-forming assays. Expression of Hes1 and Hes5 was not increased by overexpression of Id proteins under the condition in which Nestin expression was increased. In utero electroporation of Id yielded higher numbers of Ki67-positive and Sox2-positive cells in the mouse embryonic brain. The study suggests Id proteins play independent roles in the maintenance of neural stem properties.
Introduction
M
Interference with bHLH protein to form functional complex leads to inhibition of premature neuronal differentiation. Members of the Hes gene family, particularly Hesl and Hes5, associate with Mashl and inhibit Mashl's binding to its target genes [6,7]. Homodimers of Hes proteins can also directly bind to the N-box sequences (CACNAG) in the Mashl promoter and inhibit Mashl transcription by recruiting a co-repressor such as Groucho/TLE [8]. These negative regulatory functions of Hes proteins are critical for sustained proliferation of neural precursor cells. Hes1, Hes3, and Hes5 are expressed in neural precursors of the ventricular zone of developing telencephalon [6,7,9]. Consistently, gene targeting of both Hes1 and Hes5 causes premature differentiation of neural precursor cells [10 –12] and disorganizes the cyto-architecture of the brain [13].
Like Hes1, Id also exerts antagonistic functions against bHLH transcription factors. Most of the mechanistic studies on Id proteins have been carried out in non-neuronal tissues. Id1 and Id2 protein inhibit E-box-mediated gene expression by forming nonfunctional complexes with myogenic bHLH factors such as MyoD and myogenin during muscle differentiation [14,15]. In the process of lymphopoiesis, Id2 or Id3 protein interacts with ubiquitous bHLH factors such as E-proteins and inhibits lymphocyte differentiation [16,17]. Additionally, Id proteins can promote cell cycle progression by sequestering inhibitors of cell cycle progression (see Discussion). Interestingly, targeted mutations of Id1 in conjunction with Id3, or Id4, produce a small brain phenotype [18 –20], suggesting that Id proteins may play critical roles in the cell cycle regulation of neural precursor cells. Surprisingly, little progress has since been made with regards to understanding the molecular functions of Id proteins during neurogenesis.
In this study, using neural stem cells (NSCs) isolated from the developing mouse telencephalon, we show that Id proteins play dual roles at molecular levels. First, Id proteins maintain the self-renewal potential and promote proliferation of NSCs. Second, Id proteins interfere with formation of functional proneural bHLH complexes and thereby inhibiting neuronal differentiation while stimulating astrocyte differentiation.
Materials and Methods
Plasmids
Full-length cDNAs of Id proteins were isolated by yeast 2 hybrid screening as previously described [21]. The sequences for Id cDNAs were identical to the previously published sequences: Id1 (GenBank number M31885), Id2 (GenBank number BC006921), and Id3 (GenBank number NM_008321.1). Three different types of mammalian expression plasmids for Id proteins were generated. For pcDNA3.1-Id, the full-length cDNA with FLAG epitope at the N-terminus was inserted to pcDNA3.1 (Invitrogen, Carlsbad, CA); for pMSCV-Id, the FLAG-Id cDNA fragment into the pMSCV-puro (Clonetech, Palo Alto, CA); and for Id-epidermal growth factor (EGF)P-CLBC3 the Id cDNA was inserted in front of the internal ribosome entry site (IRES) sequence [22] and enhanced GFP sequence. Expression vectors for NeuroD, pcDNA3.1-NeuroD, and for E47 (E2A gene product), pSVE2–5, were previously described [23].
Cell culture and transfection assay
293T and P19 cells were maintained in Dulbecco's modified Eagle's medium (DMEM) with high glucose supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 mg/mL streptomycin (growth medium) (Invitrogen, Carlsbad, CA) at 37°C in a humidified 5% CO2 atmosphere. Retroviral vectors were produced using a standard method by co-transfecting 293T cells with pMSCV-puro/Id or pMSCV-puro/β-galactosidase and gag- and env-expression vectors. Forty-eight hours later, the viral supernatant was collected and filtered through a 0.45 μm filter.
P19 cells were transfected with pcDNA3.1-Id using LipofectAMINE PLUS™ (Invitrogen) and selected in the presence of 400 μg/mL G418 (Invitrogen). Neuronal differentiation of P19-driven cells was induced by culturing 2 × 106 cells as aggregates in a Petri dish for 2 days in the presence of 0.5 μM all-trans retinoic acid (RA) and for additional 2 days in the absence of RA as previously described [24,25]. The aggregates were plated onto tissue culture dishes coated with laminin and poly-
Immunocytochemistry
Cells were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) and incubated for 1 h at room temperature in a blocking solution (0.1% bovine serum albumin, 10% normal horse serum, 0.1% Triton X-100 in PBS). After blocking, the cells were incubated in 0.1% Triton X-100 in PBS containing primary antibodies against β-tubulin III (BAbCO, Richmond, CA) (1:1,000), glial fibrilliary acidic protein (GFAP) (Sigma-Aldrich) (1:1,000), or Nestin (Chemicon, Temecula, CA) at 4°C overnight. Antibody reaction was visualized with Alexa Fluor 488 or 566 antibodies (Molecular Probes, Eugene, OR), and the cells were counterstained with Hoechst (Molecular Probes).
Luciferase assay
HIT cells were grown in the same media used for P19 cells. For luciferase assay, the cells were transfected with 0.5 μg pcHA-N233 or 0.1, 0.3, and 1 μg of pcDNA3.1-Id plasmids using LipofectAMINE PLUS™. Thirty-six hours later, cell extracts were prepared according to the manufacturer's protocol, and luciferase activity was determined with 5–20 μg proteins of cell extracts using the Dual-Luciferase assay system (Promega, Madison, WI). All the transfections were carried out with 0.1 μg pCMV β-gal. Luciferase activity was normalized for the transfection efficiency with respect to the β-galactosidase activity.
Neural stem cells
Mouse forebrain was dissected from the 12.5-day embryos triturated in Accutase (Cell Technologies, San Diego, CA) to single cells using a micropipette. After washing with HBSS, the cells were plated at a density of 1 × 105 cells/mL in DMEM:F12 (1:1) with N2 supplement (Invitrogen), 2 μg/mL heparin sulfate, 20 ng/mL EGF (Sigma-Aldrich), 20 ng/mL bFGF (Dong-a Pharmaceutical Research Lab, Yongin, Korea). To induce neuronal differentiation, growth factors were gradually eliminated from the media by replacing a half of the medium with new medium without growth factors every other day. To obtain Id-overexpressing NSCs, we first isolated primary neural cells from E12.5 forebrain and cultured as neurospheres for 7 days until they form small clusters composed of 20–40 cells. Then, we transduced the cells with pMSCV-Id or pMSCV-LacZ retroviral vector in the presence of 8 μg/mL polybrene (Sigma-Aldrich) for 6 h. The cells were incubated in the presence of 2 μg/mL puromycin (Sigma-Aldrich) for 14 days to form primary neurospheres. Since free-floating neurospheres are highly motile and prone to fuse [26], we utilized 1.16 mg/mL bovine collagen (type I:type III = 95∼98:5∼2) in final culture medium to eliminate mobilization and fusion of neurospheres in our clonal analyses. In brief, 1 × 104 cells were plated in bFGF- and EGF-containing media using NeuroCult neural colony-forming cell assay kit following the manufacturer's instruction (Stem Cell Technology, Vancouver, BC, Canada). For secondary neurosphere-forming assay, cells in primary neurospheres were dissociated in Accutase mechanically by repeated pipetting and finally plated 1 × 104 cells in the bFGF- and EGF-containing media.
Electrophoretic mobility supershift assay (EMSA)
293T cells were co-transfected with 4 μg pcDNA3.1-NeuroD, 2 μg pSVE2–5, and 4–8 μg each of pcDNA3.1-Id1–3. After 36-h incubation, nuclear extracts were prepared from transfected cells as described [23]. One microgram of nuclear extracts was incubated with 3 × 104 cpm of 32 P-labeled double-stranded oligonucleotide containing 3 copies of E-box of the rat insulin promoter (RIPE3) in 7% glycerol, 60 mM LiCl, 0.5 mM phenylmethylsulfonyl fluoride, 5 mM MgCl2, 2 mM dithiothreitol, 10 mM Tris-HCl, pH 7.4, and 2 μg of poly dI/dC for 30 min at room temperature as described [23]. To confirm specific binding, the unlabeled probe of 30- to 100-fold excess or 0.6 μg of an anti-NeuroD antibody (Santa Cruz Biotech, Santa Cruz, CA) was added to the reaction mixture. Samples were loaded onto the 6% polyacrylamide gels and subjected to electrophoresis at 8 V/cm. Gels were dried and autoradiography was performed.
Western analysis
Cells were lysed in 15 mM NaCl, 1% NP-40, 0.5% sodium deoxycholate, 0.1% SDS, 50 mM Tris, pH 8.0. After centrifugation at 10,000g for 5 min, the supernatant was harvested and 40 μg proteins were separated on SDS-polyacrylamide gel electrophoresis. The proteins on the gel were electrophoretically transferred onto PVDF membrane (Millipore, Bedford, UK) and probed with an anti-FLAG antibody (Sigma-Aldrich) (1:1,000). Finally, the membrane was incubated with horseradish peroxidase-conjugated anti-mouse IgG antibody (Vector, Burlingame, CA). The proteins were visualized using SuperSignal enhanced chemiluminescent kit (PIERCE, Rockford, IL) following the manufacturer's recommendation.
Reverse transcriptase-polymerase chain reaction (RT-PCR)
Total RNAs from neurospheres and differentiated cells were isolated using a RNAzolB (TEL-TEST, County Road, TX), and cDNA was synthesized using 1 μg of total RNA using first-strand cDNA synthesis kit following the manufacturer's instructions (Roche, Mannheim, Germany). PCR was conducted with 3 μL of the first-strand cDNA products. Primers were designed to recognize separate exons in order to distinguish products from genomic DNA contamination (Table 1). PCR was conducted with activation at 98°C for 1 min, followed by 25 cycles of denaturation at 94°C for 1 min, annealing at the indicated temperatures for 30 s (Table 1), and extension at 72°C for 1 min. Final extension was performed at 72°C for 7 min. PCR products were separated on 2% agarose gels. The experiments were performed 3 times and the numbers of spheres with various diameters are presented as means ± S.E.
In utero electroporation and immunohistochemistry
Pregnant ICR mice with 13.5 days of gestation were anesthetized with intraperitoneal injection of 50 mg/kg of ketamine (Yuhan, Seoul, Korea) and 2.5 mg/kg of rumpun (Beyer Healthcare, Seoul, Korea). DNA in PBS (4 μg/pL) containing 0.05% fast green was injected into the lateral ventricle of the embryonic forebrain using a 90-mm glass capillary (GD-1; Narishige, Tokyo, Japan). Electroporation was performed with a tweezer type electrode with 5 pulses of 40 V for 50 ms duration and 950 ms interval using a ECM 830 pulse generator (BTX, San Diego, CA) as previously described [27]. On 15.5 day of gestation, the GFP-expressing embryos were removed from uteri, fixed with 4% neutralized paraformaldehyde at 4°C for 2 h, and then further incubated in 30% sucrose in PBS overnight at 4°C. The embryo was embedded in tissue-freezing medium (Leica, Deerfield, IL), serially sectioned in a 10-μm thickness using a CM3050 S cryostat (Leica), and mounted on the silane-coated glass slides. The sections were incubated with primary antibodies (1:200) against GFP (Abcam, Cambridge, MA), Ki67 (Chemicon), or Sox2 (Abcam, Cambridge, UK) overnight at 4°C and then incubated in Alexa 488 or 568 conjugated secondary antibodies (Molecular Probe) (1:500) for 1 h at room temperature. After washing, the specimens were mounted onto coverslips using Vectashield (Vector Labs., Burlingame, CA). The fluorescent images were acquired with a LSM 510 confocal microscope (Zeiss, Oberkochen, Germany).
Statistical analysis
The data are presented as means ± S.E from at least 3 independent experiments. P values for statistical significance were estimated from t-test with respect to the indicated control value (*P < 0.05, **P < 0.01).
Results
Id expression correlates with stemness of NSCs
To determine the role of Id proteins, we first investigated the expression levels of Id1–3 during neuronal differentiation. NSCs were isolated from the E12.5 embryonic forebrain and grown as neurospheres in the presence of bFGF and EGF (Fig. 1A). The cells in neurospheres expressed nestin, a neural precursor cell marker (Fig. 1B). When growth factors were gradually removed from the culture, these cells spontaneously differentiated into several lineages of neural cells that were positive for neurofilament H (NF-H), a terminally differentiated neuronal marker, or for GFAP, an astrocyte marker (Fig. 1C and 1D). RT-PCR analysis showed that Id1, Id2, and Id3 were highly expressed in neurospheres but decreased in differentiated cells (Fig. 1E and 1F). These data indicate that expression of Id protein is tightly correlated to the proliferative stage of NSCs.

Expression of inhibitor of differentiation (Id) correlates with proliferation status of neural stem cells (NSCs). (
Id proteins promote self-renewal of NSCs
To test the effects of Id proteins in NSCs, we first generated Id-expressing NSCs by transducing with retroviral vectors encoding each of Id1–3. The cells stably expressing Id proteins were plated at a low density (0.5 × 104 cells/mL) in collagen gel and grown in the presence of bFGF and EGF for 7 days. The collagen gel prohibited random aggregation of cells and allowed us to detect the self-renewal potential and proliferation of seeded cells. Overexpression of Id proteins in NSCs increased the total numbers of neurospheres by 1.8-fold (Fig. 2A and 2E). Specifically, while NSCs overexpressing LacZ as a negative control yielded 80 ± 4 neurospheres, NSCs overexpressing Id1, Id2, and Id3 yielded 162 ± 29, 150 ± 8, and 161 ± 15 neurospheres, respectively (Fig. 2E). We also noted that there was a proportionately higher number of neurospheres that attained large size of 51–100 μm in diameter from Id-transduced cells. Neurospheres with a diameter of 51–100 μm was less than 39 ± 3 for NSCs with LacZ, whereas NSCs with Id proteins yielded 76–84 neurospheres of this size (Fig. 2E). To further verify the self-renewal potential of the neurosphere-forming cells, the neurospheres grown in the collagen gel were dissociated and subjected to secondary sphere forming assay for 7 days. Again, Id increased both the total numbers and sizes of neurospheres by 3–5-folds compared to LacZ (Fig. 2F). These data suggest that the Id proteins accelerate proliferation rates (shown by the increase diameters) and self-renewal potential of NSCs (shown by the increase in the total numbers of neurospheres). RT-PCR analysis showed that overexpression of Id1–3 enhanced the expression of nestin by 2.5-fold under the undifferentiating condition in the presence of growth factors (Fig. 2C). Under the same condition, expression of Hes1 and Hes5 was not altered. These data suggest that Id proteins prohibit spontaneous differentiation of NSCs while preserving NSC properties via a pathway independent of Hes genes. It is notable that overexpression of Id proteins reduced the NF-H (Fig. 2D) while increasing the GFAP in differentiated cells (Fig. 2B and 2D). These data suggest that the Id proteins do not simply inhibit differentiation by inhibiting cell cycle exit, but rather specifically inhibit neuronal differentiation while promoting astroglial differentiation. This result in turn suggested the specific functions of Id proteins via regulation of proneural bHLH transcription factors.
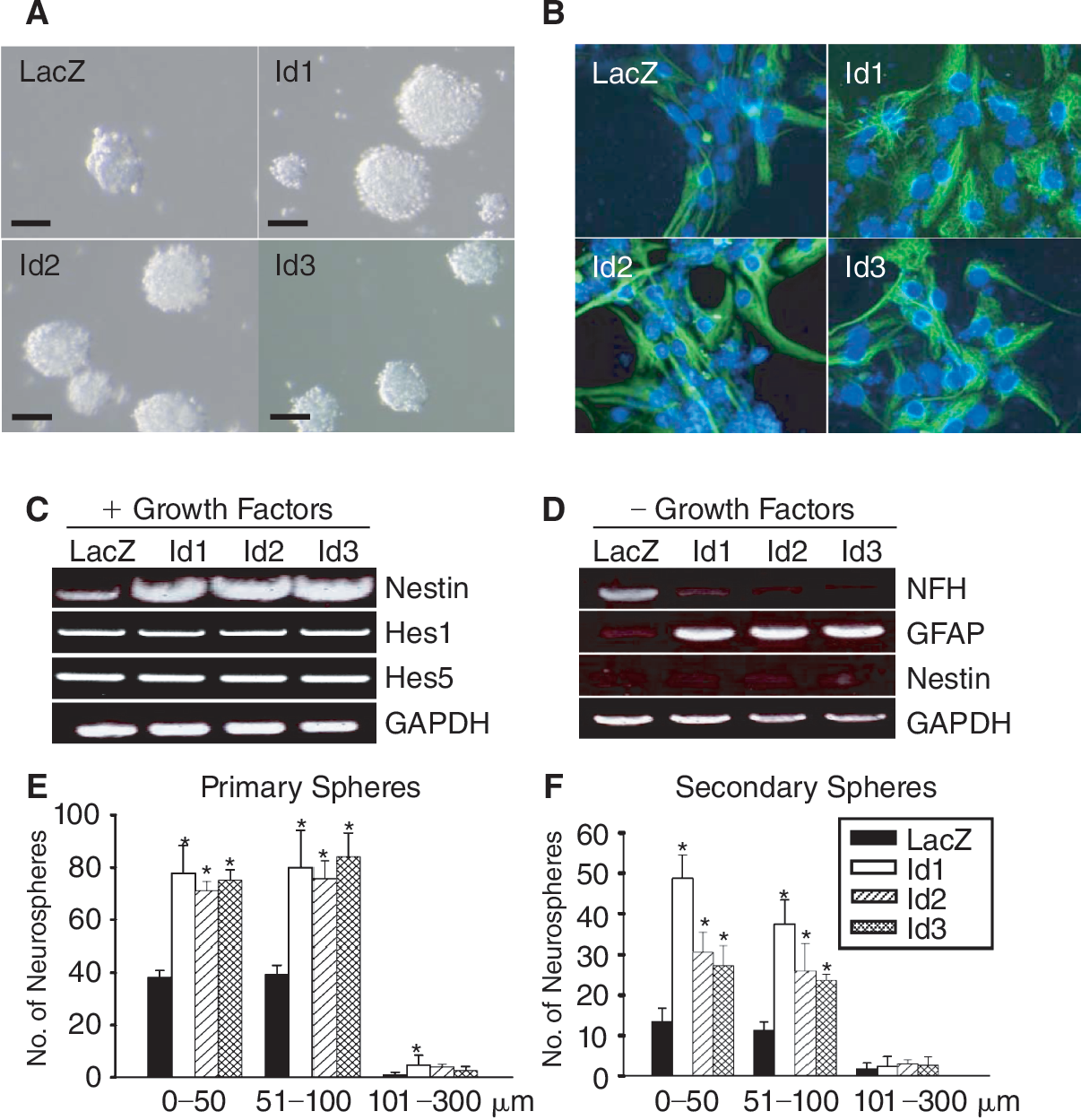
Id (inhibitor of differentiation) proteins potentiate stem cell properties of neural stem cells (NSCs). (
Inhibition of E-box-mediated gene expression by Id proteins
To determine whether the inhibition of neuronal differentiation is due to Id's antagonistic functions against proneural bHLH proteins, we used hamster insulinoma tumor cells (HIT cell). HIT cells express endogenous NeuroD and provide an in vitro system to measure E-box-mediated gene expression [23]. We co-transfected the cells with increasing amounts of expression vectors for Id1–3 and a luciferase reporter gene driven by 3 copies of E-box sequences of the rat insulin gene (RIPE3-luc) [28]. Due to the expression of endogenous NeuroD/BETA2, the basal expression of the reporter gene was high in HIT cells (Fig. 3B). Overexpression of Id1–3 reduced the luciferase activity in a dose-dependent manner. Similarly, N233, a dominant negative mutant of NeuroD containing the DNA-binding domain but lacking the activation domain, also inhibited the NeuroD-mediated transactivation of the RIPE3 reporter gene. The data suggest that Id proteins can counteract the E-box activation driven by proneural bHLH transcription factors. We also found that the inhibition of E-box-mediated transcription by Id proteins was correlated with reduced neuronal differentiation. P19 embryonal carcinoma cells differentiated into β-tubulin III-positive neuron-like cells in the absence of growth factor stimuli, and overexpression of Id proteins reduced the number of neuron-like cells in P19 cells (Fig. 3D and 3E).
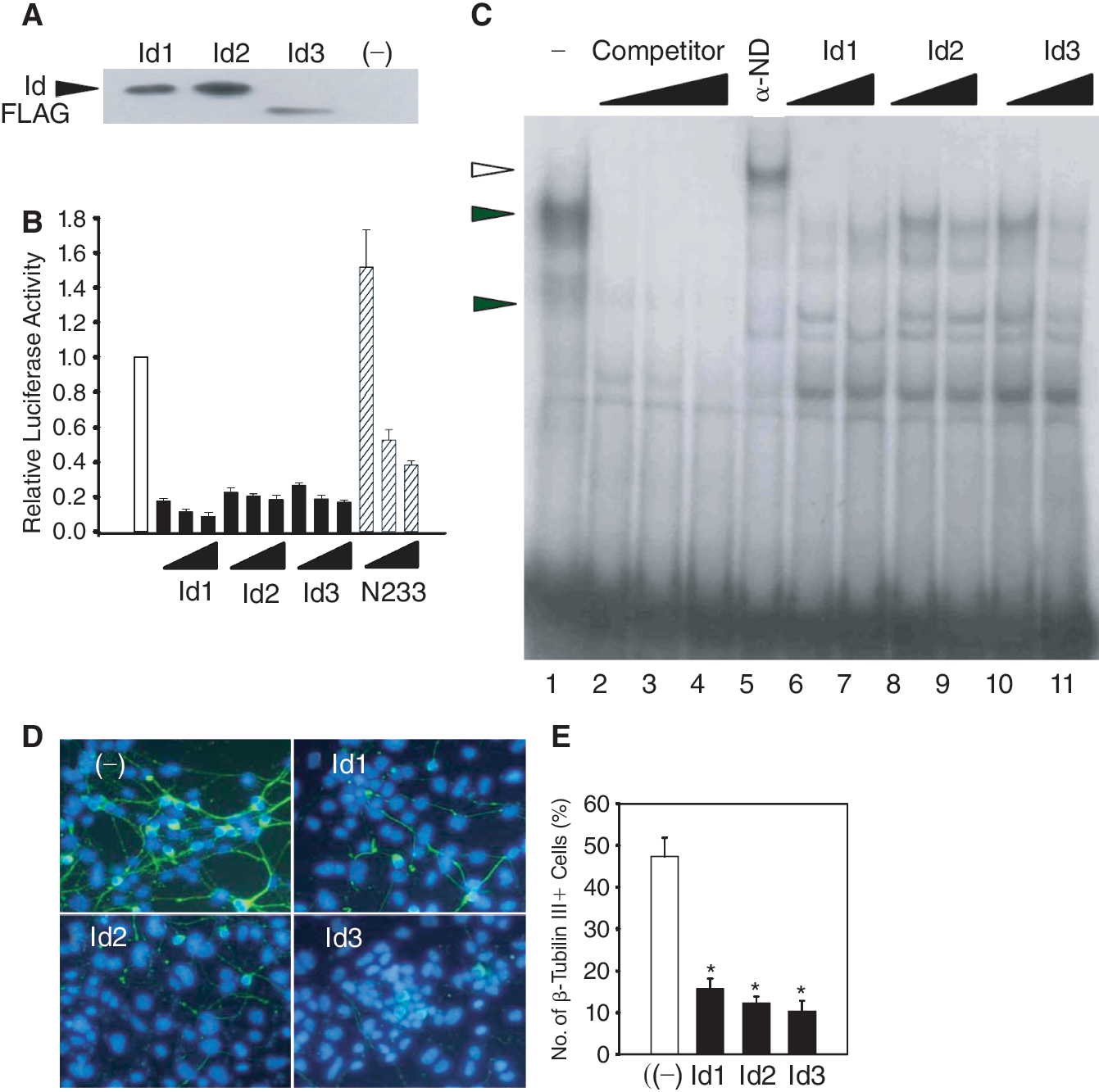
Inhibition of E-box-mediated gene expression by inhibitor of differentiation (Id) proteins. (
To further demonstrate that the Id proteins function by reducing the binding of bHLH complexes to the E-box sequences, we carried out electrophoretic mobility shift assay with RIPE3 as a probe. We obtained the functional complexes of NeuroD::E47 by co-transfecting 293T cells with their expression vectors. Heterodimers of NeuroD::E47 yielded specific complexes (closed triangles in lane 1, Fig. 3C) that could be supershifted with an anti-NeuroD antibody (an open triangle, lane 5). Consistent with that nonfunctional heterodimers containing Id proteins being unable to bind E-box sequences, this complex diminished in the presence of excess amount of Id proteins (lanes 6–11). The results indicate that the inhibitory effects of Id proteins on neuronal differentiation can be attributed to formation of nonfunctional heterodimers of Id and bHLH factors.
Id proteins increase the proliferating neural precursors in the developing brain
To confirm the roles of Id genes in neural precursor cells in vivo, we overexpressed Id proteins in the developing forebrain by in utero electroporation on embryonic day 13.5 (E13.5). Id expression vectors, Id-EGFP-CLBC3, contained the internal ribosome entry site (IRES) sequence [22] in front of the GFP sequence and allowed us to identify the transfected cells. At E15.5, we removed the embryonic brain expressing GFP (Fig. 4A–4C) and immunostained for the expression of Sox2, a NSC marker [29,30] or Ki67, a proliferation marker [31]. The number of Ki67-positive cells in the hemispheres electroporated with Id-EGFP-CLBC3 plasmids were increased in VZ and SVZ; 13% ± 2.9% for Id1, 17% ± 2.1% for Id2, 13% ± 2.8% for Id3 (Fig. 4I–4K and 4L); in the intermediate zone (IZ) 5% ± 1.7% for Id1, 5% ± 2.1% for Id2, 5% ± 2.9% for Id3 (Fig. 4E– and 4L). In contrast, Ki67-positive cells were not found in IZ of the control animal brain (Fig. 4D and 4L). In the ventricular zone (VZ), most cells expressed Sox2 (Fig. 5I and 5M), and the expression of Id proteins did not further increase the Sox2 immunoreactivity (Fig. 5J–5L and 5M). Overexpression of Id proteins increased the number of Sox2-positive cells in the subventricular zone (SVZ). The number of Sox2-positive cells among transfected cells was 14% ± 11.3% for Id1, 26% ± 10.7 % for Id2, and 10% ± 0.5% for Id3 (Fig. 5F–5H). Such Sox2-positive cells were not found in the SVZ of control embryos that were electroporated with LacZ-IRES-GFP (Fig. 5E and 5M). Taken together, these results demonstrate that Id proteins induce proliferation of NSCs in the developing brain as well as in NSCs in vitro.
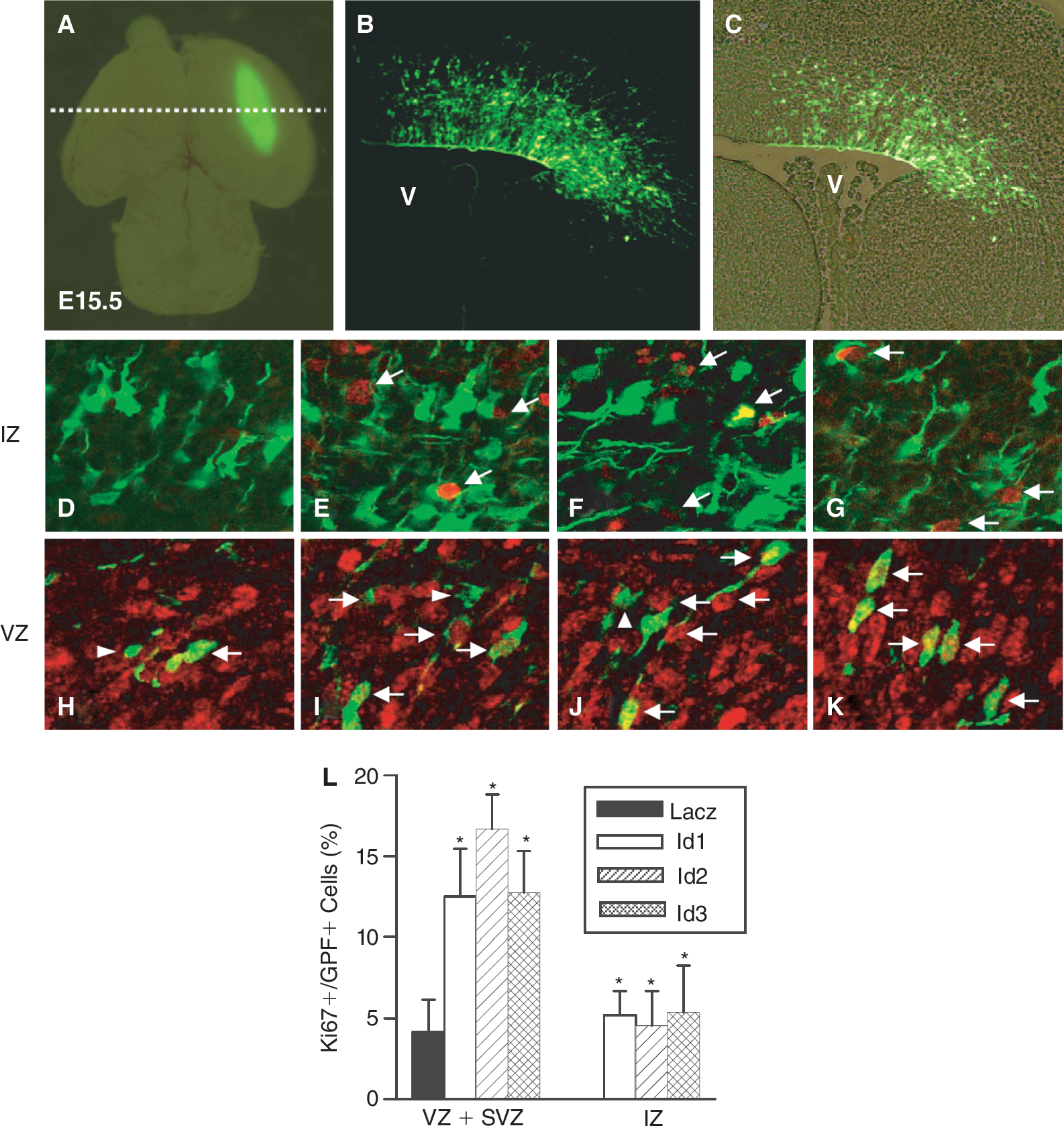
Electroporation of inhibitor of differentiation (Id) increases proliferation of neural stem cells (NSCs) in the embryonic brain. Mouse telencephalic vesicles (E13.5) were electroporated with LacZ-epidermal growth factor (EGF)P-CLBC3 (
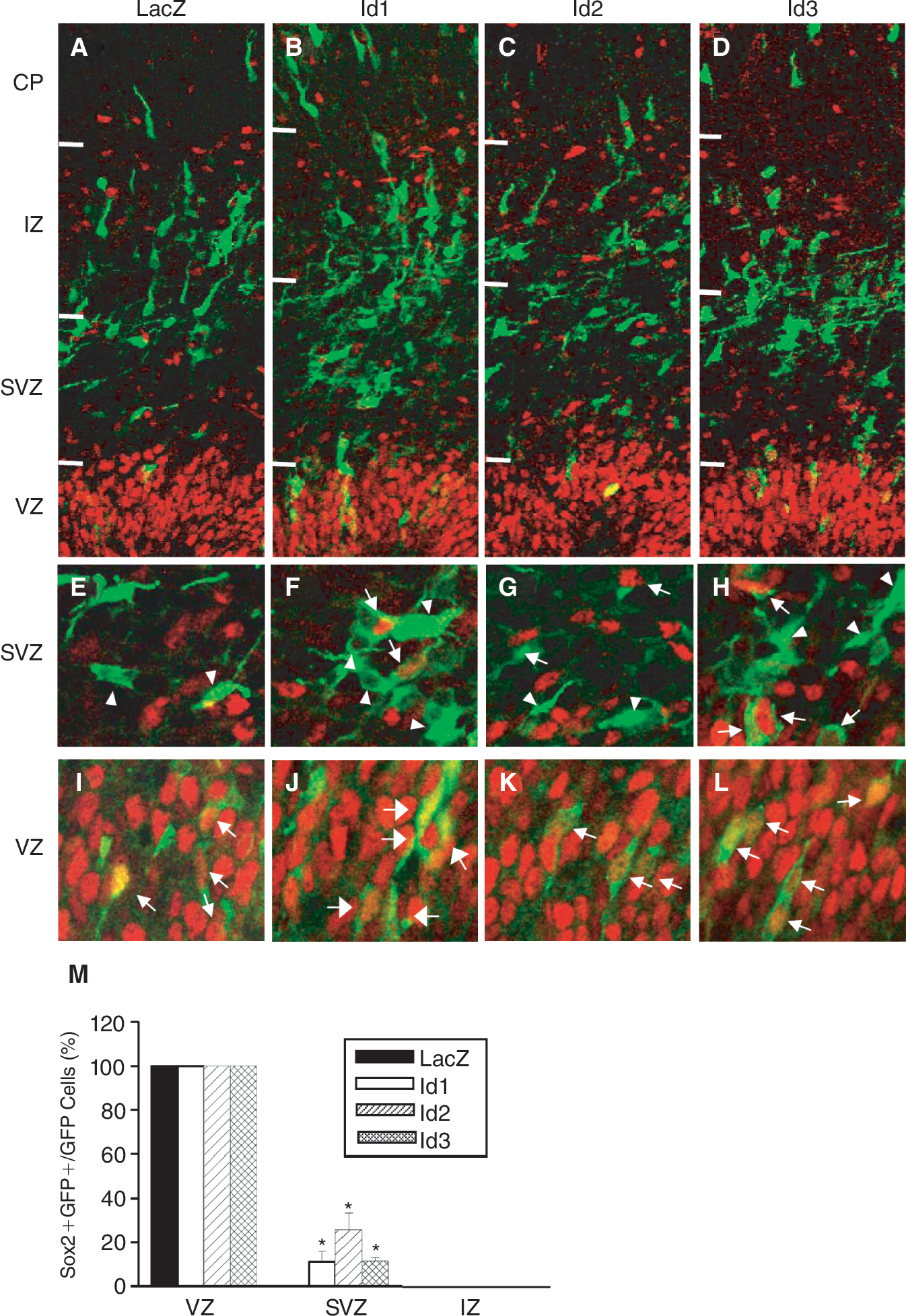
Inhibitor of differentiation (Id) genes can increase Sox2-positive cells in the developing embryonic brain. Two days later, electroporation embryos were stained for the presence of GFP (green) and Sox2 (red). In SVZ, among Id-expressing GFP-positive cells of SVZ, Sox2-positive cells were found (arrows in
Discussion
In this study, we show that Id proteins promote proliferation and self-renewal of NSCs. The stimulating functions of Id proteins in NSCs are derived from their ability to interfere with functional complex formation between proneural bHLH transcription factors and E-proteins.
During development of the nervous system, NSCs proliferate in response to trophic stimuli including fibroblast growth factor (FGF), epidermal growth factor (EGF), and Delta-Notch [32,33]. Proliferation of NSCs is important for expansion of the neural tissue to the appropriate size and is finely tuned by positive and negative regulators of differentiation as mentioned earlier. In the absence of trophic signals, NSCs gradually differentiate into lineage-restricted cells while migrating to the ultimate position [34]. The lineage-restricted neuronal differentiation as well as cell-type specification is regulated by transcriptional cascades of proneural bHLH factors [35]. In this study, we showed that Id proteins can antagonize the functions of proneural bHLH factors. HIT cells endogenously express NeuroD, a proneural bHLH transcription factor that is required for development of hippocampal [36] and cerebellar granule cells [37] in addition to pancreatic β-cell functions [38]. The results with HIT cells indicate that Id proteins can interfere with the functional complex formation of NeuroD::E47 (Fig. 3B) and their subsequent binding to the E-box sequence (Fig. 3C). This artificial in vitro assay does not necessarily mean that NeuroD is the target that Id proteins counteract in vivo. The cells in neurospheres express other proneural bHLH transcription factors such as Mash1, Ngn1, and Ngn2 in addition to Id proteins (data not shown). In situ hybridization assays locate the expression of Id1 and Id3 in the ventricular zone of the developing telencephalic vesicles where the progenitor cells actively proliferate and take up BrdU (Supplementary Fig. 1; Supplementary materials are available online at
The Id's counteracting functions against proneural bHLH factors are important for maintenance of proliferative stages of NSCs. Id proteins increased proliferation of NSCs in neurosphere-forming cultures (Fig. 2) as well as in monolayer cultures (Supplementary Fig. 2). In monolayer culture, however, the effect of Id2 was not profound compared to its effects shown in Figures 2 and 4(Supplementary Fig. 2). Further study may unravel the molecular basis of Id2's diverse functions in neurosphere cultures (Fig. 2), monolayer cultures in the presence of fibronectin (Supplementary Fig. 2), or developing telencephalon (Fig. 4). Removing trophic signals reduces the Id expression and concomitant differentiation into either neuronal or glial cells (Fig. 1). Interestingly, persistent expression of Id proteins forces the cells to differentiate into astrocytes while inhibiting neuronal differentiation (Figs. 2B and 3D). The multiple functions of Id proteins during differentiation are similar to those of Hes proteins.
Specifically, Hes proteins inhibit the proneural bHLH factors in dual modes. Homo- and heterodimers of Hes proteins repress the expression of proneural bHLH transcription factors through binding to N-boxes and interfere with the protein functions of proneural bHLH transcription factors. Expression of Hes1 oscillates because the Hes1 promoter contains 3 N-boxes that can be repressed by its own gene product. During early chick brain development, Id can interact with Hes1 and release the autoregulatory negative feedback. Therefore, Hes1 expression is sustained in NSCs [42]. In our study, however, expression levels of Hes1 and Hes5 in Id-overexpressing cells were shown to be similar to that in LacZ-expressing NSCs. In contrast, under the same condition, the nestin expression was increased in Id-overexpressing cells (Fig. 2C), suggesting that the enhanced stem cell property following Id overexpression is not due to elevated expression of Hes1 and Hes5. The discrepancy may be due to the various sources of neural precursor cells. Hes1 is known to be expressed in 2 different modes in a region-specific manner. In the non-boundary regions, the Hes1 expression oscillates due to the negative feedback loop [43]. In the boundary regions where the cell proliferation is limited, Hes1 expression is persistent probably due to the presence of Id proteins [42,44]. The NSCs used in the present study were originally isolated from the non-boundary telencephalic vesicles. Overexpression of Id proteins in the similar region by in utero electroporation yields higher numbers of Sox2- and Ki67-positive cells (Figs. 4 and 5). Taken together, Id proteins may regulate proliferation rates of neural precursors in Hes-dependent and -independent manners depending on the locations in the developing brain regions.
Although Id and Hes share multiple strategies in mediating negative effects on neuronal differentiation, Id proteins may also have unique function of regulating cell cycle progression. Id2 and Id4 stimulate the cell cycle progression by inhibiting the interaction of pRb and E2F [45]. Id1 and Id3 can inhibit cyclin-dependent kinase inhibitors such as p2iWAF1/Cip1 [46] and p16INK4a [47]; Id2 can suppress p57Kip2 [48]. The cell cycle progression by Id proteins is more prominent in cancer cells. Id1 expression is elevated in human breast cancer [49] and seminoma [50], while Id2 is expressed at a high level in pancreatic cancer [51]. In addition, Id genes can immortalize fibroblast cells when co-transfected with Bcl-2 gene [52] or human keratinocytes [53]. Thus, proliferative effects of Id proteins in NSCs may be partly ascribed to the Id's unique functions of promoting cell cycle progression.
Id genes block precocious differentiation of cortical progenitors into neurons and oligodendrocytes, but not astrocytes [54,55]. It has been shown that oligodendrocyte differentiation requires bHLH transcription factors such as Olig1 and Olig2 [56] and Id proteins can block oligodendrocyte differentiation by sequestering Olig1/2 [57]. In contrast, astrocyte differentiation is facilitated by Id proteins (Fig. 3), suggesting that astrogliogenesis may not require any bHLH transcription factors. Astrocyte differentiation is known to be regulated by intrinsic and extrinsic factors. JAK-STAT3 pathway is a key component of the signaling machinery promoting astrocyte differentiation [58,59]. However, NSCs from the E12.5 embryo are insensitive to the gliogenic activity of cytokines [60,61] since STAT3-binding sites of the GFAP and S100β promoters are methylated and are unable to be activated by STAT3 [62,63]. Hes proteins associate with JAK2 and STAT3, and facilitate phosphorylation and activation of STAT3 [64]. Proneural bHLH transcription factors such as Ngn1 sequester co-activators such as p300/CREB-binding protein (CBP) away from STAT3 and block astrocyte differentiation [65]. Therefore, it is conceivable that persistent expression of Id proteins promote astrocyte differentiation by sequestering proneural bHLH factors away from the neuronal differentiation genes and releasing more p300/CBP that would be subsequently available for interaction with STAT3.
In conclusion, Id proteins play important roles in expansion of undifferentiated neural precursor cells by increasing the self-renewal and proliferation potentials of neural precursor cells. The stimulatory effects of Id proteins are derived from sequestration of proneural bHLH transcription factors and possibly from Id's unique function as cell cycle regulators as Id proteins do not increase the Hes expression. Further studies will be necessary to examine whether Id proteins sequester cell cycle inhibitors and directly regulate the cell cycle progression in NSCs.
Footnotes
Acknowledgments
This work was partly supported by Korean Research Foundation Grant (KRF-2002-042-C00081) to YDL and HSK; and Brain Research Center of the 21st Century Frontier Research Program (M103KV010008-07K2201-00810) and the Korea Health 21R&D Project (0405-DB01-0104-0006), Ministry of Health & Welfare, and the Priority Center Program (2009-0093826) to HSK.
Author Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.